
Y-Box Binding Protein 1: Unraveling the Multifaceted Role in Cancer Development and Therapeutic Potential



Abstract
:1. Introduction
2. YBX1: Multifaceted Roles in Biology
3. Effects of YBX1 on Cancer Hallmarks Originated from Cancer Itself
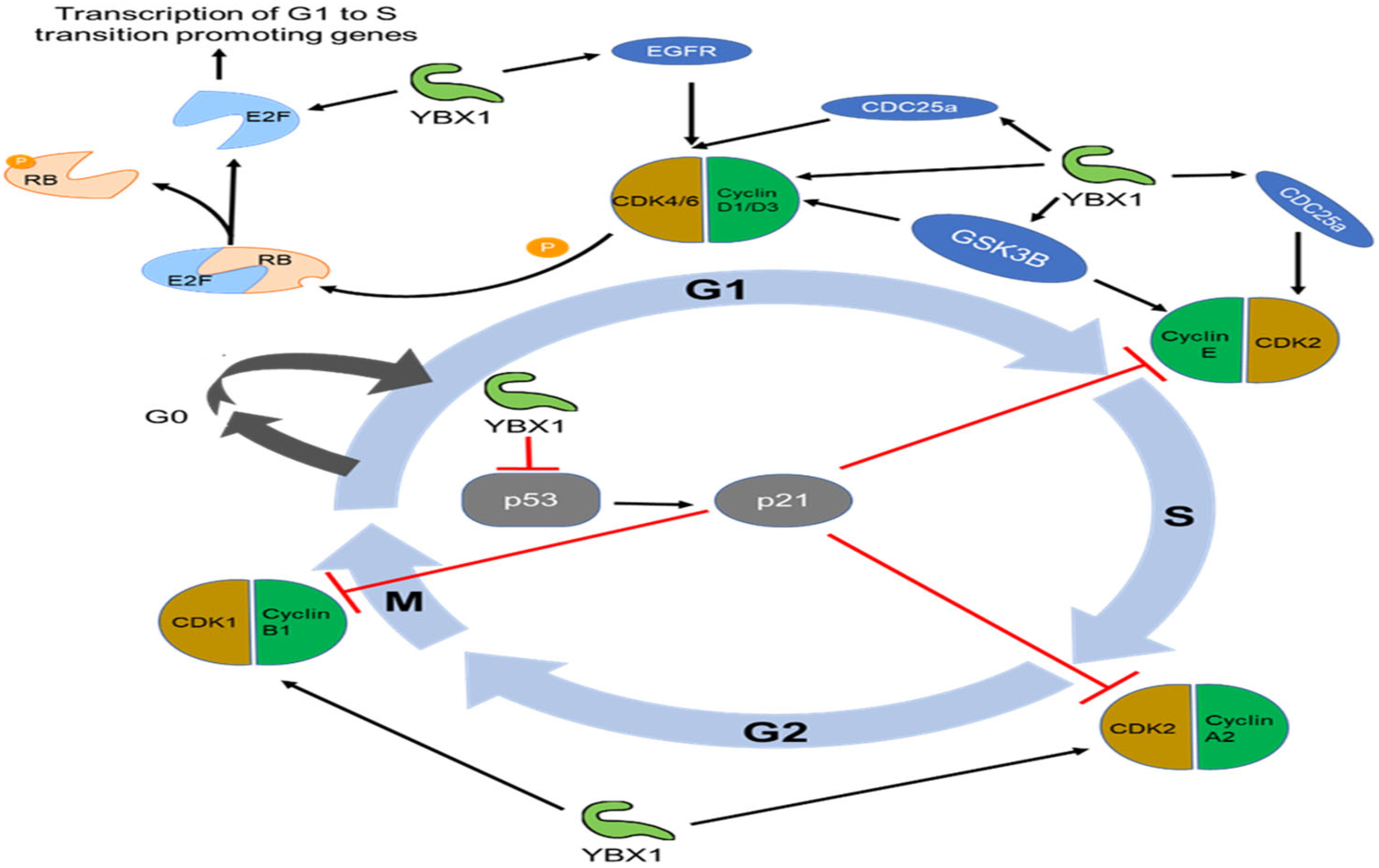
3.1. Role of YBX1 in Sustaining Proliferative Signaling and Evading Growth Suppressors
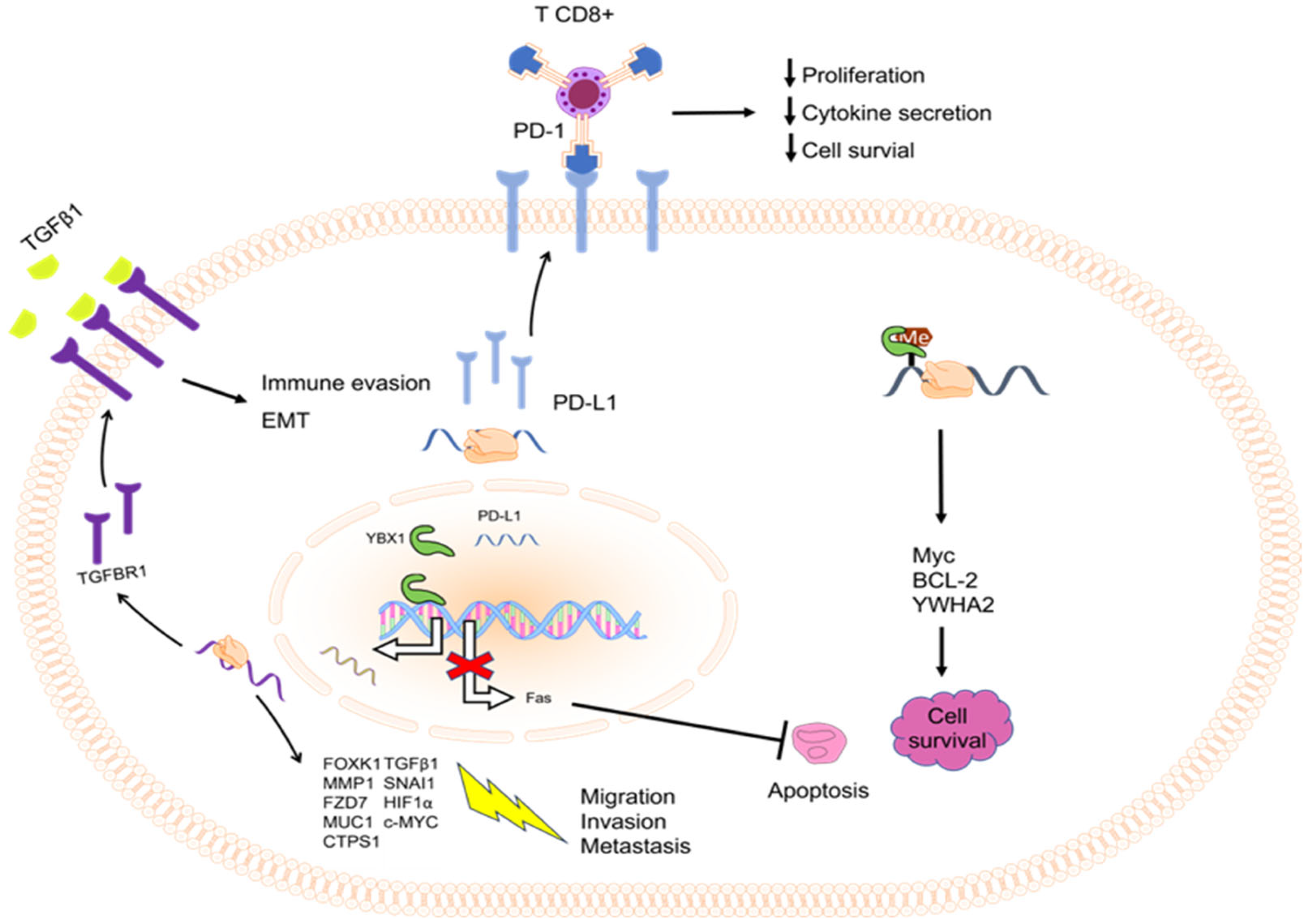
3.2. Role of YBX1 in Cell Death Resistance
3.3. Role of YBX1 in Enabling Replicative Immortality and Senescence
3.4. Role of YBX1 in Activating Invasion and Metastasis

3.5. Role of YBX1 in Deregulating Cellular Energetics
3.6. Role of YBX1 in Genome Instability and Mutation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Partner | Name | Reference | Cancer Type |
|---|---|---|---|---|
| Bind to and stabilize YBX1 mRNA or enhance S102 phosphorylation leading to facilitating translation or transcription of target genes | Long noncoding RNA (lncRNA) | HOXC-AS3 | [174,175] | Lung cancer, pancreatic cancer, esophageal squamous cell carcinoma, breast cancer, acute myeloid leukemia, clear cell renal cell carcinoma, hepatocellular carcinoma (HCC), skin cancer |
| HIF1A-AS1 | [129] | |||
| MIR31HG | [94] | |||
| LINC00857 | [176] | |||
| linc02042 | [177] | |||
| AC073352.1 | [137] | |||
| GAS6-AS1 | [178] | |||
| MILIP | [124] | |||
| USP2-AS1 | [179] | |||
| HUMT | [122] | |||
| BASP1-AS1 | [180] | |||
| DSCAM-AS1 | [181] | |||
| LINC00941 | [182] | |||
| HOTAIR | [183] | |||
| GAS5 | [184] | |||
| SNHG6 | [185] | |||
| linc00665 | [186] | |||
| PIK3CD-AS2 | [187] | |||
| Circular RNA (circRNA) | circACTN4 | [120] | Intrahepatic cholangiocarcinoma, nasopharyngeal carcinoma, HCC | |
| circIPO7 | [106] | |||
| hsa_circ_0062682 | [188] | |||
| Bind to and inhibit YBX1 or nuclear translocation | lncRNA | Linc01612 | [189] | HCC, lung adenocarcinoma, colorectal cancer |
| LINC00472 | [190] | |||
| TP53TG1 | [191] | |||
| HITT | [192] | |||
| circRNA | circNEIL3 | [119] | Colorectal carcinoma, gastric cancer | |
| circFAT1(e2) | [193] | |||
| Micro RNA (miRNA) | miR-6509-5p | [194] | Gastric cancer, diffuse large B-cell lymphoma, malignant pleural mesothelioma, cervical carcinoma, breast cancer, renal carcinoma | |
| MiR-216a | [195] | |||
| miR-137 | [196,197] | |||
| miR-375 | [198,199,200] |
4. Role of YBX1 in Tumor Microenvironment
4.1. YBX1 in Inducing Angiogenesis
4.2. YBX1 in Avoiding Immune Destruction
4.3. YBX1 in Tumor-Promoting Inflammation
4.4. Role of YBX1 in Neural Input
4.5. Effects of YBX1 on Extracellular Matrix
5. Targeting YBX1 in Cancer Therapy Research and Development
6. Perspective
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, K.; Nakano, K.; Cui, L.; Mizuuchi, Y.; Onishi, H.; Oda, Y.; Obika, S.; Tanaka, M.; Katano, M. Nuclear expression of Y-box binding protein-1 is associated with poor prognosis in patients with pancreatic cancer and its knockdown inhibits tumor growth and metastasis in mice tumor models. Int. J. Cancer 2016, 139, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Kawahara, A.; Basaki, Y.; Hattori, S.; Nakashima, K.; Nakano, K.; Shirouzu, K.; Kohno, K.; Yanagawa, T.; Yamana, H.; et al. Expression of HER2 and estrogen receptor alpha depends upon nuclear localization of Y-box binding protein-1 in human breast cancers. Cancer Res. 2008, 68, 1504–1512. [Google Scholar] [CrossRef]
- Dahl, E.; En-Nia, A.; Wiesmann, F.; Krings, R.; Djudjaj, S.; Breuer, E.; Fuchs, T.; Wild, P.J.; Hartmann, A.; Dunn, S.E.; et al. Nuclear detection of Y-box protein-1 (YB-1) closely associates with progesterone receptor negativity and is a strong adverse survival factor in human breast cancer. BMC Cancer 2009, 9, 410. [Google Scholar] [CrossRef]
- Gessner, C.; Woischwill, C.; Schumacher, A.; Liebers, U.; Kuhn, H.; Stiehl, P.; Jürchott, K.; Royer, H.D.; Witt, C.; Wolff, G. Nuclear YB-1 expression as a negative prognostic marker in nonsmall cell lung cancer. Eur. Respir. J. 2004, 23, 14–19. [Google Scholar] [CrossRef]
- Shibahara, K.; Sugio, K.; Osaki, T.; Uchiumi, T.; Maehara, Y.; Kohno, K.; Yasumoto, K.; Sugimachi, K.; Kuwano, M. Nuclear expression of the Y-box binding protein, YB-1, as a novel marker of disease progression in non-small cell lung cancer. Clin. Cancer Res. 2001, 7, 3151–3155. [Google Scholar]
- Chatterjee, M.; Rancso, C.; Stühmer, T.; Eckstein, N.; Andrulis, M.; Gerecke, C.; Lorentz, H.; Royer, H.D.; Bargou, R.C. The Y-box binding protein YB-1 is associated with progressive disease and mediates survival and drug resistance in multiple myeloma. Blood 2008, 111, 3714–3722. [Google Scholar] [CrossRef]
- Oda, Y.; Sakamoto, A.; Shinohara, N.; Ohga, T.; Uchiumi, T.; Kohno, K.; Tsuneyoshi, M.; Kuwano, M.; Iwamoto, Y. Nuclear expression of YB-1 protein correlates with P-glycoprotein expression in human osteosarcoma. Clin. Cancer Res. 1998, 4, 2273–2277. [Google Scholar] [PubMed]
- Oda, Y.; Ohishi, Y.; Saito, T.; Hinoshita, E.; Uchiumi, T.; Kinukawa, N.; Iwamoto, Y.; Kohno, K.; Kuwano, M.; Tsuneyoshi, M. Nuclear expression of Y-box-binding protein-1 correlates with P-glycoprotein and topoisomerase II alpha expression, and with poor prognosis in synovial sarcoma. J. Pathol. 2003, 199, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Bonafé, P.; Fedoruk, M.N.; Whitmore, T.G.; Akbari, M.; Ralph, J.L.; Ettinger, S.; Gleave, M.E.; Nelson, C.C. YB-1 is upregulated during prostate cancer tumor progression and increases P-glycoprotein activity. Prostate 2004, 59, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Basaki, Y.; Hosoi, F.; Oda, Y.; Fotovati, A.; Maruyama, Y.; Oie, S.; Ono, M.; Izumi, H.; Kohno, K.; Sakai, K.; et al. Akt-dependent nuclear localization of Y-box-binding protein 1 in acquisition of malignant characteristics by human ovarian cancer cells. Oncogene 2007, 26, 2736–2746. [Google Scholar] [CrossRef] [PubMed]
- Kloks, C.P.; Spronk, C.A.; Lasonder, E.; Hoffmann, A.; Vuister, G.W.; Grzesiek, S.; Hilbers, C.W. The solution structure and DNA-binding properties of the cold-shock domain of the human Y-box protein YB-1. J. Mol. Biol. 2002, 316, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Wolffe, A.P. Structural and functional properties of the evolutionarily ancient Y-box family of nucleic acid binding proteins. Bioessays 1994, 16, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Lyabin, D.N.; Eliseeva, I.A.; Ovchinnikov, L.P. YB-1 protein: Functions and regulation. Wiley Interdiscip. Rev. RNA 2014, 5, 95–110. [Google Scholar] [CrossRef]
- Ito, K.; Tsutsumi, K.; Kuzumaki, T.; Gomez, P.F.; Otsu, K.; Ishikawa, K. A novel growth-inducible gene that encodes a protein with a conserved cold-shock domain. Nucleic Acids Res. 1994, 22, 2036–2041. [Google Scholar] [CrossRef]
- Kashanchi, F.; Duvall, J.F.; Dittmer, J.; Mireskandari, A.; Reid, R.L.; Gitlin, S.D.; Brady, J.N. Involvement of transcription factor YB-1 in human T-cell lymphotropic virus type I basal gene expression. J. Virol. 1994, 68, 561–565. [Google Scholar] [CrossRef]
- Didier, D.K.; Schiffenbauer, J.; Woulfe, S.L.; Zacheis, M.; Schwartz, B.D. Characterization of the cDNA encoding a protein binding to the major histocompatibility complex class II Y box. Proc. Natl. Acad. Sci. USA 1988, 85, 7322–7326. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Li, X.; Zhao, J.; Wu, S.; Wu, H.; Gou, S. Overexpression of YBX1 promotes pancreatic ductal adenocarcinoma growth via the GSK3B/Cyclin D1/Cyclin E1 pathway. Mol. Ther. Oncolytics 2020, 17, 21–30. [Google Scholar] [CrossRef]
- Jurchott, K.; Bergmann, S.; Stein, U.; Walther, W.; Janz, M.; Manni, I.; Piaggio, G.; Fietze, E.; Dietel, M.; Royer, H.D. YB-1 as a cell cycle-regulated transcription factor facilitating cyclin A and cyclin B1 gene expression. J. Biol. Chem. 2003, 278, 27988–27996. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shen, D.; Li, S.; Li, Q.; Zuo, Q.; Lu, J.; Tang, D.; Feng, Y.; Yin, P.; Chen, C. LINC00665 activating Wnt3a/β-catenin signaling by bond with YBX1 promotes gastric cancer proliferation and metastasis. Cancer Gene Ther. 2023, 30, 1530–1542. [Google Scholar] [CrossRef] [PubMed]
- Berquin, I.M.; Pang, B.; Dziubinski, M.L.; Scott, L.M.; Chen, Y.Q.; Nolan, G.P.; Ethier, S.P. Y-box-binding protein 1 confers EGF independence to human mammary epithelial cells. Oncogene 2005, 24, 3177–3186. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Ma, Y.; Yong, L.; Yang, C.; Wang, P.; Liu, X.; Zhu, B.; Zhou, H.; Liu, X.; Liu, Z. Y-box binding protein-1 promotes tumorigenesis and progression via the epidermal growth factor receptor/AKT pathway in spinal chordoma. Cancer Sci. 2019, 110, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lee, C.; Yokom, D.; Jiang, H.; Cheang, M.C.; Yorida, E.; Turbin, D.; Berquin, I.M.; Mertens, P.R.; Iftner, T.; et al. Disruption of the Y-box binding protein-1 results in suppression of the epidermal growth factor receptor and HER-2. Cancer Res. 2006, 66, 4872–4879. [Google Scholar] [CrossRef]
- Sutherland, B.W.; Kucab, J.; Wu, J.; Lee, C.; Cheang, M.C.; Yorida, E.; Turbin, D.; Dedhar, S.; Nelson, C.; Pollak, M.; et al. Akt phosphorylates the Y-box binding protein 1 at Ser102 located in the cold shock domain and affects the anchorage-independent growth of breast cancer cells. Oncogene 2005, 24, 4281–4292. [Google Scholar] [CrossRef]
- Hussain, S.A.; Venkatesh, T. YBX1/lncRNA SBF2-AS1 interaction regulates proliferation and tamoxifen sensitivity via PI3K/AKT/MTOR signaling in breast cancer cells. Mol. Biol. Rep. 2023, 50, 3413–3428. [Google Scholar] [CrossRef]
