

Function of Presynaptic Inhibitory Cannabinoid CB1 Receptors in Spontaneously Hypertensive Rats and Its Modification by Enhanced Endocannabinoid Tone


Abstract
:
1. Introduction
2. Results
2.1. General
2.2. Effects of the Cannabinoid Receptor Agonist CP55940 in the Presence and Absence of the CB1 Receptor Antagonist AM251

2.3. Effects of the Endocannabinoid Degradation Enzyme Inhibitors URB597, MJN110 and JZL195 and Their Influence on the CP55940 Action

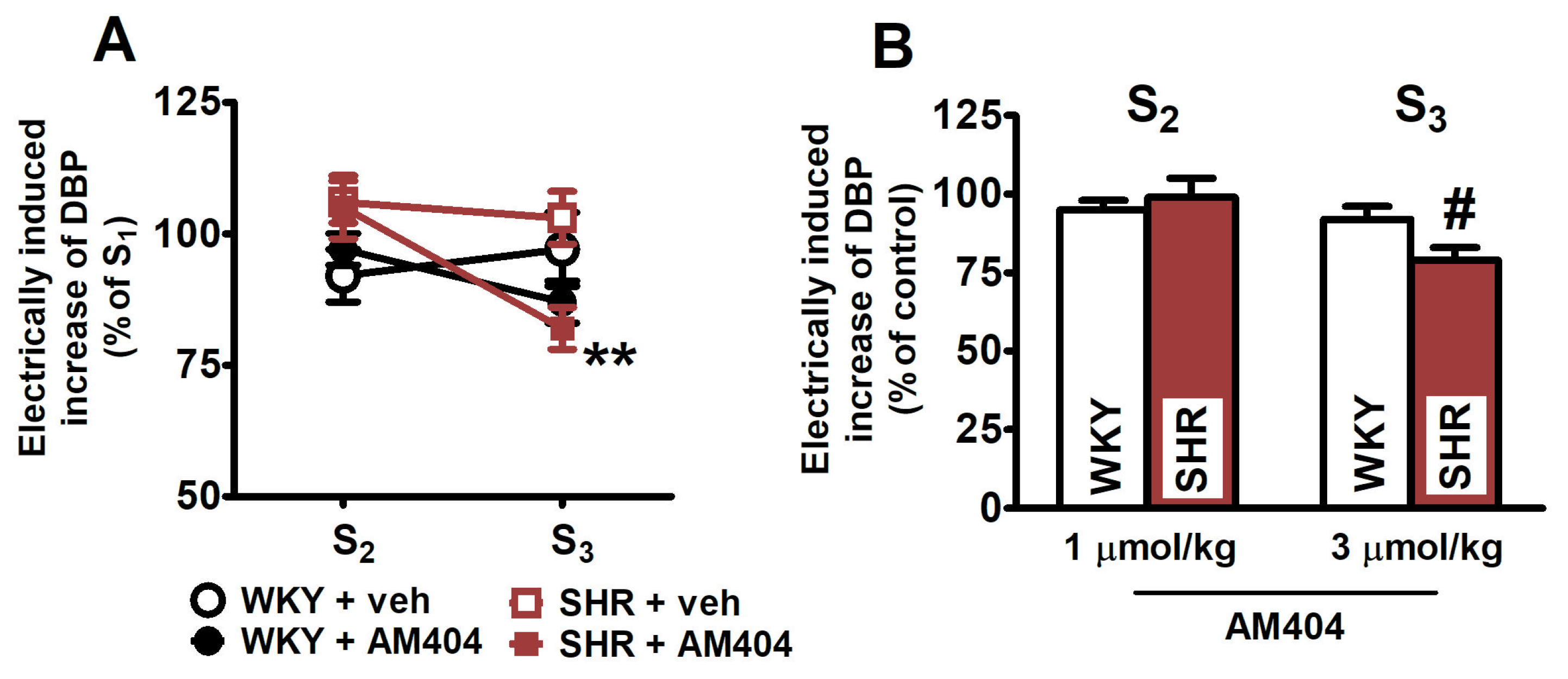
2.4. Effects of the Endocannabinoid Transporter Inhibitor AM404
3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter/Experimental Condition | Change in SHR vs. WKY | Ref. to SHR | Change in DOCA-Salt vs. UNX | Ref. to DOCA |
|---|---|---|---|---|
| Cardiovascular parameters in pithed rats (if not stated otherwise) | present paper | [35] | ||
| BP (conscious) | ↑ | ↑ | ||
| BP | ↑ | ↓ | ||
| HR (conscious) | ↑ | ↔ | ||
| HR | ↑ | ↓ | ||
| Electrically induced increase in DBP (S1) | ↔ | ↓ | ||
| Phenylephrine induced increase in DBP (S1) | ↔ | ↓ | ||
| Function of presynaptic inhibitory CB1 receptors (inhibitory effect of cannabinoid agonist CP55940) | ↔ | ↑ | ||
| Influence of FAAH and/or MAGL or endocannabinoid reuptake inhibitors on BP in conscious rats | ||||
| URB597 or AM3506 (acute) | ↓ | [31,32] | ND | |
| URB597 (chronic) | ↔ | [33] | ↔ a/↓ b | |
| JZL195 (acute) | ↓ | [34] | ND | [41] |
| JZL195 (chronic) | ↓ | [34] | ND | |
| AM404 or OMDM-2 (acute) | ↓ | [31] | ND | |
| AEA level | ||||
| plasma | ↓ c ↑ d | c [42], d [29] | ↑ | [42] |
| aorta/mesenteric G3 | ↑ | [30] | ↔/ND | [30] |
| heart | ↓ | [42] | ↑ | [42] |
| 2-AG level | ||||
| plasma | ↓ | [42] | ↑ | [42] |
| aorta/mesenteric G3 | ↑ | [30] | ↔/ND | [30] |
| heart | ↓ | [42] | ↑ | [42] |
| CB1 receptor density | ||||
| aorta/mesenteric G3 | ↑ | [30] | ↑ | [30] |
4. Materials and Methods
4.1. Animals
4.2. Conscious Rats
4.3. Pithed Rats
4.4. Experimental Protocol
4.5. Statistical Analysis
4.6. Materials
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, J.; Chen, X. Advances in pathogenesis and treatment of essential hypertension. Front. Cardiovasc. Med. 2022, 9, 1003852. [Google Scholar] [CrossRef] [PubMed]
- Dybiec, J.; Krzemińska, J.; Radzioch, E.; Szlagor, M.; Wronka, M.; Młynarska, E.; Rysz, J.; Franczyk, B. Advances in the pathogenesis and treatment of resistant hypertension. Int. J. Mol. Sci. 2023, 24, 12911. [Google Scholar] [CrossRef] [PubMed]
- DeLalio, L.J.; Sved, A.F.; Stocker, S.D. Sympathetic nervous system contributions to hypertension: Updates and therapeutic relevance. Can. J. Cardiol. 2020, 36, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Seravalle, G.; Grassi, G. Sympathetic nervous system and hypertension: New evidences. Auton. Neurosci. 2022, 238, 102954. [Google Scholar] [CrossRef] [PubMed]
