
QTL Mapping-Based Identification of Visceral White-Nodules Disease Resistance Genes in Larimichthys polyactis

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Genotyping and Genetic Marker Development
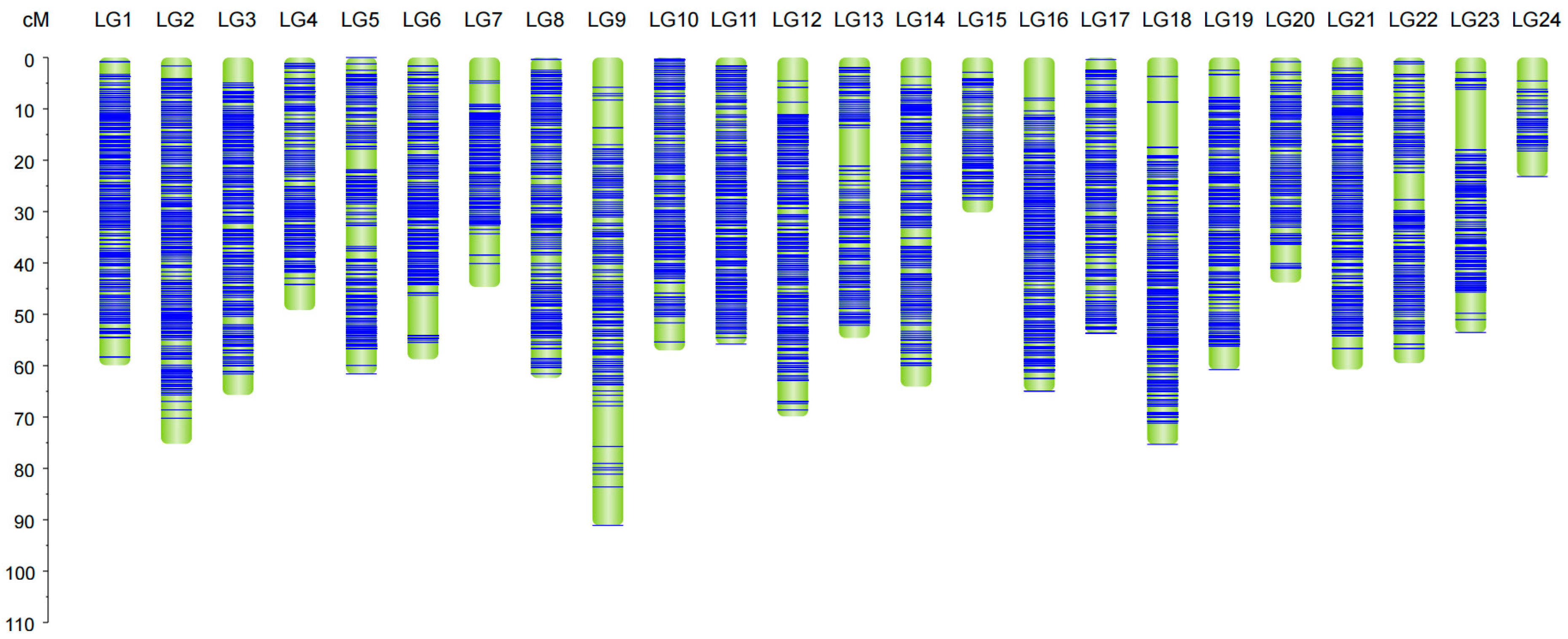
2.2. Construction of Linkage Maps
2.3. QTL Mapping of Disease-Related Traits
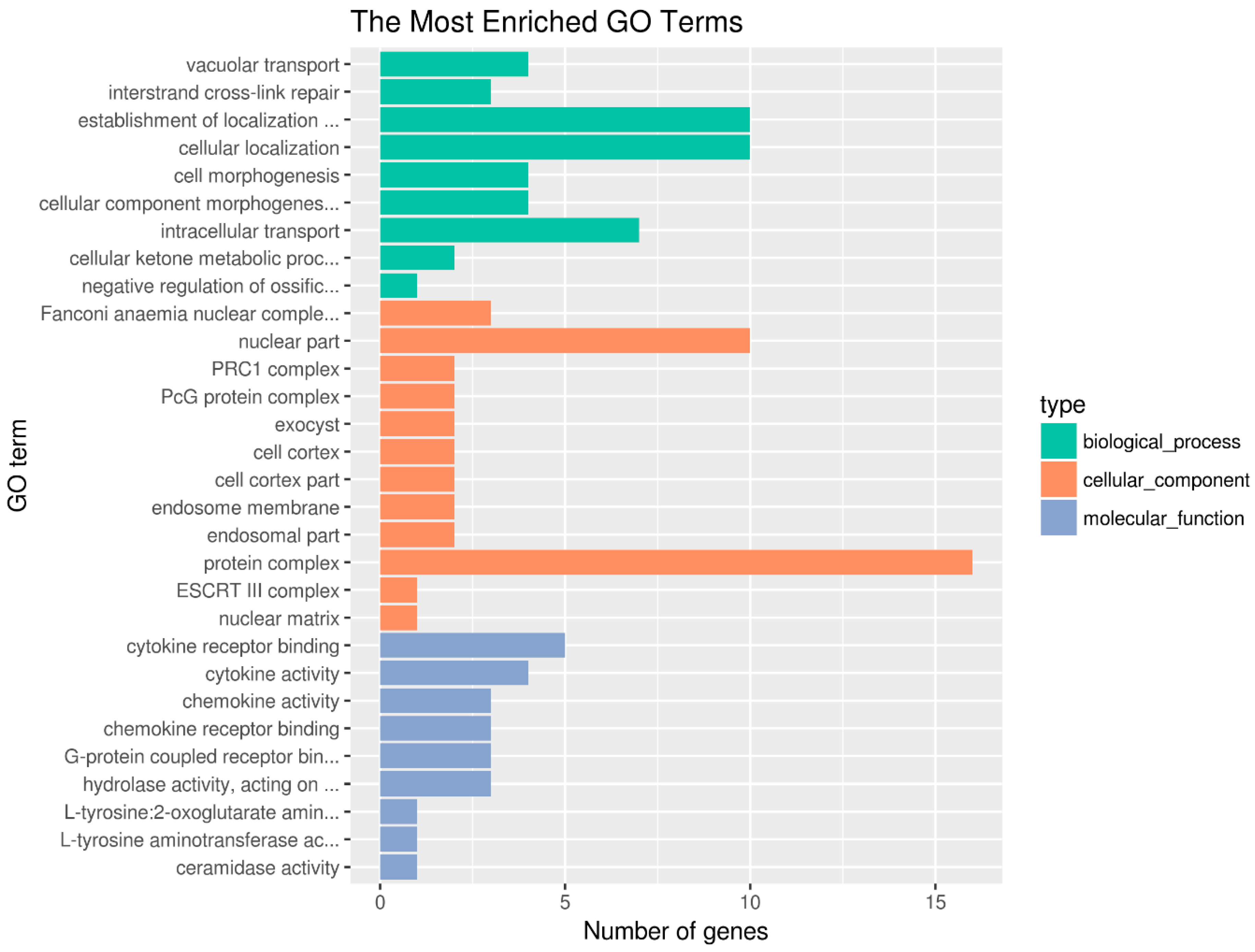
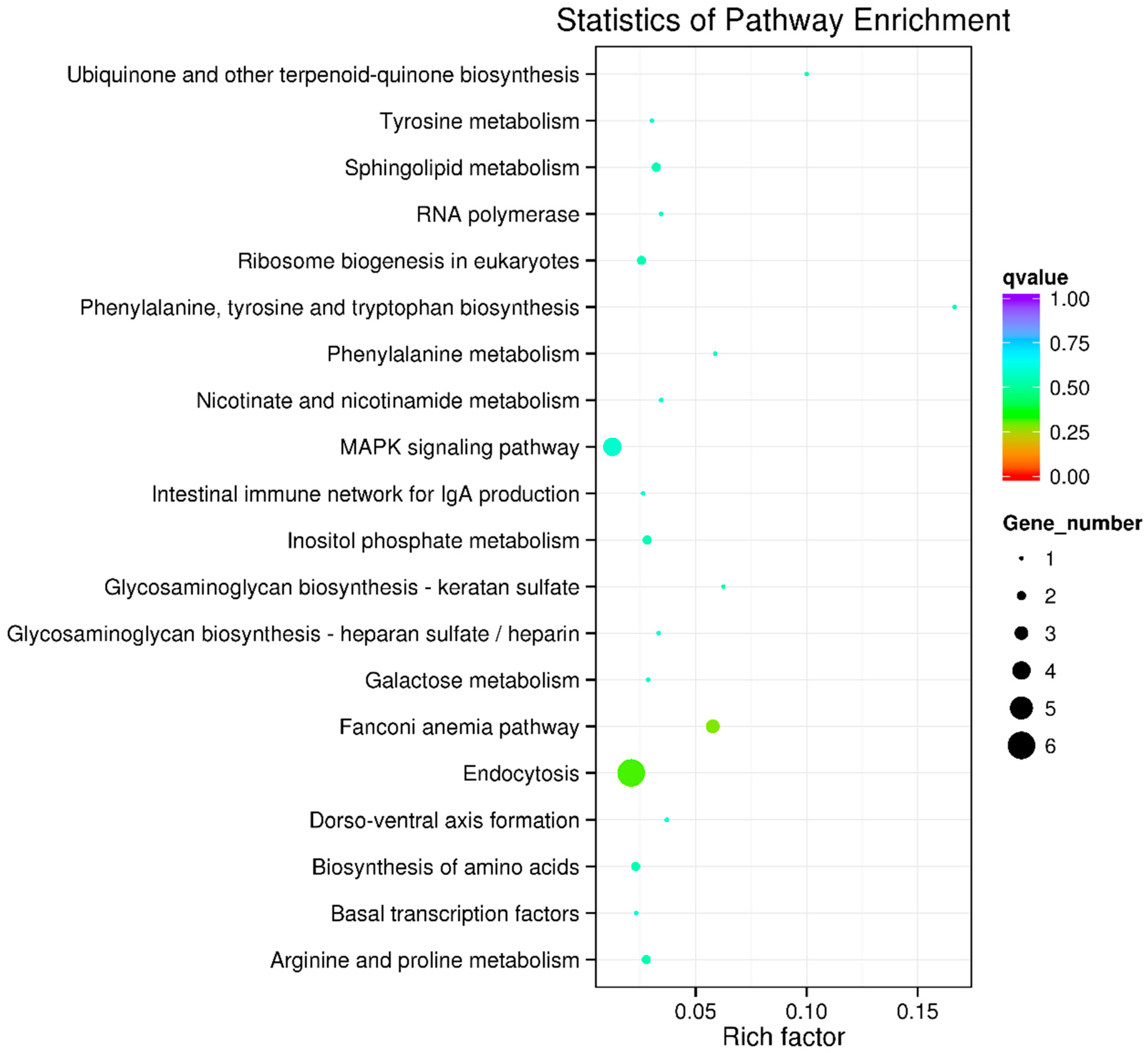
2.4. Functional Enrichment Analysis of Candidate Genes
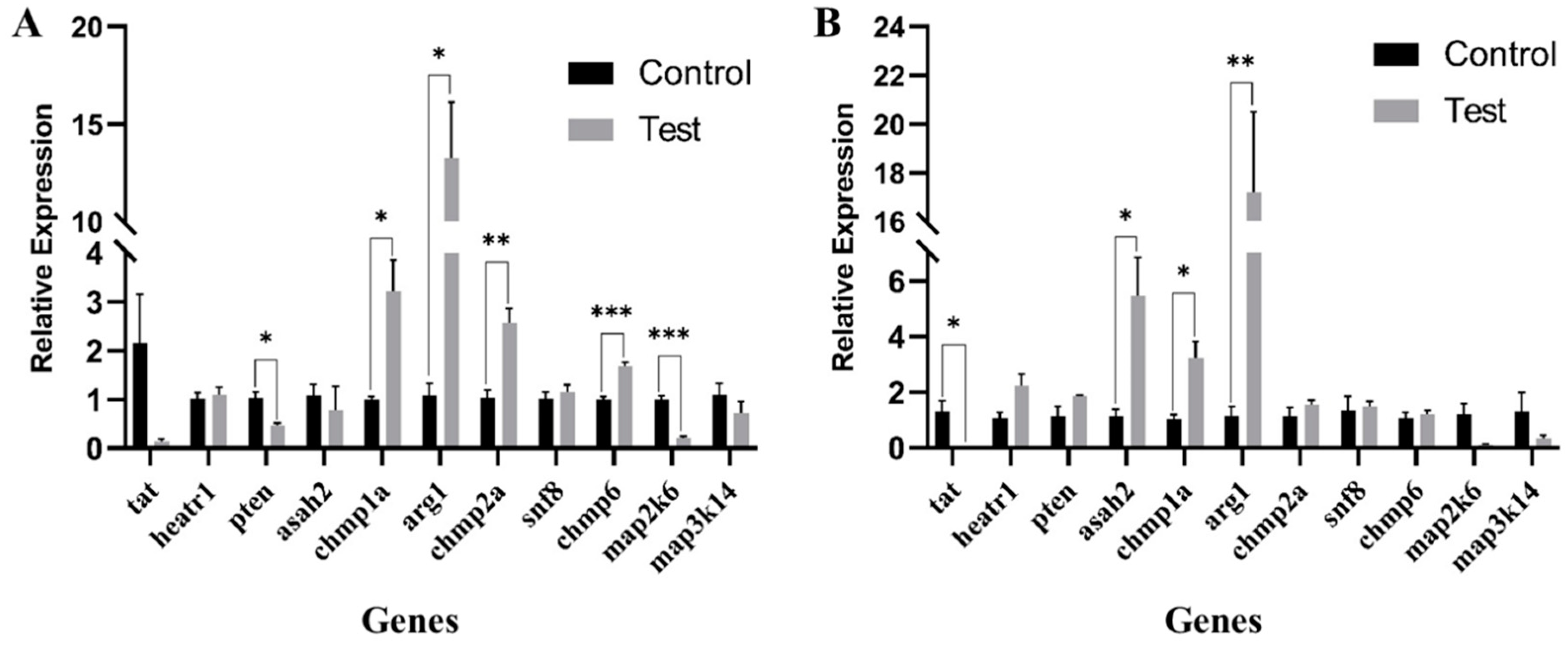
2.5. Identification of Disease Resistance Gene Candidates
3. Discussion
4. Materials and Methods
4.1. Challenge Experiment and Sample Collection
4.2. Sequencing, Genotyping, and SNP Filtering
4.3. Genetic Map Construction and QTL Mapping
4.4. Gene Annotation and Enrichment Analysis
4.5. Analysis of Candidate Gene Expression Levels
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.; Wang, W.; Zhou, W.; Hu, F.; Wu, M. Shifting Feeding Habits During Settlement Among Small Yellow Croakers (Larimichthys polyactis). Front. Mar. Sci. 2022, 8, 786724. [Google Scholar] [CrossRef]
- Wu, T.; Wu, C.; Fang, Z.; Ma, X.; Chen, S.; Hu, Y. Effect of chitosan microcapsules loaded with nisin on the preservation of small yellow croaker. Food Control 2017, 79, 317–324. [Google Scholar] [CrossRef]
- Chen, L.; Zeng, W.; Rong, Y.; Lou, B. Nutritional composition and textural quality of wild-caught and cage-cultured small yellow croaker (Larimichthys polyactis). J. Food Compos. Anal. 2021, 107, 104370. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, F.; Wang, Z.; You, Q.; Lou, B.; Xu, D.; Chen, R.; Zhan, W.; Liu, F. Mitochondrial DNA variation and population genetic structure in the small yellow croaker at the coast of Yellow Sea and East China Sea. Biochem. Syst. Ecol. 2017, 71, 236–243. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, T.; He, Y.; Zhan, W.; Xie, Q.; Lou, B. Integration of transcriptome and proteome analyses reveals the regulation mechanisms of Larimichthys polyactis liver exposed to heat stress. Fish Shellfish Immunol. 2023, 135, 108704. [Google Scholar] [CrossRef]
- Ina-Salwany, M.Y.; Al-Saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef]
- Duan, X.; Li, J.; Shi, H.; Tao, Z.; Wei, X.; Ye, Y.; Guo, B. Establishment of Nested PCR for the Detection of Pseudomonas plecoglossicida and Epidemiological Survey of Larimichthys crocea in the Southeast Coastal Region. Animals 2024, 14, 1427. [Google Scholar] [CrossRef]
- Jiao, J.; Zhao, L.; Huang, L.; Qin, Y.; Su, Y.; Zheng, W.; Zhang, J.; Yan, Q. The contributions of fliG gene to the pathogenicity of Pseudomonas plecoglossicida and pathogen-host interactions with Epinephelus coioides. Fish Shellfish Immunol. 2021, 119, 238–248. [Google Scholar] [CrossRef]
- Akaylı, T.; Çanak, Ö.; Başaran, B. A new Pseudomonas species observed in cultured young rainbow trout (Oncorhynchus mykiss Walbaum, 1792): Pseudomonas plecoglossicida. BIBAD 2011, 4, 107–111. [Google Scholar]
- Yan, L.; Jin, D.; Yang, S.; Li, X.; Li, H.; Hu, S.; Sun, Y.; Yi, G.; Wang, P.; Rang, J.; et al. Pathogenicity of fish pathogen Pseudomonas plecoglossicida and preparation of its inactivated vaccine. Microb. Pathog. 2022, 166, 105488. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, Y.; Sun, F.; Zhang, Y.; Liu, Q.; Wang, Q.; Yang, D.; Zhang, Y. The c-di-GMP signalling component YfiR regulates multiple bacterial phenotypes and virulence in Pseudomonas plecoglossicida. J. Appl. Microbiol. 2023, 134, 157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, S.; An, S.; Chen, L.; Wang, G. Visceral granulomas in farmed large yellow croaker, Larimichthys crocea (Richardson), caused by a bacterial pathogen, Pseudomonas plecoglossicida. J. Fish. Dis. 2014, 37, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.H.; Chiu, Y.T.; Lee, H.T.; Gong, H.Y.; Huang, C.W. Development of EST-Molecular Markers from RNA Sequencing for Genetic Management and Identification of Growth Traits in Potato Grouper (Epinephelus tukula). Biology 2021, 10, 36. [Google Scholar] [CrossRef]