- Zheng, C.; Wei, Y.; Zhang, Q.; Sun, M.; Wang, Y.; Hou, J.; Zhang, P.; Lv, X.; Su, D.; Jiang, Y. Multiomics analyses reveal DARS1-AS1/YBX1–controlled posttranscriptional circuits promoting glioblastoma tumorigenesis/radioresistance. Sci. Adv. 2023, 9, eadf3984. [Google Scholar] [CrossRef]
- Xie, N.; Zhang, R.; Bi, Z.; Ren, W.; You, K.; Hu, H.; Xu, Y.; Yao, H. H3K27 acetylation activated long noncoding RNA RP11-162G10. 5 promotes breast cancer progression via the YBX1/GLO1 axis. Cell Oncol. 2023, 46, 375–390. [Google Scholar] [CrossRef]
- Stratford, A.L.; Fry, C.J.; Desilets, C.; Davies, A.H.; Cho, Y.Y.; Li, Y.; Dong, Z.; Berquin, I.M.; Roux, P.P.; Dunn, S.E. Y-box binding protein-1 serine 102 is a downstream target of p90 ribosomal S6 kinase in basal-like breast cancer cells. Breast Cancer Res. 2008, 10, R99. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Watari, K.; Kawahara, A.; Sudo, T.; Hattori, S.; Murakami, Y.; Izumi, H.; Itou, J.; Toi, M.; Akiba, J.; et al. Targeting Phosphorylation of Y-Box-Binding Protein YBX1 by TAS0612 and Everolimus in Overcoming Antiestrogen Resistance. Mol. Cancer Ther. 2020, 19, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, L.; Mundade, R.; Wang, B.; Wei, H.; Hartley, A.V.; Martin, M.; McElyea, K.; Temm, C.J.; Sandusky, G.; Liu, Y.; et al. Critical role of phosphorylation of serine 165 of YBX1 on the activation of NF-κB in colon cancer. Oncotarget 2015, 6, 29396–29412. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Hua, L.; Wang, B.; Wei, H.; Prabhu, L.; Hartley, A.V.; Jiang, G.; Liu, Y.; Lu, T. Novel Serine 176 Phosphorylation of YBX1 Activates NF-κB in Colon Cancer. J. Biol. Chem. 2017, 292, 3433–3444. [Google Scholar] [CrossRef]
- Hartley, A.V.; Wang, B.; Mundade, R.; Jiang, G.; Sun, M.; Wei, H.; Sun, S.; Liu, Y.; Lu, T. PRMT5-mediated methylation of YBX1 regulates NF-κB activity in colorectal cancer. Sci. Rep. 2020, 10, 15934. [Google Scholar] [CrossRef]
- Frye, B.C.; Halfter, S.; Djudjaj, S.; Muehlenberg, P.; Weber, S.; Raffetseder, U.; En-Nia, A.; Knott, H.; Baron, J.M.; Dooley, S.; et al. Y-box protein-1 is actively secreted through a non-classical pathway and acts as an extracellular mitogen. EMBO Rep. 2009, 10, 783–789. [Google Scholar] [CrossRef]
- Rauen, T.; Raffetseder, U.; Frye, B.C.; Djudjaj, S.; Mühlenberg, P.J.; Eitner, F.; Lendahl, U.; Bernhagen, J.; Dooley, S.; Mertens, P.R. YB-1 acts as a ligand for Notch-3 receptors and modulates receptor activation. J. Biol. Chem. 2009, 284, 26928–26940. [Google Scholar] [CrossRef]
- Wu, S.L.; Fu, X.; Huang, J.; Jia, T.T.; Zong, F.Y.; Mu, S.R.; Zhu, H.; Yan, Y.; Qiu, S.; Wu, Q.; et al. Genome-wide analysis of YB-1-RNA interactions reveals a novel role of YB-1 in miRNA processing in glioblastoma multiforme. Nucleic Acids Res. 2015, 43, 8516–8528. [Google Scholar] [CrossRef]
- To, K.; Fotovati, A.; Reipas, K.M.; Law, J.H.; Hu, K.; Wang, J.; Astanehe, A.; Davies, A.H.; Lee, L.; Stratford, A.L.; et al. Y-box binding protein-1 induces the expression of CD44 and CD49f leading to enhanced self-renewal, mammosphere growth, and drug resistance. Cancer Res. 2010, 70, 2840–2851. [Google Scholar] [CrossRef]
- Evdokimova, V.; Tognon, C.; Ng, T.; Ruzanov, P.; Melnyk, N.; Fink, D.; Sorokin, A.; Ovchinnikov, L.P.; Davicioni, E.; Triche, T.J.; et al. Translational activation of snail1 and other developmentally regulated transcription factors by YB-1 promotes an epithelial-mesenchymal transition. Cancer Cell 2009, 15, 402–415. [Google Scholar] [CrossRef]
- Evdokimova, V.; Tognon, C.; Ng, T.; Sorensen, P.H. Reduced proliferation and enhanced migration: Two sides of the same coin? Molecular mechanisms of metastatic progression by YB-1. Cell Cycle 2009, 8, 2901–2906. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, D.; Chen, H.; Wang, W.; Liu, Y.; Wang, Y.; Duan, C.; Ning, Z.; Guo, X.; Otkur, W.; et al. YB1 regulates miR-205/200b-ZEB1 axis by inhibiting microRNA maturation in hepatocellular carcinoma. Cancer Commun. 2021, 41, 576–595. [Google Scholar] [CrossRef] [PubMed]
- Harbour, J.W.; Dean, D.C. The Rb/E2F pathway: Expanding roles and emerging paradigms. Genes. Dev. 2000, 14, 2393–2409. [Google Scholar] [CrossRef] [PubMed]
- Giacinti, C.; Giordano, A. RB and cell cycle progression. Oncogene 2006, 25, 5220–5227. [Google Scholar] [CrossRef]
- Basaki, Y.; Taguchi, K.; Izumi, H.; Murakami, Y.; Kubo, T.; Hosoi, F.; Watari, K.; Nakano, K.; Kawaguchi, H.; Ohno, S.; et al. Y-box binding protein-1 (YB-1) promotes cell cycle progression through CDC6-dependent pathway in human cancer cells. Eur. J. Cancer 2010, 46, 954–965. [Google Scholar] [CrossRef]
- Harada, M.; Kotake, Y.; Ohhata, T.; Kitagawa, K.; Niida, H.; Matsuura, S.; Funai, K.; Sugimura, H.; Suda, T.; Kitagawa, M. YB-1 promotes transcription of cyclin D1 in human non-small-cell lung cancers. Genes Cells 2014, 19, 504–516. [Google Scholar] [CrossRef]
- Wang, H.; Sun, R.; Gu, M.; Li, S.; Zhang, B.; Chi, Z.; Hao, L. shRNA-Mediated Silencing of Y-Box Binding Protein-1 (YB-1) Suppresses Growth of Neuroblastoma Cell SH-SY5Y In Vitro and In Vivo. PLoS ONE 2015, 10, e0127224. [Google Scholar] [CrossRef]
- Lindqvist, A.; Rodríguez-Bravo, V.; Medema, R.H. The decision to enter mitosis: Feedback and redundancy in the mitotic entry network. J. Cell Biol. 2009, 185, 193–202. [Google Scholar] [CrossRef]
- Terada, Y.; Tatsuka, M.; Jinno, S.; Okayama, H. Requirement for tyrosine phosphorylation of Cdk4 in G1 arrest induced by ultraviolet irradiation. Nature 1995, 376, 358–362. [Google Scholar] [CrossRef]
- Iavarone, A.; Massagué, J. Repression of the CDK activator Cdc25A and cell-cycle arrest by cytokine TGF-beta in cells lacking the CDK inhibitor p15. Nature 1997, 387, 417–422. [Google Scholar] [CrossRef]
- Bartek, J.; Lukas, J. Mammalian G1- and S-phase checkpoints in response to DNA damage. Curr. Opin. Cell Biol. 2001, 13, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Wang, Y.; Guo, T.; Yu, W.; Li, J.; Tang, Z.; Yu, Z.; Zhao, L.; Zhang, Y.; Wang, Z.; et al. YBX1 regulates tumor growth via CDC25a pathway in human lung adenocarcinoma. Oncotarget 2016, 7, 82139–82157. [Google Scholar] [CrossRef] [PubMed]
- Lasham, A.; Samuel, W.; Cao, H.; Patel, R.; Mehta, R.; Stern, J.L.; Reid, G.; Woolley, A.G.; Miller, L.D.; Black, M.A.; et al. YB-1, the E2F pathway, and regulation of tumor cell growth. J. Natl. Cancer Inst. 2012, 104, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, M.R.; Astanehe, A.; To, K.; Fotovati, A.; Davies, A.H.; Zhao, Y.; Jiang, H.; Stratford, A.L.; Shadeo, A.; Boccaccio, C.; et al. Profiling YB-1 target genes uncovers a new mechanism for MET receptor regulation in normal and malignant human mammary cells. Oncogene 2009, 28, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, D.; Fu, Q.; Wang, S.; Wang, Z.; Zhang, X.; Chen, X.; Zhu, X.; An, N.; Chen, Y. YBX1 as an oncogenic factor in T-cell acute lymphoblastic leukemia. Blood Adv. 2023, 7, 4874–4885. [Google Scholar] [CrossRef] [PubMed]
- Schelch, K.; Emminger, D.; Zitta, B.; Johnson, T.G.; Kopatz, V.; Eder, S.; Ries, A.; Stefanelli, A.; Heffeter, P.; Hoda, M.A. Targeting YB-1 via entinostat enhances cisplatin sensitivity of pleural mesothelioma in vitro and in vivo. Cancer Lett. 2023, 574, 216395. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Strasser, A.; McDunn, J.E.; Swanson, P.E. Cell death. N. Engl. J. Med. 2009, 361, 1570–1583. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene 2007, 26, 1324–1337. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. Bcl-2-regulated apoptosis: Mechanism and therapeutic potential. Curr. Opin. Immunol. 2007, 19, 488–496. [Google Scholar] [CrossRef]
- Lasham, A.; Lindridge, E.; Rudert, F.; Onrust, R.; Watson, J. Regulation of the human fas promoter by YB-1, Puralpha and AP-1 transcription factors. Gene 2000, 252, 1–13. [Google Scholar] [CrossRef]
- Bhullar, J.; Sollars, V.E. YBX1 expression and function in early hematopoiesis and leukemic cells. Immunogenetics 2011, 63, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Xie, X.; Han, G.; Zhang, T.; Li, Y.; Li, Y.; Yin, R.; Wang, Q.; Zhang, T.; Wang, P.; et al. YBX1 is required for maintaining myeloid leukemia cell survival by regulating BCL2 stability in an m6A-dependent manner. Blood 2021, 138, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Wang, C.; Liu, S.; Du, R.; Tian, S.; Chen, R.; Geng, H.; Subramanian, S.; Niu, Y.; Wang, Y.; et al. YBX1 knockdown induces renal cell carcinoma cell apoptosis via Kindlin-2. Cell Cycle 2021, 20, 2413–2427. [Google Scholar] [CrossRef] [PubMed]
- Nagasu, S.; Sudo, T.; Kinugasa, T.; Yomoda, T.; Fujiyoshi, K.; Shigaki, T.; Akagi, Y. Y-box-binding protein 1 inhibits apoptosis and upregulates EGFR in colon cancer. Oncol. Rep. 2019, 41, 2889–2896. [Google Scholar] [CrossRef] [PubMed]
- Kloetgen, A.; Duggimpudi, S.; Schuschel, K.; Hezaveh, K.; Picard, D.; Schaal, H.; Remke, M.; Klusmann, J.H.; Borkhardt, A.; McHardy, A.C.; et al. YBX1 Indirectly Targets Heterochromatin-Repressed Inflammatory Response-Related Apoptosis Genes through Regulating CBX5 mRNA. Int. J. Mol. Sci. 2020, 21, 4453. [Google Scholar] [CrossRef]
- Gong, H.; Gao, S.; Yu, C.; Li, M.; Liu, P.; Zhang, G.; Song, J.; Zheng, J. Effect and mechanism of YB-1 knockdown on glioma cell growth, migration, and apoptosis. Acta Biochim. Biophys. Sin. 2020, 52, 168–179. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, N.; Sato, F.; Tanimoto, K.; Okada, H.; Liu, Y.; Bhawal, U.K. The roles of Y-box-binding protein (YB)-1 and C-X-C motif chemokine ligand 14 (CXCL14) in the progression of prostate cancer via extracellular-signal-regulated kinase (ERK) signaling. Bioengineered 2021, 12, 9128–9139. [Google Scholar] [CrossRef]