- Matanes, F.; Khan, M.B.; Siddiqui, M.; Dudenbostel, T.; Calhoun, D.; Oparil, S. An update on refractory hypertension. Curr. Hypertens. Rep. 2022, 24, 225–234. [Google Scholar] [CrossRef]
- Kubista, H.; Boehm, S. Molecular mechanisms underlying the modulation of exocytotic noradrenaline release via presynaptic receptors. Pharmacol. Ther. 2006, 112, 213–242. [Google Scholar] [CrossRef]
- Molderings, G.J.; Likungu, J.; Göthert, M. Modulation of noradrenaline release from the sympathetic nerves of human right atrial appendages by presynaptic EP3- and DP-receptors. Naunyn Schmiedebergs Arch. Pharmacol. 1998, 358, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, A.; Esler, M.D.; Socratous, F.; Kaye, D.M. Evidence for functional presynaptic alpha-2 adrenoceptors and their down-regulation in human heart failure. J. Am. Coll. Cardiol. 2001, 37, 1246–1251. [Google Scholar] [CrossRef]
- Parker, D.A.; Marino, V. GABA heteroreceptors modulate noradrenaline release in human dental pulp. J. Dent. Res. 2013, 92, 1017–1021. [Google Scholar] [CrossRef]
- Misu, Y.; Kubo, T. Presynaptic β-adrenoceptors. Med. Res. Rev. 1986, 6, 197–225. [Google Scholar] [CrossRef]
- de Champlain, J. Pre- and postsynaptic adrenergic dysfunctions in hypertension. J. Hypertens. Suppl. 1990, 8, S77–S85. [Google Scholar]
- Foucart, S.; Patrick, S.K.; Oster, L.; de Champlain, J. Effects of chronic treatment with losartan and enalaprilat on [3H]-norepinephrine release from isolated atria of Wistar-Kyoto and spontaneously hypertensive rats. Am. J. Hypertens. 1996, 9, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Stegbauer, J.; Oberhauser, V.; Vonend, O.; Rump, L.C. Angiotensin-(1–7) modulates vascular resistance and sympathetic neurotransmission in kidneys of spontaneously hypertensive rats. Cardiovasc. Res. 2004, 61, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Bott-Flügel, L.; Bernshausen, A.; Schneider, H.; Luppa, P.; Zimmermann, K.; Albrecht-Küpper, B.; Kast, R.; Laugwitz, K.L.; Ehmke, H.; Knorr, A.; et al. Selective attenuation of norepinephrine release and stress-induced heart rate increase by partial adenosine A1 agonism. PLoS ONE 2011, 6, e18048. [Google Scholar] [CrossRef]
- Rocha-Pereira, C.; Arribas, S.M.; Fresco, P.; González, M.C.; Gonçalves, J.; Diniz, C. Impaired inhibitory function of presynaptic A1-adenosine receptors in SHR mesenteric arteries. J. Pharmacol. Sci. 2013, 122, 59–70. [Google Scholar] [CrossRef]
- Tabuchi, Y.; Nakamaru, M.; Rakugi, H.; Nagano, M.; Higashimori, K.; Mikami, H.; Ogihara, T. Effects of endothelin on neuroeffector junction in mesenteric arteries of hypertensive rats. Hypertension 1990, 15, 739–743. [Google Scholar] [CrossRef]
