
A Rare Case of TP63-Associated Lymphopenia Revealed by Newborn Screening Using TREC

 , ,
, ,  , , , , , , , ,
, , , , , , , ,
Abstract
1. Introduction
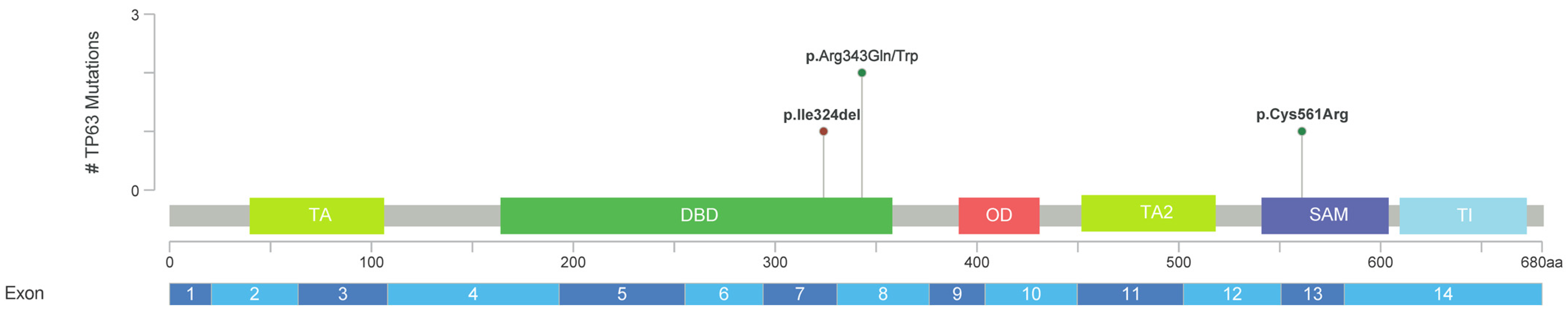
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tangye, S.G.; Al-Herz, W.; Bousfiha, A.; Cunningham-Rundles, C.; Franco, J.L.; Holland, S.M.; Klein, C.; Morio, T.; Oksenhendler, E.; Picard, C.; et al. Human Inborn Errors of Immunity: 2022 Update on the Classification from the International Union of Immunological Societies Expert Committee. J. Clin. Immunol. 2022, 42, 1473–1507. [Google Scholar] [CrossRef] [PubMed]
- Lev, A.; Somech, R.; Somekh, I. Newborn screening for severe combined immunodeficiency and inborn errors of immunity. Curr. Opin. Pediatr. 2023, 35, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Amatuni, G.S.; Currier, R.J.; Church, J.A.; Bishop, T.; Grimbacher, E.; Nguyen, A.A.; Agarwal-Hashmi, R.; Aznar, C.P.; Butte, M.J.; Cowan, M.J.; et al. Newborn Screening for Severe Combined Immunodeficiency and T-cell Lymphopenia in California, 2010-2017. Pediatrics 2019, 143, e20182300. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.Y.; Mukhina, A.A.; Balinova, N.V.; Matulevich, S.A.; Pershin, D.E.; Khoreva, A.L.; Marakhonov, A.V.; Voronin, S.V.; Zinchenko, R.A.; Shcherbina, A.Y.; et al. Newborn screening for primary immunodeficiencies as a way to detect syndromal disorders in neonates: A clinical case of 22q11.2DS syndrome. Pediatr. Hematol./Oncol. Immunopathol. 2022, 21, 158–162. (In Russian) [Google Scholar] [CrossRef]
- Li, Y.; Giovannini, S.; Wang, T.; Fang, J.; Li, P.; Shao, C.; Wang, Y.; TOR centre; Shi, Y.; Candi, E.; et al. p63: A crucial player in epithelial stemness regulation. Oncogene 2023, 42, 3371–3384. [Google Scholar] [CrossRef]
- van Bokhoven, H.; Brunner, H.G. Splitting p63. Am. J. Hum. Genet. 2002, 71, 1–13. [Google Scholar] [CrossRef]
- Sutton, V.R.; van Bokhoven, H. TP63-Related Disorders. In GeneReviews((R)); Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Giampietro, P.F.; Baker, M.W.; Basehore, M.J.; Jones, J.R.; Seroogy, C.M. Novel mutation in TP63 associated with ectrodactyly ectodermal dysplasia and clefting syndrome and T cell lymphopenia. Am. J. Med. Genetics. Part. A 2013, 161A, 1432–1435. [Google Scholar] [CrossRef]
- Helenius, K.; Ojala, L.; Kainulainen, L.; Peltonen, S.; Hietala, M.; Pohjola, P.; Parikka, V. Overlap between EEC and AEC syndrome and immunodeficiency in a preterm infant with a TP63 variant. Eur. J. Med. Genet. 2023, 66, 104735. [Google Scholar] [CrossRef]