- Shah, A.; Lindquist, J.A.; Rosendahl, L.; Schmitz, I.; Mertens, P.R. Novel insights into YB-1 signaling and cell death decisions. Cancers 2021, 13, 3306. [Google Scholar] [CrossRef]
- Li, Z.; Chen, H.; Li, B.; Wang, T.; Ji, S.; Qin, Y.; Xu, X.; Yu, X. Holistic anti-tumor resistance mechanism of YBX1 and its potential as a chemoresistance target in pancreatic ductal adenocarcinoma. Holist. Integr. Oncol. 2023, 2, 16. [Google Scholar] [CrossRef]
- Chai, J.; Wang, Q.; Qiu, Q.; Han, G.; Chen, Y.; Li, W.; Zhang, H. YBX1 regulates the survival of chronic myeloid leukemia stem cells by modulating m(6)A-mediated YWHAZ stability. Cell. Oncol. 2023, 46, 451–464. [Google Scholar] [CrossRef]
- Wu, Q.N.; Luo, X.J.; Liu, J.; Lu, Y.X.; Wang, Y.; Qi, J.; Liu, Z.X.; Huang, Q.T.; Liu, Z.K.; Lu, J.B.; et al. MYC-Activated LncRNA MNX1-AS1 Promotes the Progression of Colorectal Cancer by Stabilizing YB1. Cancer Res. 2021, 81, 2636–2650. [Google Scholar] [CrossRef] [PubMed]
- Fritzke, M.; Chen, K.; Tang, W.; Stinson, S.; Pham, T.; Wang, Y.; Xu, L.; Chen, E.Y. The MYC-YBX1 Circuit in Maintaining Stem-like Vincristine-Resistant Cells in Rhabdomyosarcoma. Cancers 2023, 15, 2788. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zeng, Y.; Yu, Z.; Chen, S.; Xie, J.; Rao, B.; Yang, B.; Qiu, F.; Lu, J.; Yang, L. EIF4a3-regulated circRABL2B regulates cell stemness and drug sensitivity of lung cancer via YBX1-dependent downregulation of MUC5AC expression. Int. J. Biol. Sci. 2023, 19, 2725. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, G.; Liu, B.; Tao, Z.; Wu, Y.; Zhang, K.; Feng, Z.; Huang, Y.; Wang, H. PLK1 inhibition promotes apoptosis and DNA damage in glioma stem cells by regulating the nuclear translocation of YBX1. Cell Death Discov. 2023, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.W.; Zhu, W.J.; Ma, R.R.; Tian, Y.R.; Chen, X.; Gao, P. PIN1P1 is activated by CREB1 and promotes gastric cancer progression via interacting with YBX1 and upregulating PIN1. J. Cell Mol. Med. 2023. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qi, Y.; Yu, J.; Hao, Y.; Xu, L.; Ding, X.; Zhang, M.; Geng, J. Aurora kinase A regulates cancer-associated RNA aberrant splicing in breast cancer. Heliyon 2023, 9, e17386. [Google Scholar] [CrossRef] [PubMed]
- Homer, C.; Knight, D.A.; Hananeia, L.; Sheard, P.; Risk, J.; Lasham, A.; Royds, J.A.; Braithwaite, A.W. Y-box factor YB1 controls p53 apoptotic function. Oncogene 2005, 24, 8314–8325. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Liu, P.; Wen, J.; Gu, Y.; Yang, Z.; Lan, J.; Fan, H.; Liu, Z.; Guo, D. FCN3 inhibits the progression of hepatocellular carcinoma by suppressing SBDS-mediated blockade of the p53 pathway. Int. J. Biol. Sci. 2023, 19, 362–376. [Google Scholar] [CrossRef]
- Meltendorf, S.; Fu, H.; Pierau, M.; Lindquist, J.A.; Finzel, S.; Mertens, P.R.; Gieseler-Halbach, S.; Ambach, A.; Thomas, U.; Lingel, H.; et al. Cell Survival Failure in Effector T Cells From Patients With Systemic Lupus Erythematosus Following Insufficient Up-Regulation of Cold-Shock Y-Box Binding Protein 1. Arthritis Rheumatol. 2020, 72, 1721–1733. [Google Scholar] [CrossRef]
- Holm, P.S.; Bergmann, S.; Jurchott, K.; Lage, H.; Brand, K.; Ladhoff, A.; Mantwill, K.; Curiel, D.T.; Dobbelstein, M.; Dietel, M.; et al. YB-1 relocates to the nucleus in adenovirus-infected cells and facilitates viral replication by inducing E2 gene expression through the E2 late promoter. J. Biol. Chem. 2002, 277, 10427–10434. [Google Scholar] [CrossRef]
- Glockzin, G.; Mantwill, K.; Jurchott, K.; Bernshausen, A.; Ladhoff, A.; Royer, H.D.; Gansbacher, B.; Holm, P.S. Characterization of the recombinant adenovirus vector AdYB-1: Implications for oncolytic vector development. J. Virol. 2006, 80, 3904–3911. [Google Scholar] [CrossRef] [PubMed]
- Birch, J.; Gil, J. Senescence and the SASP: Many therapeutic avenues. Genes Dev. 2020, 34, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef]
- Kwon, E.; Todorova, K.; Wang, J.; Horos, R.; Lee, K.K.; Neel, V.A.; Negri, G.L.; Sorensen, P.H.; Lee, S.W.; Hentze, M.W.; et al. The RNA-binding protein YBX1 regulates epidermal progenitors at a posttranscriptional level. Nat. Commun. 2018, 9, 1734. [Google Scholar] [CrossRef] [PubMed]
- Chiba, M.; Miyata, K.; Okawa, H.; Tanaka, Y.; Ueda, K.; Seimiya, H.; Takahashi, A. YBX1 Regulates Satellite II RNA Loading into Small Extracellular Vesicles and Promotes the Senescent Phenotype. Int. J. Mol. Sci. 2023, 24, 16399. [Google Scholar] [CrossRef]
- Avolio, R.; Inglés-Ferrándiz, M.; Ciocia, A.; Coll, O.; Bonnin, S.; Guitart, T.; Ribó, A.; Gebauer, F. Coordinated post-transcriptional control of oncogene-induced senescence by UNR/CSDE1. Cell Rep. 2022, 38, 110211. [Google Scholar] [CrossRef]
- Kotake, Y.; Ozawa, Y.; Harada, M.; Kitagawa, K.; Niida, H.; Morita, Y.; Tanaka, K.; Suda, T.; Kitagawa, M. YB1 binds to and represses the p16 tumor suppressor gene. Genes Cells 2013, 18, 999–1006. [Google Scholar] [CrossRef]
- Bisio, A.; Latorre, E.; Andreotti, V.; Bressac-de Paillerets, B.; Harland, M.; Scarra, G.B.; Ghiorzo, P.; Spitale, R.C.; Provenzani, A.; Inga, A. The 5′-untranslated region of p16INK4a melanoma tumor suppressor acts as a cellular IRES, controlling mRNA translation under hypoxia through YBX1 binding. Oncotarget 2015, 6, 39980–39994. [Google Scholar] [CrossRef]
- Xiao, Y.; Cai, G.P.; Feng, X.; Li, Y.J.; Guo, W.H.; Guo, Q.; Huang, Y.; Su, T.; Li, C.J.; Luo, X.H.; et al. Splicing factor YBX1 regulates bone marrow stromal cell fate during aging. EMBO J. 2023, 42, e111762. [Google Scholar] [CrossRef]
- Acosta, J.C.; Banito, A.; Wuestefeld, T.; Georgilis, A.; Janich, P.; Morton, J.P.; Athineos, D.; Kang, T.W.; Lasitschka, F.; Andrulis, M.; et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat. Cell Biol. 2013, 15, 978–990. [Google Scholar] [CrossRef]
- Coppé, J.P.; Patil, C.K.; Rodier, F.; Sun, Y.; Muñoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.Y.; Campisi, J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008, 6, 2853–2868. [Google Scholar] [CrossRef]
- Wang, B.; Kohli, J.; Demaria, M. Senescent Cells in Cancer Therapy: Friends or Foes? Trends Cancer 2020, 6, 838–857. [Google Scholar] [CrossRef] [PubMed]
- Faget, D.V.; Ren, Q.; Stewart, S.A. Unmasking senescence: Context-dependent effects of SASP in cancer. Nat. Rev. Cancer 2019, 19, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Montes, M.; Lubas, M.; Arendrup, F.S.; Mentz, B.; Rohatgi, N.; Tumas, S.; Harder, L.M.; Skanderup, A.J.; Andersen, J.S.; Lund, A.H. The long non-coding RNA MIR31HG regulates the senescence associated secretory phenotype. Nat. Commun. 2021, 12, 2459. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, P.; Zhang, J.; Peng, X.; Lu, Z.; Yu, S.; Meng, Y.; Tong, W.; Chen, J. Long noncoding RNA MIR31HG exhibits oncogenic property in pancreatic ductal adenocarcinoma and is negatively regulated by miR-193b. Oncogene 2016, 35, 3647–3657. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Xue, Y.; Yang, G.; Yin, S.; Shi, W.; Cheng, Y.; Yan, X.; Fan, S.; Zhang, H.; Zeng, F. Nanog RNA-binding proteins YBX1 and ILF3 affect pluripotency of embryonic stem cells. Cell Biol. Int. 2016, 40, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, J.; Astanehe, A.; Lee, C.; Fotovati, A.; Hu, K.; Dunn, S.E. The expression of activated Y-box binding protein-1 serine 102 mediates trastuzumab resistance in breast cancer cells by increasing CD44+ cells. Oncogene 2010, 29, 6294–6300. [Google Scholar] [CrossRef]
- Yang, F.; Cui, P.; Lu, Y.; Zhang, X. Requirement of the transcription factor YB-1 for maintaining the stemness of cancer stem cells and reverting differentiated cancer cells into cancer stem cells. Stem Cell Res. Ther. 2019, 10, 233. [Google Scholar] [CrossRef]
- Bledzka, K.; Schiemann, B.; Schiemann, W.P.; Fox, P.; Plow, E.F.; Sossey-Alaoui, K. The WAVE3-YB1 interaction regulates cancer stem cells activity in breast cancer. Oncotarget 2017, 8, 104072–104089. [Google Scholar] [CrossRef]
- Kim, H.J.; Choi, W.J.; Lee, C.H. Phosphorylation and reorganization of keratin networks: Implications for carcinogenesis and epithelial mesenchymal transition. Biomol. Ther. 2015, 23, 301. [Google Scholar] [CrossRef]
- Park, M.K.; Lee, H.; Lee, C.H. Post-translational modification of ZEB family members in cancer progression. Int. J. Mol. Sci. 2022, 23, 15127. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.-W.; Kim, C.-W.; Choi, K.-C. Epithelial-mesenchymal transition-inducing factors involved in the progression of lung cancers. Biomol. Ther. 2022, 30, 213. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, W.; Gao, G.; Song, Y.; Liu, H.; Li, L.; Zhou, J.; Yan, G.; Cui, H. Effect of FTY-720 on Pulmonary Fibrosis in Mice via the TGF-β1 Signaling Pathway and Autophagy. Biomol. Ther. 2023, 31, 434. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Chen, L.; Piao, S.; Wang, Y.; Li, Z.; Lin, Y.; Tang, X.; Zhang, H.; Zhang, H.; Wang, X. M6A RNA methylation-mediated RMRP stability renders proliferation and progression of non-small cell lung cancer through regulating TGFBR1/SMAD2/SMAD3 pathway. Cell Death Differ. 2023, 30, 605–617. [Google Scholar] [CrossRef]
- Bai, Y.; Gotz, C.; Chincarini, G.; Zhao, Z.; Slaney, C.; Boath, J.; Furic, L.; Angel, C.; Jane, S.M.; Phillips, W.A. YBX1 integration of oncogenic PI3K/mTOR signalling regulates the fitness of malignant epithelial cells. Nat. Commun. 2023, 14, 1591. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Li, Q.; Li, J.; Chen, K.; He, Q.; Zhao, Y.; Liang, Y.; Zhao, Y.; Qiao, H.; Liu, N.; et al. CircIPO7 Promotes Nasopharyngeal Carcinoma Metastasis and Cisplatin Chemoresistance by Facilitating YBX1 Nuclear Localization. Clin. Cancer Res. 2022, 28, 4521–4535. [Google Scholar] [CrossRef]