- Tsuda, K.; Tsuda, S.; Goldstein, M.; Nishio, I.; Masuyama, Y. Modulation of noradrenergic transmission by neuropeptide Y and presynaptic α2-adrenergic receptors in the hypothalamus of spontaneously hypertensive rats. Jpn. Heart J. 1992, 33, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Kimura, M.; Kamata, K. Mechanism underlying γ-aminobutyric acid-induced antihypertensive effect in spontaneously hypertensive rats. Eur. J. Pharmacol. 2002, 438, 107–113. [Google Scholar] [CrossRef]
- Lerman, L.O.; Kurtz, T.W.; Touyz, R.M.; Ellison, D.H.; Chade, A.R.; Crowley, S.D.; Mattson, D.L.; Mullins, J.J.; Osborn, J.; Eirin, A.; et al. Animal models of hypertension: A scientific statement from the American Heart Association. Hypertension 2019, 73, e87–e120. [Google Scholar] [CrossRef]
- Salman, I.M. Key challenges in exploring the rat as a preclinical neurostimulation model for aortic baroreflex modulation in hypertension. Hypertens. Res. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Malinowska, B.; Godlewski, G.; Bucher, B.; Schlicker, E. Cannabinoid CB1 receptor-mediated inhibition of the neurogenic vasopressor response in the pithed rat. Naunyn Schmiedebergs Arch. Pharmacol. 1997, 356, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, B.; Piszcz, J.; Koneczny, B.; Hryniewicz, A.; Schlicker, E. Modulation of the cardiac autonomic transmission of pithed rats by presynaptic opioid OP4 and cannabinoid CB1 receptors. Naunyn Schmiedebergs Arch. Pharmacol. 2001, 364, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Niederhoffer, N.; Schmid, K.; Szabo, B. The peripheral sympathetic nervous system is the major target of cannabinoids in eliciting cardiovascular depression. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 367, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Niederhoffer, N.; Szabo, B. Effect of the cannabinoid receptor agonist WIN55212-2 on sympathetic cardiovascular regulation. Br. J. Pharmacol. 1999, 126, 457–466. [Google Scholar] [CrossRef]
- Szabo, B.; Nordheim, U.; Niederhoffer, N. Effects of cannabinoids on sympathetic and parasympathetic neuroeffector transmission in the rabbit heart. J. Pharmacol. Exp. Ther. 2001, 297, 819–826. [Google Scholar]
- Ishac, E.J.; Jiang, L.; Lake, K.D.; Varga, K.; Abood, M.E.; Kunos, G. Inhibition of exocytotic noradrenaline release by presynaptic cannabinoid CB1 receptors on peripheral sympathetic nerves. Br. J. Pharmacol. 1996, 118, 2023–2028. [Google Scholar] [CrossRef]
- Molderings, G.J.; Likungu, J.; Göthert, M. Presynaptic cannabinoid and imidazoline receptors in the human heart and their potential relationship. Naunyn Schmiedebergs Arch. Pharmacol. 1999, 360, 157–164. [Google Scholar] [CrossRef]