- Voronin, S.V.; Zakharova, E.Y.; Baydakova, G.V.; Marakhonov, A.V.; Shchagina, O.A.; Ryzhkova, O.P.; Shilova, N.V.; Rumyantsev, A.G.; Shcherbina, A.Y.; Mukhina, A.A.; et al. Advanced neonatal screening for hereditary diseases in Russia: First results and future prospects. Pediatr. n.a. G.N. Speransky 2024, 103, 16–29. (In Russian) [Google Scholar] [CrossRef]
- Celli, J.; Duijf, P.; Hamel, B.C.; Bamshad, M.; Kramer, B.; Smits, A.P.; Newbury-Ecob, R.; Hennekam, R.C.; Van Buggenhout, G.; van Haeringen, A.; et al. Heterozygous germline mutations in the p53 homolog p63 are the cause of EEC syndrome. Cell 1999, 99, 143–153. [Google Scholar] [CrossRef]
- Wenger, T.; Li, D.; Harr, M.H.; Tan, W.H.; Pellegrino, R.; Stark, Z.; Hakonarson, H.; Bhoj, E.J. Expanding the phenotypic spectrum of TP63-related disorders including the first set of monozygotic twins. Am. J. Med. Genetics. Part. A 2018, 176, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Francioli, L.C.; Goodrich, J.K.; Collins, R.L.; Kanai, M.; Wang, Q.; Alfoldi, J.; Watts, N.A.; Vittal, C.; Gauthier, L.D.; et al. A genomic mutational constraint map using variation in 76,156 human genomes. Nature 2023, 625, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.H.M.; Pires, D.E.V.; Ascher, D.B. DynaMut2: Assessing changes in stability and flexibility upon single and multiple point missense mutations. Protein Sci. 2021, 30, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Kaghad, M.; Wang, Y.; Gillett, E.; Fleming, M.D.; Dotsch, V.; Andrews, N.C.; Caput, D.; McKeon, F. p63, a p53 homolog at 3q27-29, encodes multiple products with transactivating, death-inducing, and dominant-negative activities. Mol. Cell 1998, 2, 305–316. [Google Scholar] [CrossRef]
- Flores, E.R.; Tsai, K.Y.; Crowley, D.; Sengupta, S.; Yang, A.; McKeon, F.; Jacks, T. p63 and p73 are required for p53-dependent apoptosis in response to DNA damage. Nature 2002, 416, 560–564. [Google Scholar] [CrossRef]
- Finlan, L.E.; Hupp, T.R. p63: The phantom of the tumor suppressor. Cell Cycle 2007, 6, 1062–1071. [Google Scholar] [CrossRef]
- Wu, G.; Nomoto, S.; Hoque, M.O.; Dracheva, T.; Osada, M.; Lee, C.C.; Dong, S.M.; Guo, Z.; Benoit, N.; Cohen, Y.; et al. DeltaNp63alpha and TAp63alpha regulate transcription of genes with distinct biological functions in cancer and development. Cancer Res. 2003, 63, 2351–2357. [Google Scholar]
- Melino, G.; Memmi, E.M.; Pelicci, P.G.; Bernassola, F. Maintaining epithelial stemness with p63. Sci. Signal 2015, 8, re9. [Google Scholar] [CrossRef]
- Kawata, M.; Taniguchi, Y.; Mori, D.; Yano, F.; Ohba, S.; Chung, U.I.; Shimogori, T.; Mills, A.A.; Tanaka, S.; Saito, T. Different regulation of limb development by p63 transcript variants. PLoS ONE 2017, 12, e0174122. [Google Scholar] [CrossRef] [PubMed]