- Ban, Y.; Tan, Y.; Li, X.; Li, X.; Zeng, Z.; Xiong, W.; Li, G.; Xiang, B.; Yi, M. RNA-binding protein YBX1 promotes cell proliferation and invasiveness of nasopharyngeal carcinoma cells via binding to AURKA mRNA. J. Cancer 2021, 12, 3315–3324. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.L.; Ni, J.; Feng, W.T.; Yao, R.; Yue, S.; Zhu, Y.N.; Tang, H.Y.; Lv, L.Y.; Feng, J.F.; Zhu, W.G. High YBX1 expression indicates poor prognosis and promotes cell migration and invasion in nasopharyngeal carcinoma. Exp. Cell Res. 2017, 361, 126–134. [Google Scholar] [CrossRef]
- Lim, J.P.; Nair, S.; Shyamasundar, S.; Chua, P.J.; Muniasamy, U.; Matsumoto, K.; Gunaratne, J.; Bay, B.H. Silencing Y-box binding protein-1 inhibits triple-negative breast cancer cell invasiveness via regulation of MMP1 and beta-catenin expression. Cancer Lett. 2019, 452, 119–131. [Google Scholar] [CrossRef]
- Lu, J.; Li, X.; Wang, F.; Guo, Y.; Huang, Y.; Zhu, H.; Wang, Y.; Lu, Y.; Wang, Z. YB-1 expression promotes pancreatic cancer metastasis that is inhibited by microRNA-216a. Exp. Cell Res. 2017, 359, 319–326. [Google Scholar] [CrossRef]
- Kosnopfel, C.; Sinnberg, T.; Sauer, B.; Busch, C.; Niessner, H.; Schmitt, A.; Forchhammer, S.; Grimmel, C.; Mertens, P.R.; Hailfinger, S.; et al. YB-1 Expression and Phosphorylation Regulate Tumorigenicity and Invasiveness in Melanoma by Influencing EMT. Mol. Cancer Res. 2018, 16, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, J.; Wang, Y.; Fu, D.; Ideozu, J.E.; Geng, H.; Cui, Q.; Wang, C.; Chen, R.; Yu, Y.; et al. The interaction of YBX1 with G3BP1 promotes renal cell carcinoma cell metastasis via YBX1/G3BP1-SPP1-NF-κB signaling axis. J. Exp. Clin. Cancer Res. 2019, 38, 386. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.T.; Yu, Y.N.; Yip, G.W.; Matsumoto, K.; Bay, B.H. Silencing the YB-1 gene inhibits cell migration in gastric cancer in vitro. Anat. Rec. 2013, 296, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zheng, Y.; Jia, J.; Li, C.; Duan, Q.; Li, R.; Wang, X.; Shao, Y.; Chen, C.; Yan, H. Antimicrobial peptide LL-37 promotes the viability and invasion of skin squamous cell carcinoma by upregulating YB-1. Exp. Ther. Med. 2017, 14, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Narita, S.; Habuchi, T.; Eto, M. Validated prognostic significance of YB-1 genetic variation in metastatic prostate cancer. Pharmacogenomics J. 2021, 21, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Tailor, D.; Resendez, A.; Garcia-Marques, F.J.; Pandrala, M.; Going, C.C.; Bermudez, A.; Kumar, V.; Rafat, M.; Nambiar, D.K.; Honkala, A.; et al. Y box binding protein 1 inhibition as a targeted therapy for ovarian cancer. Cell Chem. Biol. 2021, 28, 1206–1220. [Google Scholar] [CrossRef]
- Liu, B.; Shen, H.; He, J.; Jin, B.; Tian, Y.; Li, W.; Hou, L.; Zhao, W.; Nan, J.; Zhao, J. Cytoskeleton remodeling mediated by circRNA-YBX1 phase separation suppresses the metastasis of liver cancer. Proc. Natl. Acad. Sci. USA 2023, 120, e2220296120. [Google Scholar] [CrossRef]
- Ferreira, A.; Bettencourt, M.; Alho, I.; Costa, A.; Sousa, A.; Mansinho, A.; Abreu, C.; Pulido, C.; Macedo, D.; Vendrell, I. Serum YB-1 (Y-box binding protein 1) as a biomarker of bone disease progression in patients with breast cancer and bone metastases. J. Bone Oncol. 2017, 6, 16–21. [Google Scholar] [CrossRef]
- Chen, S.; Li, K.; Guo, J.; Chen, H.N.; Ming, Y.; Jin, Y.; Xu, F.; Zhang, T.; Yang, Y.; Ye, Z.; et al. circNEIL3 inhibits tumor metastasis through recruiting the E3 ubiquitin ligase Nedd4L to degrade YBX1. Proc. Natl. Acad. Sci. USA 2023, 120, e2215132120. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, H.; Li, Z.; Li, F.; Liang, L.; Zou, Y.; Shen, H.; Li, J.; Xia, Y.; Cheng, Z.; et al. Circular RNA ACTN4 promotes intrahepatic cholangiocarcinoma progression by recruiting YBX1 to initiate FZD7 transcription. J. Hepatol. 2022, 76, 135–147. [Google Scholar] [CrossRef]
- Xie, Q.; Zhao, S.; Liu, W.; Cui, Y.; Li, F.; Li, Z.; Guo, T.; Yu, W.; Guo, W.; Deng, W.; et al. YBX1 Enhances Metastasis and Stemness by Transcriptionally Regulating MUC1 in Lung Adenocarcinoma. Front. Oncol. 2021, 11, 702491. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Yang, L.; Zou, Y.; Liang, J.Y.; Liu, P.; Gao, G.; Yang, A.; Tang, H.; Xie, X. Long non-coding RNA HUMT hypomethylation promotes lymphangiogenesis and metastasis via activating FOXK1 transcription in triple-negative breast cancer. J. Hematol. Oncol. 2020, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, J.; Li, Y.; Guo, W.; Chen, L.; Chen, M.; Chen, X.; Zhang, W.; Jin, X.; Jiang, M.; et al. CTPS1 promotes malignant progression of triple-negative breast cancer with transcriptional activation by YBX1. J. Transl. Med. 2022, 20, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, Y.C.; Gan, Y.; Teng, L.; Wang, L.; La, T.; Wang, P.; Gu, Y.; Yan, L.; Li, N.; et al. LncRNA MILIP links YBX1 to translational activation of Snai1 and promotes metastasis in clear cell renal cell carcinoma. J. Exp. Clin. Cancer Res. 2022, 41, 260. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, A.M.; Veinotte, C.J.; Cheng, H.; Grunewald, T.G.; Negri, G.L.; Somasekharan, S.P.; Corkery, D.P.; Tirode, F.; Mathers, J.; Khan, D.; et al. Translational Activation of HIF1α by YB-1 Promotes Sarcoma Metastasis. Cancer Cell 2015, 27, 682–697. [Google Scholar] [CrossRef] [PubMed]
- Cobbold, L.C.; Spriggs, K.A.; Haines, S.J.; Dobbyn, H.C.; Hayes, C.; de Moor, C.H.; Lilley, K.S.; Bushell, M.; Willis, A.E. Identification of internal ribosome entry segment (IRES)-trans-acting factors for the Myc family of IRESs. Mol. Cell. Biol. 2008, 28, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, R.H.; Bennagi, R.; Martin, J.; Phillips, A.O.; Redman, J.E.; Fraser, D.J. A conserved stem loop motif in the 5′untranslated region regulates transforming growth factor-β(1) translation. PLoS ONE 2010, 5, e12283. [Google Scholar] [CrossRef]
- Lv, J.; Liu, Y.; Mo, S.; Zhou, Y.; Chen, F.; Cheng, F.; Li, C.; Saimi, D.; Liu, M.; Zhang, H.; et al. Gasdermin E mediates resistance of pancreatic adenocarcinoma to enzymatic digestion through a YBX1-mucin pathway. Nat. Cell. Biol. 2022, 24, 364–372. [Google Scholar] [CrossRef]
- Xu, F.; Huang, M.; Chen, Q.; Niu, Y.; Hu, Y.; Hu, P.; Chen, D.; He, C.; Huang, K.; Zeng, Z.; et al. LncRNA HIF1A-AS1 Promotes Gemcitabine Resistance of Pancreatic Cancer by Enhancing Glycolysis through Modulating the AKT/YB1/HIF1α Pathway. Cancer Res. 2021, 81, 5678–5691. [Google Scholar] [CrossRef]
- Wong, C.H.; Lou, U.K.; Li, Y.; Chan, S.L.; Tong, J.H.; To, K.F.; Chen, Y. CircFOXK2 Promotes Growth and Metastasis of Pancreatic Ductal Adenocarcinoma by Complexing with RNA-Binding Proteins and Sponging MiR-942. Cancer Res. 2020, 80, 2138–2149. [Google Scholar] [CrossRef]
- Deng, S.J.; Chen, H.Y.; Ye, Z.; Deng, S.C.; Zhu, S.; Zeng, Z.; He, C.; Liu, M.L.; Huang, K.; Zhong, J.X.; et al. Hypoxia-induced LncRNA-BX111 promotes metastasis and progression of pancreatic cancer through regulating ZEB1 transcription. Oncogene 2018, 37, 5811–5828. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, L.; Li, J.; Chen, Y.; Wang, Y.; Zhang, Y.; Dong, Z.; Xue, W.; Sun, R.; Cui, G. NSUN2 stimulates tumor progression via enhancing TIAM2 mRNA stability in pancreatic cancer. Cell Death Discov. 2023, 9, 219. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Pan, Y.; Chen, H.; Lei, X.; Zhang, X. YBX1, Targeted By Microrna-382-5p, Promotes Laryngeal Squamous Cell Carcinoma Progression via Modulating RAS/MAPK Signaling. Recent Pat. Anticancer Drug Discov. 2023, 19, 176–187. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, J.; Yang, X.; Shan, J.; Zhang, Y.; Shi, X.; Wang, Y.; Chenyan, A.; Chang, J.; Wang, Y. KIF14 promotes proliferation, lymphatic metastasis and chemoresistance through G3BP1/YBX1 mediated NF-κB pathway in cholangiocarcinoma. Oncogene 2023, 42, 1392–1404. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; He, Y.; He, S.; Li, J.; Xia, J.; Chen, T.; Huo, L.; Ling, Y.; Liu, Q.; Zang, W. RP11-296E3. 2 acts as an important molecular chaperone for YBX1 and promotes colorectal cancer proliferation and metastasis by activating STAT3. J. Transl. Med. 2023, 21, 418. [Google Scholar] [CrossRef]
- Green, D.; Singh, A.; Tippett, V.L.; Tattersall, L.; Shah, K.M.; Siachisumo, C.; Ward, N.J.; Thomas, P.; Carter, S.; Jeys, L. YBX1-interacting small RNAs and RUNX2 can be blocked in primary bone cancer using CADD522. J. Bone Oncol. 2023, 39, 100474. [Google Scholar] [CrossRef]
- Kong, X.; Li, J.; Li, Y.; Duan, W.; Qi, Q.; Wang, T.; Yang, Q.; Du, L.; Mao, H.; Wang, C. A novel long non-coding RNA AC073352.1 promotes metastasis and angiogenesis via interacting with YBX1 in breast cancer. Cell Death Dis. 2021, 12, 670. [Google Scholar] [CrossRef]
- Potter, M.; Newport, E.; Morten, K.J. The Warburg effect: 80 years on. Biochem. Soc. Trans. 2016, 44, 1499–1505. [Google Scholar] [CrossRef]
- Lyssiotis, C.A.; Kimmelman, A.C. Metabolic Interactions in the Tumor Microenvironment. Trends Cell Biol. 2017, 27, 863–875. [Google Scholar] [CrossRef]
- Xu, L.; Li, H.; Wu, L.; Huang, S. YBX1 promotes tumor growth by elevating glycolysis in human bladder cancer. Oncotarget 2017, 8, 65946–65956. [Google Scholar] [CrossRef]
