Abstract
The Northern snakehead (Channa argus) is a significant economic aquaculture species in China. Exhibiting sexual dimorphism in the growth rate between females and males, mono-sex breeding holds substantial value for aquaculture. This study employed GWAS and transcriptome analysis were applied to identify sex determination genomic regions and develop sex-specific markers. A total of 270 single-nucleotide polymorphisms (SNPs) and 31 insertion-deletions (InDels) were identified as being sexually dimorphic through GWAS and fixation index (Fst) scanning. Based on GWAS results, two sex-specific InDel markers were developed, effectively distinguishing genetic sex for XX females, XY males, and YY super-males via (polymerase chain reaction) PCR amplification. A major genomic segment of approximately 115 kb on chromosome 3 (Chr 03) was identified as the sex-determination region. A comparative transcriptome analysis of gonads for three sexes identified 158 overlapping differentially expressed genes (DEGs). Additionally, three sex-related candidate genes were identified near the sex determination region, including id2, sox11, and rnf144a. Further studies are required to elucidate the functions of these genes. Overall, two sex-specific InDel markers support a male heterogametic XX/XY sex-determination system in Northern snakeheads and three candidate genes offer new insights into sex determination and the evolution of sex chromosomes in teleost fish.
1. Introduction
The diversity and plasticity of sex determination systems in teleosts represent a fascinating area of study with profound implications for aquaculture, evolutionary biology, and conservation. With their range of sex chromosomes, sex-determining genes, and mechanisms of sex differentiation, teleosts offer unique insights into how sex determination systems evolve and adapt to various environmental pressures [1,2]. Fish gonadal development encompasses a series of intricate biological processes, including sex determination and differentiation. Fish sex determination can generally be classified into three types: genotypic sex determination (GSD), environmental sex determination (ESD), or a combination of both (GSD + ESD) [3]. Gonadal development initiates with sex determination. Through the influence of sex-determining genes, the genetic switch that triggers ovarian and testicular development is activated, subsequently inducing the downstream genes related to sex differentiation to regulate the expression of sex steroid synthase genes (ultimately determining the fate of the gonads), and their sex-determining genes remain unclear, making it challenging to fully understand the genetic basis of sex determination. However, advances in genomic analyses are helping to unravel the complex and plastic systems of sex determination in fish [4]. With the development of genomic tools combined with the application of quantitative genetics, more and more fish sex determination types and sex chromosomes have been demonstrated [4]. Fish species exhibit a wide range of sex determination modes, from simple genetic systems like XX/XY and ZZ/ZW to more complex systems involving multiple sex chromosomes or polygenic sex determination [5], such as XX/XY [6,7,8], ZW/ZZ [9,10], XX/XY1Y2 [11], X1X2Y [12], X1X1X2X2/X1X2Y [13,14], XX/XO and ZO/ZZ [15].
Sexual dimorphism in traits such as growth rate, maturity age, body size, and color pattern is highly significant in many cultured fish species and has direct implications for production performance [16,17]. Sex control in fish is an effective breeding strategy that allows for more efficient and sustainable fish farming, enhancing yield and economic outcomes, especially when employing mono-sex population production strategies [18,19,20,21]. The development and application of sex-linked molecular markers have become invaluable tools in aquaculture, facilitating the production of mono-sex populations, early sex identification, and the enhancement of sex-linked traits, all of which are increasingly being applied in aquaculture [22,23,24,25,26,27,28]. In certain farmed fish species, males exhibit faster growth rates than females, as observed in Nile tilapia (Oreochromis niloticus) [29], channel catfish (Ictalurus punctatus) [30], yellow catfish (Pelteobagrus fulvidraco) [31,32], and Lanzhou catfish (Silurus lanzhouensis) [33]. Conversely, in other farmed species, females outgrow males, such as the half-smooth tongue sole (Cynoglossus semilaevis) [34], rainbow trout (Oncorhynchus mykiss) [35], sea bass (Dicentrarchus labrax) [36], Europeaneel (Anguilla anguilla L.) [37], and Southern catfish (Silurus meridionalis) [38]. Understanding the genetic architecture of sex determination in aquaculture species is essential for improving the efficiency, sustainability, and precision of breeding programs [39]. By integrating these genetic insights into practical applications, production efficiency will not only be improved but also reliance on older, less sustainable methods like hormonal treatments will be reduced, leading to a more technologically advanced and ethical aquaculture industry. Therefore, research on the genetic mechanisms underlying sex determination holds significant value for the execution of fish production strategies and sex control breeding [40].