- Gawrych, E.; Binczak-Kuleta, A.; Janiszewska-Olszowska, J.; Ciechanowicz, A. Ectrodactyly-ectodermal dysplasia-cleft syndrome (EEC syndrome) with a developmental delay caused by R304W mutation in the tp63 gene. Ann. Acad. Med. Stetin. 2013, 59, 11–14. [Google Scholar] [PubMed]
- Beaudry, V.G.; Pathak, N.; Koster, M.I.; Attardi, L.D. Differential PERP regulation by TP63 mutants provides insight into AEC pathogenesis. Am. J. Med. Genetics. Part. A 2009, 149A, 1952–1957. [Google Scholar] [CrossRef] [PubMed]
- Dotto, J.; Pelosi, G.; Rosai, J. Expression of p63 in thymomas and normal thymus. Am. J. Clin. Pathol. 2007, 127, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Pan, W.; Zheng, L.; Zhong, X.P.; Tan, L.; Liang, Z.; He, J.; Feng, P.; Zhao, Y.; Qiu, Y.R. Thymic Epithelial Cells Contribute to Thymopoiesis and T Cell Development. Front. Immunol. 2019, 10, 3099. [Google Scholar] [CrossRef]
- Martinez-Ruiz, G.U.; Morales-Sanchez, A.; Bhandoola, A. Transcriptional and epigenetic regulation in thymic epithelial cells. Immunol. Rev. 2022, 305, 43–58. [Google Scholar] [CrossRef]
- Collins, C.; Sharpe, E.; Silber, A.; Kulke, S.; Hsieh, E.W.Y. Congenital Athymia: Genetic Etiologies, Clinical Manifestations, Diagnosis, and Treatment. J. Clin. Immunol. 2021, 41, 881–895. [Google Scholar] [CrossRef]
- Di Como, C.J.; Urist, M.J.; Babayan, I.; Drobnjak, M.; Hedvat, C.V.; Teruya-Feldstein, J.; Pohar, K.; Hoos, A.; Cordon-Cardo, C. p63 expression profiles in human normal and tumor tissues. Clin. Cancer Res. 2002, 8, 494–501. [Google Scholar]
- Candi, E.; Rufini, A.; Terrinoni, A.; Giamboi-Miraglia, A.; Lena, A.M.; Mantovani, R.; Knight, R.; Melino, G. DeltaNp63 regulates thymic development through enhanced expression of FgfR2 and Jag2. Proc. Natl. Acad. Sci. USA 2007, 104, 11999–12004. [Google Scholar] [CrossRef]
- Senoo, M.; Pinto, F.; Crum, C.P.; McKeon, F. p63 Is essential for the proliferative potential of stem cells in stratified epithelia. Cell 2007, 129, 523–536. [Google Scholar] [CrossRef]
- Russo, C.; Osterburg, C.; Sirico, A.; Antonini, D.; Ambrosio, R.; Wurz, J.M.; Rinnenthal, J.; Ferniani, M.; Kehrloesser, S.; Schafer, B.; et al. Protein aggregation of the p63 transcription factor underlies severe skin fragility in AEC syndrome. Proc. Natl. Acad. Sci. USA 2018, 115, E906–E915. [Google Scholar] [CrossRef] [PubMed]
- Akahoshi, K.; Sakazume, S.; Kosaki, K.; Ohashi, H.; Fukushima, Y. EEC syndrome type 3 with a heterozygous germline mutation in the P63 gene and B cell lymphoma. Am. J. Med. Genet. Part A 2003, 120A, 370–373. [Google Scholar] [CrossRef]
- Osterburg, C.; Dotsch, V. Structural diversity of p63 and p73 isoforms. Cell Death Differ. 2022, 29, 921–937. [Google Scholar] [CrossRef] [PubMed]