- Lai, Y.W.; Hsu, W.J.; Lee, W.Y.; Chen, C.H.; Tsai, Y.H.; Dai, J.Z.; Yang, C.C.; Lin, C.W. Prognostic Value of a Glycolytic Signature and Its Regulation by Y-Box-Binding Protein 1 in Triple-Negative Breast Cancer. Cells 2021, 10, 1890. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zhang, Q.; Yu, S.-K.; Nie, F.-Q.; Zhang, M.-L.; Wang, Q.; Lu, K.-H. THOC3 interacts with YBX1 to promote lung squamous cell carcinoma progression through PFKFB4 mRNA modification. Cell Death Dis. 2023, 14, 475. [Google Scholar] [CrossRef] [PubMed]
- Negrini, S.; Gorgoulis, V.G.; Halazonetis, T.D. Genomic instability—An evolving hallmark of cancer. Nat. Rev. Mol. Cell Biol. 2010, 11, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Salk, J.J.; Fox, E.J.; Loeb, L.A. Mutational heterogeneity in human cancers: Origin and consequences. Annu. Rev. Pathol. 2010, 5, 51–75. [Google Scholar] [CrossRef] [PubMed]
- Delicato, A.; Montuori, E.; Angrisano, T.; Pollice, A.; Calabrò, V. YB-1 Oncoprotein Controls PI3K/Akt Pathway by Reducing Pten Protein Level. Genes 2021, 12, 1551. [Google Scholar] [CrossRef] [PubMed]
- Ming, M.; He, Y.Y. PTEN in DNA damage repair. Cancer Lett. 2012, 319, 125–129. [Google Scholar] [CrossRef]
- Lasham, A.; Moloney, S.; Hale, T.; Homer, C.; Zhang, Y.F.; Murison, J.G.; Braithwaite, A.W.; Watson, J. The Y-box-binding protein, YB1, is a potential negative regulator of the p53 tumor suppressor. J. Biol. Chem. 2003, 278, 35516–35523. [Google Scholar] [CrossRef]
- Li, B.; Li, F.; Gu, T.; Guo, Y.; Shen, B.; Xu, X.; Shen, Z.; Chen, L.; Zhang, Q.; Dong, H.; et al. Specific knockdown of Y-box binding protein 1 in hepatic progenitor cells inhibits proliferation and alleviates liver fibrosis. Eur. J. Pharmacol. 2022, 921, 174866. [Google Scholar] [CrossRef]
- Tong, H.; Zhao, K.; Zhang, J.; Zhu, J.; Xiao, J. YB-1 modulates the drug resistance of glioma cells by activation of MDM2/p53 pathway. Drug Des. Dev. Ther. 2019, 13, 317–326. [Google Scholar] [CrossRef]
- Williams, A.B.; Schumacher, B. p53 in the DNA-Damage-Repair Process. Cold Spring Harb. Perspect. Med. 2016, 6, a026070. [Google Scholar] [CrossRef]
- Snowden, T.; Acharya, S.; Butz, C.; Berardini, M.; Fishel, R. hMSH4-hMSH5 recognizes Holliday Junctions and forms a meiosis-specific sliding clamp that embraces homologous chromosomes. Mol. Cell 2004, 15, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, X.; Dang, Y.; Li, D.; Lu, G.; Chan, W.Y.; Leung, P.C.K.; Zhao, S.; Qin, Y.; Chen, Z.J. Long noncoding RNA HCP5 participates in premature ovarian insufficiency by transcriptionally regulating MSH5 and DNA damage repair via YB1. Nucleic Acids Res. 2020, 48, 4480–4491. [Google Scholar] [CrossRef] [PubMed]
- Kishikawa, T.; Otsuka, M.; Yoshikawa, T.; Ohno, M.; Ijichi, H.; Koike, K. Satellite RNAs promote pancreatic oncogenic processes via the dysfunction of YBX1. Nat. Commun. 2016, 7, 13006. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Gao, C.; Xu, X.; Li, M.; Zhao, X.; Wang, Y.; Wang, Y.; Zhang, S.; Yan, Z.; Liu, X. FBL promotes cancer cell resistance to DNA damage and BRCA1 transcription via YBX1. EMBO Rep. 2023, 24, e56230. [Google Scholar] [CrossRef] [PubMed]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, A.; Sun, B.F.; Yang, Y.; Han, Y.N.; Yuan, X.; Chen, R.X.; Wei, W.S.; Liu, Y.; Gao, C.C.; et al. 5-methylcytosine promotes pathogenesis of bladder cancer through stabilizing mRNAs. Nat. Cell Biol. 2019, 21, 978–990. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Sun, B.F.; Chen, Y.S.; Xu, J.W.; Lai, W.Y.; Li, A.; Wang, X.; Bhattarai, D.P.; Xiao, W.; et al. 5-methylcytosine promotes mRNA export—NSUN2 as the methyltransferase and ALYREF as an m(5)C reader. Cell Res. 2017, 27, 606–625. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Wan, F.; Xu, W.; Liu, Z.; Wang, J.; Zhang, H.; Huang, S.; Ye, D. Positive epigenetic regulation loop between AR and NSUN2 promotes prostate cancer progression. Clin. Transl. Med. 2022, 12, e1028. [Google Scholar] [CrossRef]
- Liu, K.; Xu, P.; Lv, J.; Ge, H.; Yan, Z.; Huang, S.; Li, B.; Xu, H.; Yang, L.; Xu, Z.; et al. Peritoneal high-fat environment promotes peritoneal metastasis of gastric cancer cells through activation of NSUN2-mediated ORAI2 m5C modification. Oncogene 2023, 42, 1980–1993. [Google Scholar] [CrossRef]
- Jiang, D.; Qiu, T.; Peng, J.; Li, S.; Tala; Ren, W.; Yang, C.; Wen, Y.; Chen, C.H.; Sun, J.; et al. YB-1 is a positive regulator of KLF5 transcription factor in basal-like breast cancer. Cell Death Differ. 2022, 29, 1283–1295. [Google Scholar] [CrossRef]
- Yang, L.; Yin, H.; Chen, Y.; Pan, C.; Hang, H.; Lu, Y.; Ma, W.; Li, X.; Gan, W.; Guo, H.; et al. Low expression of PEBP1P2 promotes metastasis of clear cell renal cell carcinoma by post-transcriptional regulation of PEBP1 and KLF13 mRNA. Exp. Hematol. Oncol. 2022, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, J.; Su, Y.; Maimaitiyiming, Y.; Yang, S.; Shen, Z.; Lin, S.; Shen, S.; Zhan, G.; Wang, F.; et al. Distinct Roles of m(5)C RNA Methyltransferase NSUN2 in Major Gynecologic Cancers. Front. Oncol. 2022, 12, 786266. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, Y.; Sun, B.F.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.J.; Ping, X.L.; Chen, Y.S.; Wang, W.J.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.B. Dynamic m(6)A mRNA methylation directs translational control of heat shock response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef]
- Dominissini, D.; Nachtergaele, S.; Moshitch-Moshkovitz, S.; Peer, E.; Kol, N.; Ben-Haim, M.S.; Dai, Q.; Di Segni, A.; Salmon-Divon, M.; Clark, W.C.; et al. The dynamic N(1)-methyladenosine methylome in eukaryotic messenger RNA. Nature 2016, 530, 441–446. [Google Scholar] [CrossRef]
- Li, X.; Xiong, X.; Wang, K.; Wang, L.; Shu, X.; Ma, S.; Yi, C. Transcriptome-wide mapping reveals reversible and dynamic N(1)-methyladenosine methylome. Nat. Chem. Biol. 2016, 12, 311–316. [Google Scholar] [CrossRef]
- Li, D.; Li, K.; Zhang, W.; Yang, K.W.; Mu, D.A.; Jiang, G.J.; Shi, R.S.; Ke, D. The m6A/m5C/m1A Regulated Gene Signature Predicts the Prognosis and Correlates With the Immune Status of Hepatocellular Carcinoma. Front. Immunol. 2022, 13, 918140. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, F.; Lan, L.; Wen, N.; Li, H.; Sun, X. Diagnostic and prognostic value of m5C regulatory genes in hepatocellular carcinoma. Front. Genet. 2022, 13, 972043. [Google Scholar] [CrossRef]
- Yu, G.; Bao, J.; Zhan, M.; Wang, J.; Li, X.; Gu, X.; Song, S.; Yang, Q.; Liu, Y.; Wang, Z.; et al. Comprehensive Analysis of m5C Methylation Regulatory Genes and Tumor Microenvironment in Prostate Cancer. Front. Immunol. 2022, 13, 914577. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Yu, B.; Zhang, C.; Yi, P.; Li, H.; Xu, C.; Cao, L.; Chen, P.; Li, M.; Shen, K.; et al. Long noncoding RNA HOXC-AS3 indicates a poor prognosis and regulates tumorigenesis by binding to YBX1 in breast cancer. Am. J. Transl. Res. 2020, 12, 6335–6350. [Google Scholar] [PubMed]
- Su, H.; Fan, G.; Huang, J.; Qiu, X. LncRNA HOXC-AS3 promotes non-small-cell lung cancer growth and metastasis through upregulation of YBX1. Cell Death Dis. 2022, 13, 307. [Google Scholar] [CrossRef]
- Su, W.; Wang, L.; Zhao, H.; Hu, S.; Zhou, Y.; Guo, C.; Wu, B.; Li, L.; Yang, Z.; Beer, D.G.; et al. LINC00857 Interacting with YBX1 to Regulate Apoptosis and Autophagy via MET and Phosphor-AMPKa Signaling. Mol. Ther. Nucleic Acids 2020, 22, 1164–1175. [Google Scholar] [CrossRef]
- Du, J.; Zhang, G.; Qiu, H.; Yu, H.; Yuan, W. A novel positive feedback loop of linc02042 and c-Myc mediated by YBX1 promotes tumorigenesis and metastasis in esophageal squamous cell carcinoma. Cancer Cell Int. 2020, 20, 75. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, W.; Zhou, Y.; Hong, Z.; Ni, J.; Zhang, X.; Li, Z.; Li, M.; He, W.; Zhang, D.; et al. Therapeutic inhibition of GAS6-AS1/YBX1/MYC axis suppresses cell propagation and disease progression of acute myeloid leukemia. J. Exp. Clin. Cancer Res. 2021, 40, 353. [Google Scholar] [CrossRef]
- Chen, S.P.; Zhu, G.Q.; Xing, X.X.; Wan, J.L.; Cai, J.L.; Du, J.X.; Song, L.N.; Dai, Z.; Zhou, J. LncRNA USP2-AS1 Promotes Hepatocellular Carcinoma Growth by Enhancing YBX1-Mediated HIF1α Protein Translation Under Hypoxia. Front. Oncol. 2022, 12, 882372. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Niu, X.; Tang, M.; Li, J.; Song, B.; Guan, X. LncRNA BASP1-AS1 interacts with YBX1 to regulate Notch transcription and drives the malignancy of melanoma. Cancer Sci. 2021, 112, 4526–4542. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, Y.X.; Wang, D.L.; Yang, B.; Yan, H.Y.; Lin, L.H.; Li, Y.; Chen, J.; Xie, L.M.; Huang, Y.S.; et al. LncRNA DSCAM-AS1 interacts with YBX1 to promote cancer progression by forming a positive feedback loop that activates FOXA1 transcription network. Theranostics 2020, 10, 10823–10837. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.T.; Yan, Z.Y.; Xu, T.X.; Zhao, F.; Liu, L.; Li, F.; Guo, W. Reciprocal regulation of LINC00941 and SOX2 promotes progression of esophageal squamous cell carcinoma. Cell Death Dis. 2023, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiong, Q.; Chen, M.; Wang, B.; Yang, X.; Yang, M.; Wang, Q.; Cui, Z.; Ge, F. Long noncoding RNA HOTAIR interacts with Y-Box Protein-1 (YBX1) to regulate cell proliferation. Life Sci. Alliance 2021, 4, e202101139. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ye, Z.; Hou, L.; Zhang, X.; Liu, Z.; Wu, R.; Huang, F.; Wang, G.; Geng, X.; Zhao, H. Hepatitis B virus x gene-downregulated growth-arrest specific 5 inhibits the cell viability and invasion of hepatocellular carcinoma cell lines by activating Y-box-binding protein 1/p21 signaling. J. Cell Commun. Signal 2022, 16, 179–190. [Google Scholar] [CrossRef]