- Maccarrone, M.; Di Marzo, V.; Gertsch, J.; Grether, U.; Howlett, A.C.; Hua, T.; Makriyannis, A.; Piomelli, D.; Ueda, N.; van der Stelt, M. Goods and bads of the endocannabinoid system as a therapeutic target: Lessons learned after 30 years. Pharmacol. Rev. 2023, 75, 885–958. [Google Scholar] [CrossRef]
- Li, D.; Chen, B.M.; Peng, J.; Zhang, Y.S.; Li, X.H.; Yuan, Q.; Hu, C.P.; Deng, H.W.; Li, Y.J. Role of anandamide transporter in regulating calcitonin gene-related peptide production and blood pressure in hypertension. J. Hypertens. 2009, 27, 1224–1232. [Google Scholar] [CrossRef]
- Baranowska-Kuczko, M.; Kozłowska, H.; Kloza, M.; Kusaczuk, M.; Harasim-Symbor, E.; Biernacki, M.; Kasacka, I.; Malinowska, B. Vasoprotective endothelial effects of chronic cannabidiol treatment and its influence on the endocannabinoid system in rats with primary and secondary hypertension. Pharmaceuticals 2021, 14, 1120. [Google Scholar] [CrossRef]
- Bátkai, S.; Pacher, P.; Osei-Hyiaman, D.; Radaeva, S.; Liu, J.; Harvey-White, J.; Offertáler, L.; Mackie, K.; Rudd, M.A.; Bukoski, R.D.; et al. Endocannabinoids acting at cannabinoid-1 receptors regulate cardiovascular function in hypertension. Circulation 2004, 110, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Godlewski, G.; Alapafuja, S.O.; Bátkai, S.; Nikas, S.P.; Cinar, R.; Offertáler, L.; Osei-Hyiaman, D.; Liu, J.; Mukhopadhyay, B.; Harvey-White, J.; et al. Inhibitor of fatty acid amide hydrolase normalizes cardiovascular function in hypertension without adverse metabolic effects. Chem. Biol. 2010, 17, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Pędzińska-Betiuk, A.; Weresa, J.; Toczek, M.; Baranowska-Kuczko, M.; Kasacka, I.; Harasim-Symbor, E.; Malinowska, B. Chronic inhibition of fatty acid amide hydrolase by URB597 produces differential effects on cardiac performance in normotensive and hypertensive rats. Br. J. Pharmacol. 2017, 174, 2114–2129. [Google Scholar] [CrossRef] [PubMed]
- Toczek, M.; Ryszkiewicz, P.; Remiszewski, P.; Schlicker, E.; Krzyżewska, A.; Kozłowska, H.; Malinowska, B. Weak hypotensive effect of chronic administration of the dual FAAH/MAGL inhibitor JZL195 in spontaneously hypertensive rats as revealed by area under the curve analysis. Int. J. Mol. Sci. 2023, 24, 10942. [Google Scholar] [CrossRef]
- Toczek, M.; Schlicker, E.; Grzęda, E.; Malinowska, B. Enhanced function of inhibitory presynaptic cannabinoid CB1 receptors on sympathetic nerves of DOCA-salt hypertensive rats. Life Sci. 2015, 138, 78–85. [Google Scholar] [CrossRef]