- Uchino, T.; Tabata, J.; Yoshida, K.; Suzuki, T.; Noda, T.; Fujinami, Y.; Ozaki, A. Novel Benedenia disease resistance QTLs in five F1 families of yellowtail (Seriola quinqueradiata). Aquaculture 2020, 529, 735622. [Google Scholar] [CrossRef]
- Marana, M.H.; Karami, A.M.; Ødegård, J.; Zuo, S.; Jaafar, R.M.; Mathiessen, H.; von Gersdorff Jørgensen, L.; Kania, P.W.; Dalsgaard, I.; Nielsen, T.; et al. Whole-genome association study searching for QTL for Aeromonas salmonicida resistance in rainbow trout. Sci. Rep. 2021, 11, 17857. [Google Scholar] [CrossRef] [PubMed]
- Grover, A.; Sharma, P.C. Development and use of molecular markers: Past and present. Crit. Rev. Biotechnol. 2016, 36, 290–302. [Google Scholar] [CrossRef]
- Robledo, D.; Palaiokostas, C.; Bargelloni, L.; Martínez, P.; Houston, R. Applications of genotyping by sequencing in aquaculture breeding and genetics. Rev. Aquac. 2018, 10, 670–682. [Google Scholar] [CrossRef]
- Tsai, H.Y.; Robledo, D.; Lowe, N.R.; Bekaert, M.; Taggart, J.B.; Bron, J.E.; Houston, R.D. Construction and annotation of a high density SNP linkage map of the Atlantic Salmon (Salmo salar) genome. G3 2016, 6, 2173–2179. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Qin, Z.; Waldbieser, G.; Wang, R.; Sun, L.; Bao, L.; Danzmann, R.G.; Dunham, R.; Liu, Z. Construction of a high-density, high-resolution genetic map and its integration with BAC-based physical map in channel catfish. DNA Res. 2015, 22, 39–52. [Google Scholar] [CrossRef]
- Shao, C.; Niu, Y.; Rastas, P.; Liu, Y.; Xie, Z.; Li, H.; Wang, L.; Jiang, Y.; Tai, S.; Tian, Y.; et al. Genome-wide SNP identification for the construction of a high-resolution genetic map of Japanese flounder (Paralichthys olivaceus): Applications to QTL mapping of Vibrio anguillarum disease resistance and comparative genomic analysis. DNA Res. 2015, 22, 161–170. [Google Scholar] [CrossRef]
- Wang, L.; Wan, Z.Y.; Bai, B.; Huang, S.Q.; Chua, E.; Lee, M.; Pang, H.Y.; Wen, Y.F.; Liu, P.; Liu, F.; et al. Construction of a high-density linkage map and fine mapping of QTL for growth in Asian seabass. Sci. Rep. 2015, 5, 16358. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Zhang, G. A high-density SNP genetic linkage map and QTL analysis of growth-related traits in a hybrid family of oysters (Crassostrea gigas × Crassostrea angulata) using genotyping-by-sequencing. G3 2016, 6, 1417–1426. [Google Scholar] [CrossRef]
- Xiao, S.; Wang, P.; Zhang, Y.; Fang, L.; Liu, Y.; Li, J.T.; Wang, Z.Y. Gene map of large yellow croaker (Larimichthys crocea) provides insights into teleost genome evolution and conserved regions associated with growth. Sci. Rep. 2015, 5, 18661. [Google Scholar] [CrossRef] [PubMed]
- Weller, C.A.; Andreev, I.; Chambers, M.J.; Park, M.; Program, N.C.S.; Bloom, J.S.; Sadhu, M.J. Highly complete long-read genomes reveal pangenomic variation underlying yeast phenotypic diversity. Genome Res. 2023, 33, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Sawayama, E.; Tanizawa, S.; Kitamura, S.I.; Nakayama, K.; Ohta, K.; Ozaki, A.; Takagi, M. Identification of quantitative trait loci for resistance to RSIVD in red sea bream (Pagrus major). Mar. Biotechnol. 2017, 19, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Divilov, K.; Merz, N.; Schoolfield, B.; Green, T.; Langdon, C. Marker-assisted selection in a Pacific oyster population for an antiviral QTL conferring increased survival to OsHV-1 mortality events in Tomales Bay. Aquaculture 2023, 567, 739291. [Google Scholar] [CrossRef]