- Zhao, P.; Deng, Y.; Wu, Y.; Guo, Q.; Zhou, L.; Yang, X.; Wang, C. Long noncoding RNA SNHG6 promotes carcinogenesis by enhancing YBX1-mediated translation of HIF1α in clear cell renal cell carcinoma. FASEB J. 2021, 35, e21160. [Google Scholar] [CrossRef]
- Cong, Z.; Diao, Y.; Li, X.; Jiang, Z.; Xu, Y.; Zhou, H.; Qiang, Y.; Wu, H.; Shen, Y. Long non-coding RNA linc00665 interacts with YB-1 and promotes angiogenesis in lung adenocarcinoma. Biochem. Biophys. Res. Commun. 2020, 527, 545–552. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, J.; Fang, T.; Wang, X.; Wang, S.; Ma, Z.; Xu, Y.; Han, C.; Sun, M.; Xu, L.; et al. The long non-coding RNA PIK3CD-AS2 promotes lung adenocarcinoma progression via YBX1-mediated suppression of p53 pathway. Oncogenesis 2020, 9, 34. [Google Scholar] [CrossRef]
- Razpotnik, R.; Vidmar, R.; Fonović, M.; Rozman, D.; Režen, T. Circular RNA hsa_circ_0062682 Binds to YBX1 and Promotes Oncogenesis in Hepatocellular Carcinoma. Cancers 2022, 14, 4524. [Google Scholar] [CrossRef]
- Liu, P.; Zhong, Q.; Song, Y.; Guo, D.; Ma, D.; Chen, B.; Lan, J.; Liu, Q. Long noncoding RNA Linc01612 represses hepatocellular carcinoma progression by regulating miR-494/ATF3/p53 axis and promoting ubiquitination of YBX1. Int. J. Biol. Sci. 2022, 18, 2932–2948. [Google Scholar] [CrossRef]
- Deng, X.; Xiong, W.; Jiang, X.; Zhang, S.; Li, Z.; Zhou, Y.; Xiang, B.; Zhou, M.; Li, X.; Li, G.; et al. LncRNA LINC00472 regulates cell stiffness and inhibits the migration and invasion of lung adenocarcinoma by binding to YBX1. Cell Death Dis. 2020, 11, 945. [Google Scholar] [CrossRef]
- Diaz-Lagares, A.; Crujeiras, A.B.; Lopez-Serra, P.; Soler, M.; Setien, F.; Goyal, A.; Sandoval, J.; Hashimoto, Y.; Martinez-Cardús, A.; Gomez, A.; et al. Epigenetic inactivation of the p53-induced long noncoding RNA TP53 target 1 in human cancer. Proc. Natl. Acad. Sci. USA 2016, 113, E7535–E7544. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, L.; Zhao, K.; Lin, Q.; Li, H.; Xue, X.; Ge, W.; He, H.; Liu, D.; Xie, H.; et al. A novel LncRNA HITT forms a regulatory loop with HIF-1α to modulate angiogenesis and tumor growth. Cell Death Differ. 2020, 27, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Hong, H.; Xue, X.; Zhu, X.; Jiang, L.; Qin, M.; Liang, H.; Gao, L. A novel circular RNA, circFAT1(e2), inhibits gastric cancer progression by targeting miR-548g in the cytoplasm and interacting with YBX1 in the nucleus. Cancer Lett. 2019, 442, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lu, C.; Li, X.; Li, F.; Zhao, Y.; Xu, M.; Jia, H.; Yuan, S. LncRNA HOXA10-AS functions as an oncogene by binding miR-6509-5p to upregulate Y-box binding protein 1 in gastric cancer. Bioengineered 2022, 13, 11373–11387. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qian, J.; Yang, L. Inhibition of YBX1 by miR-216a Suppresses Proliferation and Invasion of Diffuse Large B-Cell Lymphoma. Balkan Med. J. 2021, 38, 171–176. [Google Scholar] [CrossRef]
- Johnson, T.G.; Schelch, K.; Cheng, Y.Y.; Williams, M.; Sarun, K.H.; Kirschner, M.B.; Kao, S.; Linton, A.; Klebe, S.; McCaughan, B.C.; et al. Dysregulated Expression of the MicroRNA miR-137 and Its Target YBX1 Contribute to the Invasive Characteristics of Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2018, 13, 258–272. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Z.; Wo, M.; Xu, W. The long non-coding RNA NNT-AS1 promotes clear cell renal cell carcinoma progression via regulation of the miR-137/ Y-box binding protein 1 axis. Bioengineered 2021, 12, 8994–9005. [Google Scholar] [CrossRef]
- Liu, S.L.; Sui, Y.F.; Lin, M.Z. MiR-375 is epigenetically downregulated due to promoter methylation and modulates multi-drug resistance in breast cancer cells via targeting YBX1. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3223–3229. [Google Scholar]
- Huang, S.; Zhang, X.; Guan, B.; Sun, P.; Hong, C.T.; Peng, J.; Tang, S.; Yang, J. A novel circular RNA hsa_circ_0008035 contributes to gastric cancer tumorigenesis through targeting the miR-375/YBX1 axis. Am. J. Transl. Res. 2019, 11, 2455–2462. [Google Scholar]
- Zheng, W.; Mu, H.; Chen, J.; Wang, C.; Hou, L. Circ_0002762 Regulates Oncoprotein YBX1 in Cervical Cancer via mir-375 to Regulate the Malignancy of Cancer Cells. Protein Pept. Lett. 2023, 30, 162–172. [Google Scholar] [CrossRef]
- Hanahan, D.; Folkman, J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 1996, 86, 353–364. [Google Scholar] [CrossRef]
- Lin, F.; Zeng, Z.; Song, Y.; Li, L.; Wu, Z.; Zhang, X.; Li, Z.; Ke, X.; Hu, X. YBX-1 mediated sorting of miR-133 into hypoxia/reoxygenation-induced EPC-derived exosomes to increase fibroblast angiogenesis and MEndoT. Stem Cell Res. Ther. 2019, 10, 263. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Zhao, Y.; Peng, H.; Zhang, N.; Bai, W. MEG3 sponges miRNA-376a and YBX1 to regulate angiogenesis in ovarian cancer endothelial cells. Heliyon 2023, 9, e13204. [Google Scholar] [CrossRef] [PubMed]
- Gopal, S.K.; Greening, D.W.; Mathias, R.A.; Ji, H.; Rai, A.; Chen, M.; Zhu, H.J.; Simpson, R.J. YBX1/YB-1 induces partial EMT and tumourigenicity through secretion of angiogenic factors into the extracellular microenvironment. Oncotarget 2015, 6, 13718–13730. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Huang, J.; Yu, K.; Chen, X.; He, Y.; Qi, D.; Wu, Y. YB-1 transferred by gastric cancer exosomes promotes angiogenesis via enhancing the expression of angiogenic factors in vascular endothelial cells. BMC Cancer 2020, 20, 996. [Google Scholar] [CrossRef] [PubMed]
- Quan, B.; Li, Z.; Yang, H.; Li, S.; Yan, X.; Wang, Y. The splicing factor YBX1 promotes the progression of osteosarcoma by upregulating VEGF165 and downregulating VEGF165b. Heliyon 2023, 9, e18706. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Nakamura, H.; Tanaka, Y.; Ohgi, S. Lung carcinoma: Analysis of T helper type 1 and 2 cells and T cytotoxic type 1 and 2 cells by intracellular cytokine detection with flow cytometry. Cancer 1999, 85, 2359–2367. [Google Scholar] [CrossRef]
- Nishimura, T.; Iwakabe, K.; Sekimoto, M.; Ohmi, Y.; Yahata, T.; Nakui, M.; Sato, T.; Habu, S.; Tashiro, H.; Sato, M.; et al. Distinct role of antigen-specific T helper type 1 (Th1) and Th2 cells in tumor eradication in vivo. J. Exp. Med. 1999, 190, 617–627. [Google Scholar] [CrossRef]
- Yao, D.; Zhang, X.; Wei, H.; Tian, Z. Antisense-induced blockade of GATA-3 expression could inhibit Th2 excursion of tumor cells in vitro and in vivo. Cell. Mol. Immunol. 2005, 2, 189–196. [Google Scholar]
- Che, X.; Liu, M.; Li, D.; Li, Z.; Guo, J.; Jia, R. RAN and YBX1 are required for cell proliferation and IL-4 expression and linked to poor prognosis in oral squamous cell carcinoma. Exp. Cell Res. 2021, 406, 112767. [Google Scholar] [CrossRef]
- Lv, Z.; Xue, C.; Zhang, L.; Sun, J.; Bo, C. Elevated mRNA Level of Y-Box Binding Protein 1 Indicates Unfavorable Prognosis Correlated with Macrophage Infiltration and T Cell Exhaustion in Luminal Breast Cancer. Cancer Manag. Res. 2021, 13, 6411–6428. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shao, C.; Shi, Y.; Han, W. Lessons learned from the blockade of immune checkpoints in cancer immunotherapy. J. Hematol. Oncol. 2018, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Bardhan, K.; Anagnostou, T.; Boussiotis, V.A. The PD1:PD-L1/2 Pathway from Discovery to Clinical Implementation. Front. Immunol. 2016, 7, 550. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Yu, S.; Qin, S.; Liu, Q.; Xu, H.; Zhao, W.; Chu, Q.; Wu, K. Gut microbiome modulates efficacy of immune checkpoint inhibitors. J. Hematol. Oncol. 2018, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Bao, L.; Tao, Z.; Chen, K. Flightless I Homolog Reverses Enzalutamide Resistance through PD-L1-Mediated Immune Evasion in Prostate Cancer. Cancer Immunol. Res. 2021, 9, 838–852. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Meng, L.; Liu, K.; Ji, B. Targeting the PD-L1/DNMT1 axis in acquired resistance to sorafenib in human hepatocellular carcinoma. Oncol. Rep. 2017, 38, 899–907. [Google Scholar] [CrossRef]
- Flavell, R.A.; Sanjabi, S.; Wrzesinski, S.H.; Licona-Limón, P. The polarization of immune cells in the tumour environment by TGFbeta. Nat. Rev. Immunol. 2010, 10, 554–567. [Google Scholar] [CrossRef]
- Fraser, D.J.; Phillips, A.O.; Zhang, X.; van Roeyen, C.R.; Muehlenberg, P.; En-Nia, A.; Mertens, P.R. Y-box protein-1 controls transforming growth factor-beta1 translation in proximal tubular cells. Kidney Int. 2008, 73, 724–732. [Google Scholar] [CrossRef]
- Wang, Y.; Su, J.; Fu, D.; Wang, Y.; Chen, Y.; Chen, R.; Qin, G.; Zuo, J.; Yue, D. The Role of YB1 in Renal Cell Carcinoma Cell Adhesion. Int. J. Med. Sci. 2018, 15, 1304–1311. [Google Scholar] [CrossRef]
- Ma, Y.; She, X.; Zhao, J.; Liu, S.; Li, C.; Wang, Q. Anoikis-related signature in liver hepatocellular carcinoma defines the YBX1/SPP1 axis by machine learning strategies and valid experiments. J. Gene Med. 2023, 25, e3516. [Google Scholar] [CrossRef] [PubMed]
- Ru, J.; Lu, J.; Ge, J.; Ding, B.; Su, R.; Jiang, Y.; Sun, Y.; Ma, J.; Li, Y.; Sun, J. IRGM is a novel regulator of PD-L1 via promoting S6K1-mediated phosphorylation of YBX1 in hepatocellular carcinoma. Cancer Lett. 2023, 581, 216495. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, S.; Nie, H.; Zhang, D.; Wang, T.; Sun, Q.; Wang, X.; Sun, Y. Promotion of colorectal cancer progression by immune-related lnc-SOX9–4 via suppression of YBX1 poly-ubiquitination and degradation. Cell. Signal. 2023, 111, 110854. [Google Scholar] [CrossRef] [PubMed]