- Li, G.Q.; Wang, Z.; Zhao, T.; Dai, S.X.; Liu, J.M.; Jia, B.Z.; Zhang, Y.; Li, Q. Role of cannabinoid receptor type 1 in rostral ventrolateral medulla in high-fat diet-induced hypertension in rats. J. Hypertens. 2018, 36, 801–808. [Google Scholar] [CrossRef]
- Kurz, C.; Baranowska, U.; Łupiński, S.; Göthert, M.; Malinowska, B.; Schlicker, E. Urethane, but not pentobarbitone, attenuates presynaptic receptor function in rats: A contribution to the choice of anaesthetic. Br. J. Pharmacol. 2009, 157, 1474–1482. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB1 and CB2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef]
- Pertwee, R.G. Inverse agonism and neutral antagonism at cannabinoid CB1 receptors. Life Sci. 2005, 76, 1307–1324. [Google Scholar] [CrossRef]
- van Diepen, H.; Schlicker, E.; Michel, M.C. Prejunctional and peripheral effects of the cannabinoid CB1 receptor inverse agonist rimonabant (SR 141716). Naunyn Schmiedebergs Arch. Pharmacol. 2008, 378, 345–369. [Google Scholar] [CrossRef]
- Toczek, M.; Baranowska-Kuczko, M.; Grzęda, E.; Pędzińska-Betiuk, A.; Weresa, J.; Malinowska, B. Age-specific influences of chronic administration of the fatty acid amide hydrolase inhibitor URB597 on cardiovascular parameters and organ hypertrophy in DOCA-salt hypertensive rats. Pharmacol. Rep. 2016, 68, 363–369. [Google Scholar] [CrossRef]
- Biernacki, M.; Malinowska, B.; Timoszuk, M.; Toczek, M.; Jastrząb, A.; Remiszewski, P.; Skrzydlewska, E. Hypertension and chronic inhibition of endocannabinoid degradation modify the endocannabinoid system and redox balance in rat heart and plasma. Prostaglandins Other Lipid Mediat. 2018, 138, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Reichenbacher, D.; Reimann, W.; Starke, K. α-Adrenoceptor-mediated inhibition of noradrenaline release in rabbit brain cortex slices. Receptor properties and role of the biophase concentration of noradrenaline. Naunyn Schmiedebergs Arch. Pharmacol. 1982, 319, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Iring, A.; Ruisanchez, E.; Leszl-Ishiguro, M.; Horváth, B.; Benkő, R.; Lacza, Z.; Járai, Z.; Sándor, P.; Di Marzo, V.; Pacher, P.; et al. Role of endocannabinoids and cannabinoid-1 receptors in cerebrocortical blood flow regulation. PLoS ONE 2013, 8, e53390. [Google Scholar] [CrossRef] [PubMed]
- Hama, A.T.; Germano, P.; Varghese, M.S.; Cravatt, B.F.; Milne, G.T.; Pearson, J.P.; Sagen, J. Fatty acid amide hydrolase (FAAH) inhibitors exert pharmacological effects, but lack antinociceptive efficacy in rats with neuropathic spinal cord injury pain. PLoS ONE 2014, 9, e96396. [Google Scholar] [CrossRef]
- Niphakis, M.J.; Cognetta, A.B., III; Chang, J.W.; Buczynski, M.W.; Parsons, L.H.; Byrne, F.; Burston, J.J.; Chapman, V.; Cravatt, B.F. Evaluation of NHS carbamates as a potent and selective class of endocannabinoid hydrolase inhibitors. ACS Chem. Neurosci. 2013, 4, 1322–1332. [Google Scholar] [CrossRef]