- Han, M.; Liu, Y.; Jin, C.; Wang, X.; Song, W.; He, Y.; Zhang, Q. Potential loci and candidate genes associated with Vibrio anguillarum resistance in black rockfish (Sebastes schlegelii) revealed by BSA-seq analysis. Aquaculture 2023, 566, 739201. [Google Scholar] [CrossRef]
- Rodríguez-Ramilo, S.T.; De La Herrán, R.; Ruiz-Rejón, C.; Hermida, M.; Fernández, C.; Pereiro, P.; Figueras, A.; Bouza, C.; Toro, M.A.; Martínez, P.; et al. Identification of quantitative trait loci associated with resistance to viral haemorrhagic septicaemia (VHS) in turbot (Scophthalmus maximus): A comparison between bacterium, parasite and virus diseases. Mar. Biotechnol. 2014, 16, 265–276. [Google Scholar] [CrossRef]
- Beck, E.A.; Currey, M.C.; Small, C.M.; Cresko, W.A. QTL mapping of intestinal neutrophil variation in threespine stickleback reveals possible gene targets connecting intestinal inflammation and systemic health. G3 2020, 10, 613–622. [Google Scholar] [CrossRef]
- Wang, L.; Bai, B.; Huang, S.; Liu, P.; Wan, Z.Y.; Ye, B.; Wu, J.; Yue, G.H. QTL mapping for resistance to Iridovirus in Asian Seabass using genotyping-by-sequencing. Mar. Biotechnol. 2017, 19, 517–527. [Google Scholar] [CrossRef]
- Lu, C.; Laghari, M.Y.; Laghari, M.Y.; Zheng, X.; Cao, D.; Zhang, X.; Kuang, Y.; Li, C.; Cheng, L.; Mahboob, S.; et al. Mapping quantitative trait loci and identifying candidate genes affecting feed conversion ratio based onto two linkage maps in common carp (Cyprinus carpio L). Aquaculture 2017, 468, 585–596. [Google Scholar] [CrossRef]
- Qiu, C.; Han, Z.; Li, W.; Ye, K.; Xie, Y.; Wang, Z. A high-density genetic linkage map and QTL mapping for growth and sex of yellow drum (Nibea albiflora). Sci. Rep. 2018, 8, 17271. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, A.; Mao, S.; Xu, X.; Li, J.; Shen, Y. Construction of high-density genetic linkage map and QTL mapping for growth performance in black carp (Mylopharyngodon piceus). Aquaculture 2021, 549, 737799. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, M.; Wang, S.; Xu, X.; Shen, Y.; Li, J. A high-density genetic linkage map and QTL mapping for growth related traits in grass carp (Ctenopharyngodon idella). Aquaculture 2022, 552, 738041. [Google Scholar] [CrossRef]
- Houston, R.D. Future directions in breeding for disease resistance in aquaculture species. Rev. Bras. Zootec. 2017, 46, 545–551. [Google Scholar]
- Liu, P.; Wang, L.; Wan, Z.Y.; Ye, B.Q.; Huang, S.; Wong, S.M.; Yue, G.H. Mapping QTL for Resistance Against Viral Nervous Necrosis Disease in Asian Seabass. Mar. Biotechnol. 2016, 18, 107–116. [Google Scholar] [CrossRef]
- Wang, X.; Xu, W.; Liu, Y.; Wang, L.; Sun, H.; Chen, S. Quantitative trait loci detection of Edwardsiella tarda resistance in Japanese flounder Paralichthys olivaceus using bulked segregant analysis. Chin. J. Oceanol. Limnol. 2016, 34, 1297–1308. [Google Scholar] [CrossRef]
- Bajgar, A.; Kucerova, K.; Jonatova, L.; Tomcala, A.; Schneedorferova, I.; Okrouhlik, J.; Dolezal, T. Extracellular adenosine mediates a systemic metabolic switch during immune response. PLoS Biol. 2015, 13, e1002135. [Google Scholar] [CrossRef] [PubMed]