- Zong, D.; Liu, X.; Li, J.; Ouyang, R.; Chen, P. The role of cigarette smoke-induced epigenetic alterations in inflammation. Epigenetics Chromatin 2019, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, H.; Ren, D.; Sun, Y.; Guo, F.; Cai, H.; Zhou, C.; Zhou, Y.; Jin, X.; Wu, H. CBX3 regulated By YBX1 promotes smoking-induced pancreatic cancer progression via inhibiting SMURF2 expression. Int. J. Biol. Sci. 2022, 18, 3484. [Google Scholar] [CrossRef]
- Schafer, Z.T.; Brugge, J.S. IL-6 involvement in epithelial cancers. J. Clin. Investig. 2007, 117, 3660–3663. [Google Scholar] [CrossRef]
- Chang, Q.; Bournazou, E.; Sansone, P.; Berishaj, M.; Gao, S.P.; Daly, L.; Wels, J.; Theilen, T.; Granitto, S.; Zhang, X.; et al. The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and metastasis. Neoplasia 2013, 15, 848–862. [Google Scholar] [CrossRef]
- Herzog, C.; Polanetzki, A.; Frey, S.; Schnoeder, T.M.; Wolleschak, D.; Mashamba, N.; Thangapandi, V.R.; Jentsch-Ullrich, K.; Kahl, C.; Schroeder, N. Inflammation Induced Expression of Ybx1 Mediates Resistance to JAK-Kinase Inhibitor Therapy in Myeloproliferative Neoplasia. Blood 2014, 124, 161. [Google Scholar] [CrossRef]
- Castellana, B.; Aasen, T.; Moreno-Bueno, G.; Dunn, S.E.; Ramón y Cajal, S. Interplay between YB-1 and IL-6 promotes the metastatic phenotype in breast cancer cells. Oncotarget 2015, 6, 38239–38256. [Google Scholar] [CrossRef]
- Huang, R.; Wang, S.; Wang, N.; Zheng, Y.; Zhou, J.; Yang, B.; Wang, X.; Zhang, J.; Guo, L.; Wang, S.; et al. CCL5 derived from tumor-associated macrophages promotes prostate cancer stem cells and metastasis via activating β-catenin/STAT3 signaling. Cell Death Dis. 2020, 11, 234. [Google Scholar] [CrossRef]
- Gao, L.F.; Zhong, Y.; Long, T.; Wang, X.; Zhu, J.X.; Wang, X.Y.; Hu, Z.Y.; Li, Z.G. Tumor bud-derived CCL5 recruits fibroblasts and promotes colorectal cancer progression via CCR5-SLC25A24 signaling. J. Exp. Clin. Cancer Res. 2022, 41, 81. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yao, Z.; Wang, J.; Zhang, W.; Yang, Y.; Zhang, Y.; Qu, X.; Zhu, Y.; Zou, J.; Peng, S.; et al. Macrophage-derived CCL5 facilitates immune escape of colorectal cancer cells via the p65/STAT3-CSN5-PD-L1 pathway. Cell Death Differ. 2020, 27, 1765–1781. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Fehr, A.; Brandt, S.; Jerchel, S.; Ballhause, T.M.; Philipsen, L.; Stolze, S.; Geffers, R.; Weng, H.; Fischer, K.D.; et al. Inflammatory cell infiltration and resolution of kidney inflammation is orchestrated by the cold-shock protein Y-box binding protein-1. Kidney Int. 2017, 92, 1157–1177. [Google Scholar] [CrossRef] [PubMed]
- Winkler, F.; Venkatesh, H.S.; Amit, M.; Batchelor, T.; Demir, I.E.; Deneen, B.; Gutmann, D.H.; Hervey-Jumper, S.; Kuner, T.; Mabbott, D. Cancer neuroscience: State of the field, emerging directions. Cell 2023, 186, 1689–1707. [Google Scholar] [CrossRef]
- Nguyen, T.M.; Ngoc, D.T.M.; Choi, J.-H.; Lee, C.-H. Unveiling the Neural Environment in Cancer: Exploring the Role of Neural Circuit Players and Potential Therapeutic Strategies. Cells 2023, 12, 1996. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Cho, J.; Lee, K. Tumour regression via integrative regulation of neurological, inflammatory, and hypoxic tumour microenvironment. Biomol. Ther. 2020, 28, 119. [Google Scholar] [CrossRef]
- Do, P.A.; Lee, C.H. The role of CDK5 in tumours and tumour microenvironments. Cancers 2020, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Spitkovsky, D.D.; Royer-Pokora, B.; Delius, H.; Kisseljov, F.; Jenkins, N.A.; Gilbert, D.J.; Copeland, N.G.; Royer, H.-D. Tissue restricted expression and chromosomal localization of the YB-1 gene encoding a 42kD nuclear CCAAT binding protein. Nucleic Acids Res. 1992, 20, 797–803. [Google Scholar] [CrossRef]
- Unkrüer, B.; Pekcec, A.; Fuest, C.; Wehmeyer, A.; Balda, M.S.; Horn, A.; Baumgärtner, W.; Potschka, H. Cellular localization of Y-box binding protein 1 in brain tissue of rats, macaques, and humans. BMC Neurosci. 2009, 10, 28. [Google Scholar] [CrossRef]
- Zhang, S.-B.; Lin, S.-Y.; Liu, M.; Liu, C.-C.; Ding, H.-H.; Sun, Y.; Ma, C.; Guo, R.-X.; Lv, Y.-Y.; Wu, S.-L. CircAnks1a in the spinal cord regulates hypersensitivity in a rodent model of neuropathic pain. Nat. Commun. 2019, 10, 4119. [Google Scholar] [CrossRef]
- Evans, M.K.; Matsui, Y.; Xu, B.; Willis, C.; Loome, J.; Milburn, L.; Fan, Y.; Pagala, V.; Peng, J.C. Ybx1 fine-tunes PRC2 activities to control embryonic brain development. Nat. Commun. 2020, 11, 4060. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; He, J.; Lin, L.; Li, Y.; Xia, Y. Neural Stem Cell Extracellular Vesicles Carrying YBX1 Inhibited Neuronal Pyroptosis Through Increasing m6A-modified GPR30 Stability and Expression in Ischemic Stroke. Transl. Stroke Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Tuerxun, T.; Li, X.; Hou, F.; Wang, Y.; Wang, X.; Ma, L. YBX1 Protects against Apoptosis Induced by Oxygen-Glucose Deprivation/Reoxygenation in PC12 Cells via Activation of the AKT/GSK3β Pathway. Folia Biol. 2021, 67, 150–157. [Google Scholar] [CrossRef]
- La Cognata, V.; Gentile, G.; Aronica, E.; Cavallaro, S. Splicing players are differently expressed in sporadic amyotrophic lateral sclerosis molecular clusters and brain regions. Cells 2020, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Abrakhi, S.; Kretov, D.A.; Desforges, B.; Dobra, I.; Bouhss, A.; Pastré, D.; Hamon, L. Nanoscale analysis reveals the maturation of neurodegeneration-associated protein aggregates: Grown in mRNA granules then released by stress granule proteins. ACS Nano 2017, 11, 7189–7200. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.K.; Polisetty, R.V.; Sharma, R.; Ganesh, R.A.; Gowda, H.; Purohit, A.K.; Ankathi, P.; Prasad, K.; Mariswamappa, K.; Lakshmikantha, A. Altered transcriptional regulatory proteins in glioblastoma and YBX1 as a potential regulator of tumor invasion. Sci. Rep. 2019, 9, 10986. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Fotovati, A.; Lee, C.; Wang, M.; Cote, G.; Guns, E.; Toyota, B.; Faury, D.; Jabado, N.; Dunn, S.E. Inhibition of Y-box binding protein-1 slows the growth of glioblastoma multiforme and sensitizes to temozolomide independent O 6-methylguanine-DNA methyltransferase. Mol. Cancer Ther. 2009, 8, 3276–3284. [Google Scholar] [CrossRef]
- Tamim, S.; Vo, D.T.; Uren, P.J.; Qiao, M.; Bindewald, E.; Kasprzak, W.K.; Shapiro, B.A.; Nakaya, H.I.; Burns, S.C.; Araujo, P.R. Genomic analyses reveal broad impact of miR-137 on genes associated with malignant transformation and neuronal differentiation in glioblastoma cells. PLoS ONE 2014, 9, e85591. [Google Scholar] [CrossRef]
- Wang, J.-Z.; Zhu, H.; You, P.; Liu, H.; Wang, W.-K.; Fan, X.; Yang, Y.; Xu, K.; Zhu, Y.; Li, Q. Upregulated YB-1 protein promotes glioblastoma growth through a YB-1/CCT4/mLST8/mTOR pathway. J. Clin. Investig. 2022, 132, e146536. [Google Scholar] [CrossRef]
- Ou, Y.; Zhao, Z.; Zhang, W.; Wu, Q.; Wu, C.; Liu, X.; Fu, M.; Ji, N.; Wang, D.; Qiu, J. Kindlin-2 interacts with β-catenin and YB-1 to enhance EGFR transcription during glioma progression. Oncotarget 2016, 7, 74872. [Google Scholar] [CrossRef]
- Zhang, X.; Ding, Z.; Mo, J.; Sang, B.; Shi, Q.; Hu, J.; Xie, S.; Zhan, W.; Lu, D.; Yang, M. GOLPH3 promotes glioblastoma cell migration and invasion via the mTOR-YB1 pathway in vitro. Mol. Carcinog. 2015, 54, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Robitaille, M.; Remke, M.; Maier, C.; Malhotra, A.; Gregorieff, A.; Wrana, J.L.; Taylor, M.D.; Angers, S.; Kenney, A.M. YB-1 is elevated in medulloblastoma and drives proliferation in Sonic hedgehog-dependent cerebellar granule neuron progenitor cells and medulloblastoma cells. Oncogene 2016, 35, 4256–4268. [Google Scholar] [CrossRef] [PubMed]
- McSwain, L.F.; Pillsbury, C.E.; Haji-Seyed-Javadi, R.; Rath, S.K.; Chen, V.; Huang, T.; Shahab, S.W.; Kunhiraman, H.; Ross, J.; Price, G.A. YB1 modulates the DNA damage response in medulloblastoma. Sci. Rep. 2023, 13, 8087. [Google Scholar] [CrossRef] [PubMed]
- Demir, I.E.; Friess, H.; Ceyhan, G.O. Neural plasticity in pancreatitis and pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.-Z.; Chen, C.-T.; Li, N.-C.; Lin, L.-C.; Huang, B.-S.; Chang, Y.-H.; Chow, L.-P. Y-box binding protein-1 promotes epithelial-mesenchymal transition in sorafenib-resistant hepatocellular carcinoma cells. Int. J. Mol. Sci. 2020, 22, 224. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.-H.; Zheng, B.; Li, Y.-H.; Sun, Y.; Han, A.-L.; Zhang, X.-H.; Lv, X.-R.; Chen, S.; Wen, J.-K. Novel insight into Y-box binding protein 1 in the regulation of vascular smooth muscle cell proliferation through targeting GC box-dependent genes. FEBS Lett. 2013, 587, 1326–1332. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, L.; Zhao, H.; Li, Q.; Hu, R.; Wang, H. Integrin β8 facilitates tumor growth and drug resistance through a Y-box binding protein 1-dependent signaling pathway in bladder cancer. Cancer Sci. 2020, 111, 2423–2430. [Google Scholar] [CrossRef] [PubMed]
- Kosnopfel, C.; Sinnberg, T.; Schittek, B. Y-box binding protein 1–a prognostic marker and target in tumour therapy. Eur. J. Cell Biol. 2014, 93, 61–70. [Google Scholar] [CrossRef]
- Zhang, C.; Yin, T.; Tao, R.; Xiao, B.; Chen, J.; Li, Z.; Miao, X.; Peng, Q.; Sun, L.; Zhang, W.; et al. Elevated nuclear YBX1 expression and the clinicopathological characteristics of patients with solid tumors: A meta-analysis. Cancer Manag. Res. 2019, 11, 4391–4402. [Google Scholar] [CrossRef]