- Long, J.Z.; Nomura, D.K.; Vann, R.E.; Walentiny, D.M.; Booker, L.; Jin, X.; Burston, J.J.; Sim-Selley, L.J.; Lichtman, A.H.; Wiley, J.L.; et al. Dual blockade of FAAH and MAGL identifies behavioral processes regulated by endocannabinoid crosstalk in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 20270–20275. [Google Scholar] [CrossRef]
- Godlewski, G.; Malinowska, B.; Schlicker, E.; Bucher, B. Identification of histamine H3 receptors in the tail artery from normotensive and spontaneously hypertensive rats. J. Cardiovasc. Pharmacol. 1997, 29, 801–807. [Google Scholar] [CrossRef]



| WKY | SHR | |||||
|---|---|---|---|---|---|---|
| Group | n | DBP (mmHg) | HR (beats/min) | n | DBP (mmHg) | HR (beats/min) |
| 36 | 43 ± 1 | 283 ± 4 | 37 | 49 ± 1 ### | 294 ± 3 # | |
| +AM251 | 13 | 39 ± 1 | 275 ± 6 | 14 | 52 ± 1 ### | 291 ± 6 |
| +vehicle for URB597 | 7 | 49 ± 2 | 275 ± 4 | 5 | 50 ± 1 | 282 ± 10 |
| +URB597 | 12 | 47 ± 1 | 270 ± 6 | 9 | 51 ± 1 ## | 299 ± 10 # |
| +vehicle for MJN110 | 5 | 41 ± 2 | 292 ± 4 | 3 | 49 ± 1 # | 296 ± 14 |
| +MJN110 | 10 | 43 ± 1 | 279 ± 6 | 9 | 49 ± 2 ### | 303 ± 8 # |
| +vehicle for JZL195 | 6 | 45 ± 4 | 256 ± 9 & | 5 | 52 ± 2 | 317 ± 11 ## |
| +JZL195 | 17 | 40 ± 2 | 266 ± 5 & | 18 | 50 ± 1 ### | 296 ± 6 ### |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toczek, M.; Schlicker, E.; Remiszewski, P.; Malinowska, B. Function of Presynaptic Inhibitory Cannabinoid CB1 Receptors in Spontaneously Hypertensive Rats and Its Modification by Enhanced Endocannabinoid Tone. Int. J. Mol. Sci. 2024, 25, 858. https://doi.org/10.3390/ijms25020858
Toczek M, Schlicker E, Remiszewski P, Malinowska B. Function of Presynaptic Inhibitory Cannabinoid CB1 Receptors in Spontaneously Hypertensive Rats and Its Modification by Enhanced Endocannabinoid Tone. International Journal of Molecular Sciences. 2024; 25(2):858. https://doi.org/10.3390/ijms25020858
Chicago/Turabian StyleToczek, Marek, Eberhard Schlicker, Patryk Remiszewski, and Barbara Malinowska. 2024. "Function of Presynaptic Inhibitory Cannabinoid CB1 Receptors in Spontaneously Hypertensive Rats and Its Modification by Enhanced Endocannabinoid Tone" International Journal of Molecular Sciences 25, no. 2: 858. https://doi.org/10.3390/ijms25020858
APA StyleToczek, M., Schlicker, E., Remiszewski, P., & Malinowska, B. (2024). Function of Presynaptic Inhibitory Cannabinoid CB1 Receptors in Spontaneously Hypertensive Rats and Its Modification by Enhanced Endocannabinoid Tone. International Journal of Molecular Sciences, 25(2), 858. https://doi.org/10.3390/ijms25020858