- Jeria, E.; Oyanedel, D.; Rojas, R.; Farlora, R.; Lira, G.; Mercado, A.; Muñoz, K.; Destoumieux-Garzón, D.; Brokordt, K.; Schmitt, P. Resistance of Argopecten purpuratus scallop larvae to vibriosis is associated with the front-loading of immune genes and enhanced antimicrobial response. Front. Immunol. 2023, 14, 1150280. [Google Scholar] [CrossRef]
- Zheng, Z.; Wang, F.; Aweya, J.J.; Li, R.; Yao, D.; Zhong, M.; Li, S.; Zhang, Y. Comparative transcriptomic analysis of shrimp hemocytes in response to acute hepatopancreas necrosis disease (AHPND) causing Vibrio parahemolyticus infection. Fish Shellfish Immunol. 2018, 74, 10–18. [Google Scholar] [CrossRef]
- Varghese, T.; Dasgupta, S.; Anand, G.; Rejish Kumar, V.J.; Sahu, N.P.; Pal, A.K.; Puthiyottil, M. Dietary arginine attenuates hypoxia- induced HIF expression, metabolic responses and oxidative stress in Indian Major Carp, Cirrhinus mrigala. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2022, 259, 110714. [Google Scholar] [CrossRef]
- Chen, Z.; Ceballos-Francisco, D.; Guardiola, F.A.; Huang, D.; Esteban, M.Á. Skin wound healing in gilthead seabream (Sparus aurata L.) fed diets supplemented with arginine. Fish Shellfish Immunol. 2020, 104, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Arlauckas, S.P.; Garren, S.B.; Garris, C.S.; Kohler, R.H.; Oh, J.; Pittet, M.J.; Weissleder, R. Arg1 expression defines immunosuppressive subsets of tumor-associated macrophages. Theranostics 2018, 8, 5842–5854. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Wu, C.Y.; Lin, Y.W.; Park, S.W.; Wei, L.N. Synergistic activation of Arg1 gene by retinoic acid and IL-4 involves chromatin remodeling for transcription initiation and elongation coupling. Nucleic Acids Res. 2016, 44, 7568–7579. [Google Scholar] [CrossRef]
- Bronte, V.; Zanovello, P. Regulation of immune responses by L-arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641–654. [Google Scholar] [CrossRef]
- Rossetti, I.; Zambusi, L.; Finardi, A.; Bodini, A.; Provini, L.; Furlan, R.; Morara, S. Calcitonin gene-related peptide decreases IL-1beta, IL-6 as well as Ym1, Arg1, CD163 expression in a brain tissue context-dependent manner while ameliorating experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2018, 323, 94–104. [Google Scholar] [CrossRef]
- Schmidt, O.; Teis, D. The ESCRT machinery. Curr. Biol. 2012, 22, R116–R120. [Google Scholar] [CrossRef]
- Alonso, Y.; Adell, M.; Migliano, S.M.; Teis, D. ESCRT-III and Vps4: A dynamic multipurpose tool for membrane budding and scission. FEBS J. 2016, 283, 3288–3302. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H. ESCRTs are everywhere. EMBO J. 2015, 34, 2398–2407. [Google Scholar] [CrossRef]
- Scanlon, S.T. Fine-tuning pyroptosis with ESCRT-III. Science 2018, 362, 905.5–906. [Google Scholar]
- Wu, Y.; Wu, Y.; Xu, C.; Sun, W.; You, Z.; Wang, Y.; Zhang, S. CHMP1A suppresses the growth of renal cell carcinoma cells via regulation of the PI3K/mTOR/p53 signaling pathway. Genes Genom. 2022, 44, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Zhang, D.; Ni, N.; Tang, Z.; Gao, H.; Ju, Y.; Dai, X.; Wang, J.; Gu, P.; Ji, J. miR-17 regulates the proliferation and differentiation of retinal progenitor cells by targeting CHMP1A. Biochem. Biophys. Res. Commun. 2020, 523, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Shiiki, T.; Matsui, S.; Okamoto, N.; Koshimizu, E.; Tsuchida, N.; Uchiyama, Y.; Hamanaka, K.; Fujita, A.; Miyatake, S.; et al. A novel homozygous CHMP1A variant arising from segmental uniparental disomy causes pontocerebellar hypoplasia type 8. J. Hum. Genet. 2023, 68, 247–253. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; Xin, Y.; Liu, Y.; Sun, J.; Zhou, G.; Gao, H.; Xu, P.; Chen, Y.; Chen, G.; Zhang, L.; et al. Chmp1A acts as a tumor suppressor gene that inhibits proliferation of renal cell carcinoma. Cancer Lett. 2012, 319, 190–196. [Google Scholar] [CrossRef]