- Yin, Q.; Zheng, M.; Luo, Q.; Jiang, D.; Zhang, H.; Chen, C. YB-1 as an oncoprotein: Functions, regulation, post-translational modifications, and targeted therapy. Cells 2022, 11, 1217. [Google Scholar] [CrossRef]
- Disis, M.; Liu, Y.; Stanton, S.; Gwin, W.; Coveler, A.; Liao, J.; Childs, J.; Cecil, D. 546 A phase I dose escalation study of STEMVAC, a multi-antigen, multi-epitope Th1 selective plasmid-based vaccine, targeting stem cell associated proteins in patients with advanced breast cancer. J. Immunother. Cancer 2022, 10, A571. [Google Scholar] [CrossRef]
- Kuwano, M.; Shibata, T.; Watari, K.; Ono, M. Oncogenic Y-box binding protein-1 as an effective therapeutic target in drug-resistant cancer. Cancer Sci. 2019, 110, 1536–1543. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, Q.; Wang, B.; Yuan, S.; Wang, X.; Li, K. Quercetin reversed MDR in breast cancer cells through down-regulating P-gp expression and eliminating cancer stem cells mediated by YB-1 nuclear translocation. Phytother. Res. 2018, 32, 1530–1536. [Google Scholar] [CrossRef] [PubMed]
- Kretov, D.A.; Mordovkina, D.A.; Eliseeva, I.A.; Lyabin, D.N.; Polyakov, D.N.; Joshi, V.; Desforges, B.; Hamon, L.; Lavrik, O.I.; Pastré, D. Inhibition of transcription induces phosphorylation of YB-1 at Ser102 and its accumulation in the nucleus. Cells 2019, 9, 104. [Google Scholar] [CrossRef] [PubMed]
- Chibi, M.; Meyer, M.; Skepu, A.; Rees, D.J.G.; Moolman-Smook, J.C.; Pugh, D.J. RBBP6 interacts with multifunctional protein YB-1 through its RING finger domain, leading to ubiquitination and proteosomal degradation of YB-1. J. Mol. Biol. 2008, 384, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Chen, L.; Lin, L.; Yang, M.; Li, T.; Wei, H.; Sha, C.; Xing, J.; Zhang, M.; Zhao, S. SIAH1 reverses chemoresistance in epithelial ovarian cancer via ubiquitination of YBX-1. Oncogenesis 2022, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Schnoeder, T.M.; Perner, F.; Heidel, F.H. A JAK of all trades: How global phosphoproteomics reveal the Achilles heel of MPNs. Mol. Cell. Oncol. 2021, 8, 1871172. [Google Scholar] [CrossRef]
- Maurya, P.K.; Mishra, A.; Yadav, B.S.; Singh, S.; Kumar, P.; Chaudhary, A.; Srivastava, S.; Murugesan, S.N.; Mani, A. Role of Y Box Protein-1 in cancer: As potential biomarker and novel therapeutic target. J. Cancer 2017, 8, 1900. [Google Scholar] [CrossRef]
- Khan, M.I.; Adhami, V.M.; Lall, R.K.; Sechi, M.; Joshi, D.C.; Haidar, O.M.; Syed, D.N.; Siddiqui, I.A.; Chiu, S.Y.; Mukhtar, H. YB-1 expression promotes epithelial-to-mesenchymal transition in prostate cancer that is inhibited by a small molecule fisetin. Oncotarget 2014, 5, 2462–2474. [Google Scholar] [CrossRef]
- Sechi, M.; Lall, R.K.; Afolabi, S.O.; Singh, A.; Joshi, D.C.; Chiu, S.Y.; Mukhtar, H.; Syed, D.N. Fisetin targets YB-1/RSK axis independent of its effect on ERK signaling: Insights from in vitro and in vivo melanoma models. Sci. Rep. 2018, 8, 15726. [Google Scholar] [CrossRef]
- Fang, L.; Velema, W.A.; Lee, Y.; Xiao, L.; Mohsen, M.G.; Kietrys, A.M.; Kool, E.T. Pervasive transcriptome interactions of protein-targeted drugs. Nat. Chem. 2023, 15, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Gunasekaran, V.P.; Nishi, K.; Sivakumar, D.; Sivaraman, T.; Mathan, G. Identification of 2,4-dihydroxy-5-pyrimidinyl imidothiocarbomate as a novel inhibitor to Y box binding protein-1 (YB-1) and its therapeutic actions against breast cancer. Eur. J. Pharm. Sci. 2018, 116, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, H.; Yang, Y.; Liu, J.; Chen, Z. Sesquiterpene lactone 6-O-angeloylplenolin reverses vincristine resistance by inhibiting YB-1 nuclear translocation in colon carcinoma cells. Oncol. Lett. 2018, 15, 9673–9680. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Saito, H.; Miyairi, S.; Kobayashi, S. 7-Hydorxyindirubin is capable of specifically inhibiting anticancer drug-induced YB-1 nuclear translocation without showing cytotoxicity in HepG2 hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2021, 544, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Shim, S.; Kim, Y.H.; Kim, M.J.; Park, S.; Myung, J.K. Inhibition of Y Box Binding Protein 1 Suppresses Cell Growth and Motility in Colorectal Cancer. Mol. Cancer Ther. 2020, 19, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.W.; Hung, C.M.; Lin, Y.C.; Ho, C.T.; Kao, J.Y.; Way, T.D. Aloe-emodin inhibits HER-2 expression through the downregulation of Y-box binding protein-1 in HER-2-overexpressing human breast cancer cells. Oncotarget 2016, 7, 58915–58930. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shrestha, N.; Préat, V.; Beloqui, A. Overcoming the intestinal barrier: A look into targeting approaches for improved oral drug delivery systems. J. Control. Release 2020, 322, 486–508. [Google Scholar] [CrossRef] [PubMed]
- Fischer, F.; Schumacher, A.; Meyer, N.; Fink, B.; Bauer, M.; Stojanovska, V.; Zenclussen, A.C. An old friend with a new face: YB-1 and its role in healthy pregnancy and pregnancy-associated complications. Front. Cell Dev. Biol. 2022, 10, 1039206. [Google Scholar] [CrossRef]
- Bader, A.G.; Felts, K.A.; Jiang, N.; Chang, H.W.; Vogt, P.K. Y box-binding protein 1 induces resistance to oncogenic transformation by the phosphatidylinositol 3-kinase pathway. Proc. Natl. Acad. Sci. USA 2003, 100, 12384–12389. [Google Scholar] [CrossRef]
- Bader, A.G.; Vogt, P.K. Inhibition of protein synthesis by Y box-binding protein 1 blocks oncogenic cell transformation. Mol. Cell. Biol. 2005, 25, 2095–2106. [Google Scholar] [CrossRef]


| Study Title | Clinical Trial ID | Drug in Trial | State | Aim | Disease |
|---|---|---|---|---|---|
| Ruxo-BEAT | 2013-002132-25 | Ruxolitinib | Ongoing | To prospectively validate Ybx1 as a diagnostic and putative prognostic marker for MPN (PV and ET) within the Ruxo-BEAT trial | Myeloproliferative neoplasms (MPN) including essential thrombocythemia (ET), polycythemia vera (PV) |
| STEMVAC in Patients with Early-Stage Triple-Negative Breast Cancer | NCT05455658 | STEMVAC DNA Plasmid-Based Vaccine (STEMVAC) Encoding Th1 Selective Epitopes from Five Antigens Associated with Breast Cancer Stem Cells (MDM2, YBX1, SOX2, CDH3, CD105) | Phase 2 | To test the immune system’s response to STEMVAC | Patients with stage IB-III triple-negative breast cancer |
| A Multiple Antigen Vaccine (STEMVAC) for the Treatment of Patients with Stage IV Non-Squamous Non-Small-Cell Lung Cancer | NCT05242965 | STEMVAC | Phase 2 | This phase II trial tests whether STEMVAC polyepitope plasmid DNA vaccine works to shrink tumors | Lung Non-Squamous Non-Small-Cell Carcinoma Stage IV Lung Cancer AJCC v8 |
| Vaccine Therapy in Treating Patients with HER2-Negative Stage III-IV Breast Cancer | NCT02157051 | STEMVAC | Phase 1 | To study the side effects and best dose of multiantigen deoxyribonucleic acid (DNA) plasmid-based vaccine STEMVAC in treating patients with HER2-negative stage III-IV breast cancer | HER2-negative stage III-IV breast cancer |
| Name | Cancer Type | Mechanism | Regimen | Animal Study | Reference |
|---|---|---|---|---|---|
| Azopodo-phyllotoxin SU056 | Ovarian cancer | YBX1 inhibitor | Alone 20 mg/kg daily or 10 mg/kg daily + palitaxel 5 mg/kg weekly | Xenograft | [116] |
| Fisetin | Prostate cancer Melanoma | Bind to YBX1 and hinder YBX1 from interacting with AKT Inhibit RSK kinase activity Inhibit EMT and MDR1 | 1mg Fisetin/animal intraperitoneally compared to 2 mg Vemurafenib/animal orally | Xenograft | [269,270] |
| 2,4-dihydroxy-5-pyrimidinyl imidothiocarbomate (DPI) | Breast cancer | Inhibit YBX1 nuclear translocation | 110.1 μM | Chick chorioallantoic membrane (CAM) xenograft | [272] |
| 7-hydroxyindirubin | Hepatocellular carcinoma | Inhibit Actinomycin D-induced YBX1 nuclear translocation | 7-hydroxyindirubin 5 μM Actinomycin D 65 nM | No | [274] |
| Sesquiterpene lactone 6-O-angeloylplenolin | Colon carcinoma HCT-8 | Reverse Vincristine resistance by inhibiting YBX1 nuclear translocation | 3.5–7 mg/kg/day orally Vincristine 2.5 mg/kg/day orally | Xenograft | [273] |
| TAS0612 | Triple-negative breast cancer | Suppress YBX1 expression by inhibiting multikinase (AKT, p70S6K, and p90RSK) | 50 mg/kg | Xenograft | [32] |
| BEZ235 | Colorectal cancer | Suppress YBX1 expression through inhibiting AKT/mTOr, thus enhancing the cytotoxicity of radiotherapy | 50 mg/kg once daily | Intrahepatic tumorigenesis | [275] |
| Aloe-emodin | HER2-positive breast cancer | Suppress YBX1 expression by inhibiting Twist and ILK/ATK/mTor signaling | 12.5–50 mg/kg | Xenograft | [276] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinh, N.T.M.; Nguyen, T.M.; Park, M.K.; Lee, C.H. Y-Box Binding Protein 1: Unraveling the Multifaceted Role in Cancer Development and Therapeutic Potential. Int. J. Mol. Sci. 2024, 25, 717. https://doi.org/10.3390/ijms25020717
Dinh NTM, Nguyen TM, Park MK, Lee CH. Y-Box Binding Protein 1: Unraveling the Multifaceted Role in Cancer Development and Therapeutic Potential. International Journal of Molecular Sciences. 2024; 25(2):717. https://doi.org/10.3390/ijms25020717
Chicago/Turabian StyleDinh, Ngoc Thi Minh, Tuan Minh Nguyen, Mi Kyung Park, and Chang Hoon Lee. 2024. "Y-Box Binding Protein 1: Unraveling the Multifaceted Role in Cancer Development and Therapeutic Potential" International Journal of Molecular Sciences 25, no. 2: 717. https://doi.org/10.3390/ijms25020717
APA StyleDinh, N. T. M., Nguyen, T. M., Park, M. K., & Lee, C. H. (2024). Y-Box Binding Protein 1: Unraveling the Multifaceted Role in Cancer Development and Therapeutic Potential. International Journal of Molecular Sciences, 25(2), 717. https://doi.org/10.3390/ijms25020717