- de Bruijn, I.; Kundra, R.; Mastrogiacomo, B.; Tran, T.N.; Sikina, L.; Mazor, T.; Li, X.; Ochoa, A.; Zhao, G.; Lai, B.; et al. Analysis and Visualization of Longitudinal Genomic and Clinical Data from the AACR Project GENIE Biopharma Collaborative in cBioPortal. Cancer Res. 2023, 83, 3861–3867. [Google Scholar] [CrossRef] [PubMed]
- Marakhonov, A.V.; Efimova, I.Y.; Mukhina, A.A.; Zinchenko, R.A.; Balinova, N.V.; Rodina, Y.; Pershin, D.; Ryzhkova, O.P.; Orlova, A.A.; Zabnenkova, V.V.; et al. Newborn Screening for Severe T and B Cell Lymphopenia Using TREC/KREC Detection: A Large-Scale Pilot Study of 202,908 Newborns. J. Clin. Immunol. 2024, 44, 93. [Google Scholar] [CrossRef]
- Voronin, S.V.; Zinchenko, R.A.; Efimova, I.Y.; Kutsev, S.I.; Marakhonov, A.V.; Mukhina, A.A.; Novichkova, G.A.; Pershin, D.E.; Rodina, Y.A.; Rumyantsev, A.G.; et al. Neonatal screening, postnatal diagnosis and tactics of preclinical treatment and prevention of primary immunodeficiencies in children. Guidelines by the experts from the National Association of Experts in Primary Immunodeficiencies (NAEPID) and the Association of Medical Genetics (AMG) of Russia. Pediatr. n.a. G.N. Speransky 2023, 102, 11–33. (In Russian) [Google Scholar] [CrossRef]
- Rudenskaya, G.E.; Marakhonov, A.V.; Shchagina, O.A.; Lozier, E.R.; Dadali, E.L.; Akimova, I.A.; Petrova, N.V.; Konovalov, F.A. Ataxia with Oculomotor Apraxia Type 4 with PNKP Common “Portuguese” and Novel Mutations in Two Belarusian Families. J. Pediatr. Genet. 2019, 8, 58–62. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Parameter | 19 Days of Life * | 12 Months of Life | Normal Range |
|---|---|---|---|
| T cells (CD3+), 109/L | 1.92 | 2.01 | 3.3–6.5 (58–71%) |
| Th (CD4+), 109/L (% of T cells) | 1.22 (63.4%) | 1.15 (57.2%) | 1.9–4.4 (56–79%) |
| Th naïve (CD45RA+CD197+), cells/µL (% of CD4+) | 809 (66.2%) | 773.52 (67.2%) | 1442–3731 (72–86%) |
| T-central memory, cells/µL (% of CD4+) | 284 (23.3%) | 115(10%) | 151–448 (4.5–15.1%) |
| Effector memory, cells/µL (% of CD4+) | 105 (8.6%) | 199 (17.3%) | 126–399 (4.2–13.9) |
| TEMRA, cells/µL (% of CD4+) | 24 (2%) | 64 (5.6%) | 36–225 (1.3–6.9%) |
| Tc (CD8+), 109/L (% of T cells) | 0.66 (34%) | 0.73 (36.1%) | 0.7–1.9 (18–39%) |
| Tc naïve (CD45RA+CD197+), cells/µL (% of CD8+ cells) | 600 (91.5%) | 352 (48.5%) | 455–1515 (34–88%) |
| T-central memory, cells/µL (% of CD8+) | 37 (5.6%) | 18 (2.6%) | 6–85 (0.4–8.0%) |
| Effector memory, cells/µL (% of CD8+) | 11 (1.7%) | 245 (33%) | 50–395 (3.4–24.4%) |
| TEMRA, cells/µL (% of CD8+) | 9 (1.3%) | 109 (15.1%) | 69–736 (6.1–39.0%) |
| B cells (CD19+), 109/L (% of Lymphocytes) | 1.78 (40%) | 2.57 (45%) | 1–3 (17–36%) |
| B naïve (CD19+IgD+IgM+CD27−), cells/µL (% of B cells) | n/a | 2360 (91.9%) | 290–1637 (62–83%) |