- Hattori, T.; Takahashi, Y.; Chen, L.; Tang, Z.; Wills, C.A.; Liang, X.; Wang, H.G. Targeting the ESCRT-III component CHMP2A for noncanonical Caspase-8 activation on autophagosomal membranes. Cell Death Differ. 2021, 28, 657–670. [Google Scholar] [CrossRef]
- Bernareggi, D.; Xie, Q.; Prager, B.C.; Yun, J.; Cruz, L.S.; Pham, T.V.; Kim, W.; Lee, X.; Coffey, M.; Zalfa, C.; et al. CHMP2A regulates tumor sensitivity to natural killer cell-mediated cytotoxicity. Nat. Commun. 2022, 13, 1899. [Google Scholar] [CrossRef]
- Fu, D.; Tian, L.; Peng, Z.; Deng, W.; Yuan, J.; Ma, D.; Shi, T.; Li, D.; Wang, Y. Overexpression of CHMP6 induces cellular oncosis and apoptosis in HeLa cells. Biosci. Biotechnol. Biochem. 2009, 73, 494–501. [Google Scholar] [CrossRef]
- Goliand, I.; Nachmias, D.; Gershony, O.; Elia, N. Inhibition of ESCRT-II-CHMP6 interactions impedes cytokinetic abscission and leads to cell death. Mol. Biol. Cell 2014, 25, 3740–3748. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Sharrocks, A.D.; Whitmarsh, A.J. Transcriptional regulation by the MAP kinase signaling cascades. Gene 2003, 320, 3–21. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, S.; Zheng, S.; Zhang, Z.; Xu, S. Chlorpyrifos Suppresses Neutrophil Extracellular Traps in Carp by Promoting Necroptosis and Inhibiting Respiratory Burst Caused by the PKC/MAPK Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 1763589. [Google Scholar] [CrossRef]
- Chen, J.; Shao, B.; Wang, J.; Shen, Z.; Liu, H.; Li, S. Chlorpyrifos caused necroptosis via MAPK/NF-κB/TNF-α pathway in common carp (Cyprinus carpio L.) gills. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 249, 109126. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Yang, T.; Dong, R.; Yan, Y.; Jiang, Q. Astaxanthin protects tilapia head kidney cells against polystyrene microplastics-induced inflammation through MAPK and NF-κB signaling pathways. Aquaculture 2023, 574, 739686. [Google Scholar] [CrossRef]
- Tao, S.; Li, X.; Wang, J.; Bai, Y.; Wang, J.; Yang, Y.; Zhao, Z. Examination of the relationship of carbonate alkalinity stress and ammonia metabolism disorder-mediated apoptosis in the Chinese mitten crab, Eriocheir sinensis: Potential involvement of the ROS/MAPK signaling pathway. Aquaculture 2024, 579, 740179. [Google Scholar] [CrossRef]
- Lotan, T.L.; Lyon, M.; Huo, D.; Taxy, J.B.; Brendler, C.; Foster, B.A.; Stadler, W.; Rinker-Schaeffer, C.W. Up-regulation of MKK4, MKK6 and MKK7 during prostate cancer progression: An important role for SAPK signalling in prostatic neoplasia. J. Pathol. 2007, 212, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Ou, J.; Kou, L.; Deng, J.; Luo, S. Circ_016719 plays a critical role in neuron cell apoptosis induced by I/R via targeting miR-29c/Map2k6. Mol. Cell. Probes 2020, 49, 101478. [Google Scholar] [CrossRef]
- Lin, S.; Liu, K.; Zhang, Y.; Jiang, M.; Lu, R.; Folts, C.J.; Gao, X.; Noble, M.D.; Zhao, T.; Zhou, Z.; et al. Pharmacological targeting of p38 MAP-Kinase 6 (MAP2K6) inhibits the growth of esophageal adenocarcinoma. Cell Signal. 2018, 51, 222–232. [Google Scholar] [CrossRef]
- Kumar, V.; Behera, R.; Lohite, K.; Karnik, S.; Kundu, G.C. p38 kinase is crucial for osteopontin-induced furin expression that supports cervical cancer progression. Cancer Res. 2010, 70, 10381–10391. [Google Scholar] [CrossRef]
- Lee, Y.R.; Chen, M.; Pandolfi, P.P. The functions and regulation of the PTEN tumour suppressor: New modes and prospects. Nat. Rev. Mol. Cell Biol. 2018, 19, 547–562. [Google Scholar] [CrossRef]
- Sun, C.; Tao, Y.; Gao, Y.; Xia, Y.; Liu, Y.; Wang, G.; Gu, Y. F-box protein 11 promotes the growth and metastasis of gastric cancer via PI3K/AKT pathway-mediated EMT. Biomed. Pharmacother. 2018, 98, 416–423. [Google Scholar] [CrossRef]
- Li, W.; Guan, X.; Sun, L. hosphatase and Tensin Homolog (PTEN) of Japanese Flounder-Its Regulation by miRNA and Role in Autophagy, Apoptosis and Pathogen Infection. Int. J. Mol. Sci. 2020, 21, 7725. [Google Scholar] [CrossRef]
- Howe, C.; Mitchell, J.; Kim, S.J.; Im, E.; Rhee, S.H. Pten gene deletion in intestinal epithelial cells enhances susceptibility to Salmonella Typhimurium infection in mice. J. Microbiol. 2019, 57, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Briercheck, E.L.; Trotta, R.; Chen, L.; Hartlage, A.S.; Cole, J.P.; Cole, T.D.; Mao, C.; Banerjee, P.P.; Hsu, H.T.; Mace, E.M.; et al. PTEN is a negative regulator of NK cell cytolytic function. J. Immunol. 2015, 194, 1832–1840. [Google Scholar] [CrossRef] [PubMed]
- Snider, A.J.; Wu, B.X.; Jenkins, R.W.; Sticca, J.A.; Kawamori, T.; Hannun, Y.A.; Obeid, L.M. Loss of neutral ceramidase increases inflammation in a mouse model of inflammatory bowel disease. Prostaglandins Other Lipid Mediat. 2012, 99, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Maltesen, H.R.; Troelsen, J.T.; Olsen, J. Identification of a functional hepatocyte nuclear factor 4 binding site in the neutral ceramidase promoter. J. Cell. Biochem. 2010, 111, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- García-Barros, M.; Coant, N.; Kawamori, T.; Wada, M.; Snider, A.J.; Truman, J.P.; Wu, B.X.; Furuya, H.; Clarke, C.J.; Bialkowska, A.B.; et al. Role of neutral ceramidase in colon cancer. FASEB J. 2016, 30, 4159–4171. [Google Scholar] [CrossRef]
- Zhu, H.; Klement, J.D.; Lu, C.; Redd, P.S.; Yang, D.; Smith, A.D.; Poschel, D.B.; Zou, J.; Liu, D.; Wang, P.G.; et al. Asah2 represses the p53-Hmox1 axis to protect myeloid-derived suppressor cells from ferroptosis. J. Immunol. 2021, 206, 1395–1404. [Google Scholar] [CrossRef]
- Duan, C.; Wang, Z.; Wu, J.; Tan, C.; Fang, F.; Qian, W.; Han, C.; Hou, X. Fut2 deficiency promotes intestinal stem cell aging by damaging mitochondrial functions via down-regulating α1,2-fucosylation of Asah2 and Npc1. Research 2024, 7, 0343. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, J.; Jia, Z.; Yang, Z.; Liu, S.; Wang, H.; Zhao, C.; Zhao, J.; Tang, Q.; Xiong, Y.; et al. TRIM21-mediated ubiquitylation of TAT suppresses liver metastasis in gallbladder cancer. Cancer Lett. 2024, 592, 216923. [Google Scholar] [CrossRef]
- Manzini, B.M.; da Silva Santos Duarte, A.; Sankaramanivel, S.; Ramos, A.L.; Latuf-Filho, P.; Escanhoela, C.; Kharmandayan, P.; Olalla Saad, S.T.; Boin, I.; Malheiros Luzo, Â.C. Useful properties of undifferentiated mesenchymal stromal cells and adipose tissue as the source in liver-regenerative therapy studied in an animal model of severe acute fulminant hepatitis. Cytotherapy 2015, 17, 1052–1065. [Google Scholar] [CrossRef]
- Nandi, S.S.; Zheng, H.; Sharma, N.M.; Shahshahan, H.R.; Patel, K.P.; Mishra, P.K. Lack of miR-133a decreases contractility of diabetic hearts: A role for novel cross talk between tyrosine aminotransferase and tyrosine hydroxylase. Diabetes 2016, 65, 3075–3090. [Google Scholar] [CrossRef]