| Transitional B cells (CD19+IgM+CD38+), cells/µL (% of B cells) | n/a | 41 (1.6%) | 32–235 (3.4–11.9%) |
| Natural effector B cells (IgD+IgM+CD27+), cells/µL (% of B cells) | n/a | 67 (2.6%) | 15–242 (2.7–14.6%) |
| IgM-only B cells (CD27+IgM+IgD−), cells/µL (% of B cells) | n/a | 5 (0.2%) | 2–28 (0.2–2.0%) |
| Switched B memory cells (IgD−IgM−CD27+), cells/µL (% of B cells) | n/a | 36 (1.4%) | 5–83 (0.5–5.1%) |
| Plasmablasts (CD19+IgD−IgM−CD27+CD38high), cells/µL (% of B cells) | n/a | 2.6 (0.1%) | 1–28.5 (0.07–3.17%) |
| Plasma B cells (CD19+IgD−IgM−CD27+CD38highCD138+), cells/µL (% of B cells) | n/a | 0 (0%) | – |
| CD21lowCD38low B cells, cells/µL (% of B cells) | n/a | 72 (2.8%) | 7–75 (1.1–5.0%) |
| NK cells (CD3−CD16+CD56+), 109/L (% of Lymphocytes) | 0.7 (15.9%) | 0.9 (16.1%) | 0.2–1.3 (3–16%) |
| CD56+high NK, cell/µL (% of NK-cells) | 100 (15.6%) | 70 (14%) | 10–160 (2.6–15.3%) |
| T-NK-cells CD3+CD56+, cell/µL (% of Lymphocytes) | 0 | 6 (0.1%) | 5–30 (0.05–0.39%) |
| IgG, g/L | n/a | 7.02 | 4.75–12.10 |
| IgM, g/L | n/a | 0.87 | 0.41–1.83 |
| IgA, g/L | n/a | 0.55 | 0.21–2.91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marakhonov, A.; Serebryakova, E.; Mukhina, A.; Vechkasova, A.; Prokhorov, N.; Efimova, I.; Balinova, N.; Lobenskaya, A.; Vasilyeva, T.; Zabnenkova, V.; et al. A Rare Case of TP63-Associated Lymphopenia Revealed by Newborn Screening Using TREC. Int. J. Mol. Sci. 2024, 25, 10844. https://doi.org/10.3390/ijms251910844
Marakhonov A, Serebryakova E, Mukhina A, Vechkasova A, Prokhorov N, Efimova I, Balinova N, Lobenskaya A, Vasilyeva T, Zabnenkova V, et al. A Rare Case of TP63-Associated Lymphopenia Revealed by Newborn Screening Using TREC. International Journal of Molecular Sciences. 2024; 25(19):10844. https://doi.org/10.3390/ijms251910844
Chicago/Turabian StyleMarakhonov, Andrey, Elena Serebryakova, Anna Mukhina, Anastasia Vechkasova, Nikolai Prokhorov, Irina Efimova, Natalia Balinova, Anastasia Lobenskaya, Tatyana Vasilyeva, Victoria Zabnenkova, and et al. 2024. "A Rare Case of TP63-Associated Lymphopenia Revealed by Newborn Screening Using TREC" International Journal of Molecular Sciences 25, no. 19: 10844. https://doi.org/10.3390/ijms251910844
APA StyleMarakhonov, A., Serebryakova, E., Mukhina, A., Vechkasova, A., Prokhorov, N., Efimova, I., Balinova, N., Lobenskaya, A., Vasilyeva, T., Zabnenkova, V., Ryzhkova, O., Rodina, Y., Pershin, D., Soloveva, N., Fomenko, A., Saydaeva, D., Ibisheva, A., Irbaieva, T., Koroteev, A., ... Kutsev, S. (2024). A Rare Case of TP63-Associated Lymphopenia Revealed by Newborn Screening Using TREC. International Journal of Molecular Sciences, 25(19), 10844. https://doi.org/10.3390/ijms251910844