- Adams, S.H. Emerging perspectives on essential amino acid metabolism in obesity and the insulin-resistant state. Adv. Nutr. 2011, 2, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, L.; Chen, J.; Li, C.; Sun, H.; Wang, J.; Xiao, H. Activation of tyrosine metabolism in CD13+ cancer stem cells drives relapse in Hepatocellular Carcinoma. Cancer Res. Treat. 2020, 52, 604–621. [Google Scholar] [CrossRef]
- Yamashita, D.; Bernstock, J.D.; Elsayed, G.; Sadahiro, H.; Mohyeldin, A.; Chagoya, G.; Ilyas, A.; Mooney, J.; Estevez-Ordonez, D.; Yamaguchi, S.; et al. Targeting glioma-initiating cells via the tyrosine metabolic pathway. J. Neurosurg. 2020, 134, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Lin, J.; Li, W.; Chen, H.; Zhang, J.; Zhong, M.; Xue, J.; Mo, C.; Chen, L.; Zhu, Y.; et al. Homogentisic acid metabolism inhibits papillary thyroid carcinoma proliferation through ROS and p21-induced cell cycle arrest. Life Sci. 2024, 347, 122682. [Google Scholar] [CrossRef]
- Liu, F.; Liu, Y.; Chu, T.; Lou, B.; Zhan, W.; Chen, R.Y. Interspecific hybridization and genetic characterization of Larimichthys polyactis (♀) and L. crocea (♂). Aquac. Int. 2019, 27, 663–674. [Google Scholar] [CrossRef]
- Liu, F.; Ye, T.; Zhang, T.L.; Zhu, J.J.; Liu, H.W.; Li, Q.; Liu, S.F.; Guo, D.D.; Zhan, W.; Lou, B. Development of a 100K SNP array derived from the high-quality genome of Larimichthys polyactis and its application in genomic selection for growth and disease resistance. Unpublished.
- Jung, Y.; Han, D. BWA-MEME: BWA-MEM emulated with a machine learning approach. Bioinformatics 2022, 38, 2404–2413. [Google Scholar] [CrossRef] [PubMed]
- Brouard, J.S.; Bissonnette, N. Variant calling from RNA-seq data using the GATK joint genotyping workflow. Methods Mol. Biol. 2022, 2493, 205–233. [Google Scholar]
- Rastas, P. Lep-MAP3: Robust linkage mapping even for low-coverage whole genome sequencing data. Bioinformatics 2017, 33, 3726–3732. [Google Scholar] [CrossRef] [PubMed]
- Vinod, K.K. Kosambi and the genetic mapping function. Resonance 2011, 16, 540–550. [Google Scholar] [CrossRef]
- Van Ooijn, J.W. Sofware for the Mapping of Quantitative Trait Loci in Experimental Populations of Diploid Species; MapQTL® 6.0; Kyazma BV: Wageningen, The Netherlands, 2009. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker Type | Female Genotype | Male Genotype | SNP Number | After Segregation | Bin Marker Number | Percentage |
|---|---|---|---|---|---|---|
| hkxhk | hk | hk | 11,039 | 10,287 | 6112 | 22.60% |
| nnxnp | nn | np | 18,052 | 15,912 | 6084 | 36.95% |
| lmxll | lm | ll | 19,762 | 17,566 | 5840 | 40.45% |
| Total | 48,853 | 43,765 | 18,036 | 100.00% |
| Linkage | Marker Number | Length | Average Distance | Max Gap |
|---|---|---|---|---|
| LG1 | 1202 | 59.93 | 0.05 | 3.73 |
| LG2 | 1255 | 75.23 | 0.06 | 4.98 |
| LG3 | 1125 | 65.73 | 0.06 | 4.97 |
| LG4 | 734 | 49.19 | 0.07 | 4.98 |
| LG5 | 690 | 61.60 | 0.09 | 4.14 |
| LG6 | 837 | 58.75 | 0.07 | 7.92 |
| LG7 | 796 | 44.67 | 0.06 | 4.56 |
| LG8 | 657 | 62.40 | 0.09 | 2.07 |
| LG9 | 370 | 91.09 | 0.25 | 7.92 |
| LG10 | 715 | 57.04 | 0.08 | 3.73 |
| LG11 | 994 | 55.79 | 0.06 | 1.65 |
| LG12 | 980 | 69.87 | 0.07 | 4.56 |
| LG13 | 364 | 54.61 | 0.15 | 7.49 |
| LG14 | 697 | 64.07 | 0.09 | 4.14 |
| LG15 | 357 | 30.17 | 0.08 | 2.89 |
| LG16 | 852 | 64.95 | 0.08 | 7.92 |
| LG17 | 547 | 53.72 | 0.10 | 2.07 |
| LG18 | 1017 | 75.33 | 0.07 | 8.77 |
| LG19 | 865 | 60.77 | 0.07 | 4.56 |
| LG20 | 309 | 43.81 | 0.14 | 3.72 |
| LG21 | 871 | 60.76 | 0.07 | 4.14 |
| LG22 | 621 | 59.53 | 0.10 | 5.40 |
| LG23 | 716 | 53.54 | 0.07 | 11.79 |
| LG24 | 164 | 23.17 | 0.14 | 4.97 |
| Total | 17,735 | 1395.72 | 0.08 | — |
| Group | Position | Peak Locus | Peak LOD | Expl (%) | Geno Number |
|---|---|---|---|---|---|
| 9 | 22.37–24.02 | hk2455 | 3.71 | 11–13.3 | 47 |
| 9 | 29.39 | hk2465 | 3.16 | 11.4 | 1 |
| 9 | 30.22 | hk2470 | 3.19 | 11.5 | 1 |
| 9 | 30.63 | hk2461 | 3.03 | 11 | 0 |
| 21 | 2.48–3.31 | np5073 | 3.09 | 10.9–11.2 | 4 |
| 21 | 3.31 | hk5416 | 3.16 | 11.2–11.4 | 60 |
| 21 | 3.72 | np5077 | 3.27 | 11.4–11.8 | 9 |
| 21 | 4.55 | hk5428 | 3.03 | 11 | 1 |
| 21 | 4.55–6.2 | hk5440 | 3.33 | 10.9–12 | 28 |
| 21 | 6.2 | np5097 | 3.53 | 11.9–12.7 | 17 |
| 21 | 6.2 | np5099 | 3.53 | 12–12.7 | 22 |
| 21 | 10.33 | np5112 | 3.01 | 10.9 | 16 |
| 21 | 13.64–14.46 | lm5172 | 3.06 | 10.9–11.1 | 16 |
| Gene | Gene_id | KEGG Pathway |
|---|---|---|
| chmp2a | A022229 | Endocytosis |
| snf8 | A022226 | Endocytosis |
| chmp6 | A022157 | Endocytosis |
| chmp1a | A009223 | Endocytosis |
| tat | A009234 | Phenylalanine, tyrosine and tryptophan biosynthesis; biosynthesis of amino acids; tyrosine metabolism |
| asah2 | A022051 | Sphingolipid metabolism |
| pten | A022010 | Inositol phosphate metabolism |
| arg1 | A022064 | Arginine and proline metabolism; biosynthesis of amino acids |
| heatr1 | A022065 | Ribosome biogenesis in eukaryotes |
| map2k6 | A022132 | MAPK signaling pathway |
| map3k14 | A022168 | MAPK signaling pathway; intestinal immune network for IgA production |
| Gene | Primer | Primer Sequences (5′–3′) |
|---|---|---|
| chmp2a | chmp2a-F | CGCTCAAGTCCAACAACAGC |
| chmp2a-R | TCGATGGCGTCGTTCATCAT | |
| snf8 | snf8-F | CAGGACGTGAGCCAAGATGA |
| snf8-R | CTCAGCCAGCTGCAGAACTA | |
| chmp6 | chmp6-F | CAGATCGGTAACCTGGAGCG |
| chmp6-R | TCGATGGACATCACCTCGTG | |
| chmp1a | chmp1a-F | GAAGGTCAAGAAGGCGTTGC |
| chmp1a-R | GGCGGTCTGGACTTTAGAGG | |
| tat | tat-F | TCAACGAGCTGTCCACCATC |
| tat-R | CGCTGAAGCAGGAAGACAGA | |
| asah2 | asah2-F | CGAGTGGCACATTCCTCTGT |
| asah2-R | GGCGACTTTGAAGACGTTGG | |
| pten | Pten-F | TGTGCGGAACGACATTACGA |
| Pten-R | TCCTCGCTCAACCACTTGTC | |
| arg1 | arg1-F | GATCCGTCACACAGGTCTCC |
| arg1-R | TAATCCTGCGGGTGGTTTCC | |
| heatr1 | heatr1-F | CTCAGGCCGTTCAGGAAGTT |
| heatr1-R | GTAGGTGGCGGCTTTGTAGT | |
| map2k6 | map2k6-F | AGATGTGAAGCCCTCCAACG |
| map2k6-R | AGCCTTTCTGGTTCGTCTCG | |
| map3k14 | map3k14-F | TGCAGGGCGAATGTCTAAGG |
| map3k14-R | GGAGCCACAGACAAGACTCC | |
| β-actin | β-actin-F | CTCTGTCTGGATCGGAGGCT |
| β-actin-R | GCTGAAGTTGTTGGGTGTTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Zhu, J.; Liu, S.; Liu, H.; Zhang, T.; Ye, T.; Lou, B.; Liu, F. QTL Mapping-Based Identification of Visceral White-Nodules Disease Resistance Genes in Larimichthys polyactis. Int. J. Mol. Sci. 2024, 25, 10872. https://doi.org/10.3390/ijms252010872
Li Q, Zhu J, Liu S, Liu H, Zhang T, Ye T, Lou B, Liu F. QTL Mapping-Based Identification of Visceral White-Nodules Disease Resistance Genes in Larimichthys polyactis. International Journal of Molecular Sciences. 2024; 25(20):10872. https://doi.org/10.3390/ijms252010872
Chicago/Turabian StyleLi, Qian, Jiajie Zhu, Sifang Liu, Haowen Liu, Tianle Zhang, Ting Ye, Bao Lou, and Feng Liu. 2024. "QTL Mapping-Based Identification of Visceral White-Nodules Disease Resistance Genes in Larimichthys polyactis" International Journal of Molecular Sciences 25, no. 20: 10872. https://doi.org/10.3390/ijms252010872
APA StyleLi, Q., Zhu, J., Liu, S., Liu, H., Zhang, T., Ye, T., Lou, B., & Liu, F. (2024). QTL Mapping-Based Identification of Visceral White-Nodules Disease Resistance Genes in Larimichthys polyactis. International Journal of Molecular Sciences, 25(20), 10872. https://doi.org/10.3390/ijms252010872