Developing and utilizing sex-specific markers in fish breeding is a game-changing advancement for aquaculture, allowing for more precise control over the sex of populations, leading to increased productivity, efficiency, and sustainability [2]. The development of sex-specific markers has significantly advanced aquaculture breeding over the past two decades, leading to more efficient and sustainable production systems [17]. Traditional techniques such as Restriction Fragment Length Polymorphism (RFLP), Amplified Fragment Length Polymorphism (AFLP), Random Amplified Polymorphic DNA (RAPD), and Simple Sequence Repeat (SSR) have been crucial in the identification of sex-specific markers in over 20 aquaculture species [16,17]. However, these methods are now considered to be time-consuming, labor-intensive, and less effective compared to modern genomic techniques. Based on next generation sequencing (NGS), SNP markers have become a cornerstone of modern aquaculture breeding programs, enabling more efficient, precise, and sustainable practices [17,41]. By facilitating early selection for desired traits, improving disease resistance, enhancing feed efficiency, and maintaining genetic diversity, SNP markers allow breeders to meet the growing demand for seafood in an environmentally and economically sustainable manner. For example, restriction site-associated DNA sequencing (RAD-seq) [42,43], Type IIB Restriction Site-Associated DNA Sequencing (2b-RAD) sequencing [24,44], and whole-genome resequencing (GWS) [45,46,47] have been used to identify sex-specific markers. Recently, whole-genome sequencing technology has been increasingly applied for sex-specific marker screening in a growing number of farmed fish species [48,49], including largemouth bass (Micropterus Salmoides L.) [47], blunt-snout bream (Megalobrama amblycephala) [46], army fish (Spinibarbus hollandi) [45], Spinibarbus hollandi [23], Takifugu bimaculatus [49], blue tilapia (Oreochromis aureus) [50], yellowstripe goby (Mugilogobius chulae) [51], Russian sturgeon (Acipenser gueldenstaedtii) [26], turbot (Scophthalmus maximus) [52], and spotted knifejaw (Oplegnathus punctatus) [12].
The Northern snakehead (Channa argus) is a renowned aquaculture species with significant economic value. According to the China Fishery Statistical Yearbook, national aquaculture production of Northern snakehead surpassed 605,438 tons in 2023. Male individuals exhibit a markedly faster growth rate than females, particularly after the first year, with males reaching an average weight of more than double that of females. Due to this rapid growth, males are favored, growing nearly twice as fast as females. Consequently, all-male mono-sex breeding could substantially enhance breeding yield and economic benefits in Northern snakehead aquacultures [20,53,54]. Understanding the genetic and environmental mechanisms governing sex regulation and differentiation in Northern snakeheads, aquaculture operations can develop single-sex breeding strategies to optimize growth, improve yield, and control reproduction. A rapid and accurate sex-specific marker is indispensable for aquaculture breeding programs focused on producing all-male populations and identifying sex-reversed individuals. Previous research has developed several sex markers and fine-mapped the sex quantitative trait locus (QTL) [53,55,56,57]. However, the lack of understanding of sex chromosomes and the mechanistic action of the sex-determining genomic region presents a major obstacle in the development of efficient sex-specific markers for identifying YY super-males. In this study, we aimed to identify the sex determination region and putative sex-determining genes, as well as to develop genetic sex markers for the Northern snakehead. We conducted whole-genome resequencing on 59 Northern snakehead individuals. The GWAS results revealed a specific sex-determining chromosomal region on Chr03. By integrating the GWAS and RNA-seq results, we identified key candidate sex-determining genes. Moreover, two sex-specific InDel markers were validated for practical verification of genetic sex in XX, XY, and YY individuals, confirming an XX/XY sex determination system in Northern snakeheads. This effort will not only contribute to the theoretical understanding of sex differentiation but also provide practical tools for aquaculture, enabling more efficient and sustainable breeding strategies in regard to Northern snakeheads.
2. Results
2.1. Resequencing, Variants Calling and Population Structure Analyses
The genotyping of 59 adult Northern snakeheads was conducted using whole-genome resequencing. Approximately 871.3 Gb of raw data were generated using the Illumina NovaSeq platform. Following quality control, 864.3 Gb of clean reads were obtained. For the 59 individuals, the average amount of clean data per sample was 14.6 Gb. Based on the genome size, the mean alignment rate was 98.5% and the mean sequencing depth was 21.4×. A total of 4,613,089 SNPs and 834,465 InDels were identified using HaplotypeCaller in GATK. Following additional quality control using Plink v. 1.90, which included filtering by a missing call rate and minor allele frequency (MAF), the high-quality site of 1,087,459 SNPs and 159,324 InDels were retained for subsequent GWAS. The SNP density map in Figure 1 showed that the number of SNPs per chromosome ranged from 11,125 on chromosome 19, which had the fewest SNPs, to 50,096 on chromosome 1, which had the most. SNPs were uniformly distributed across the chromosomes, with the average density exceeding 7318 loci per Mb.
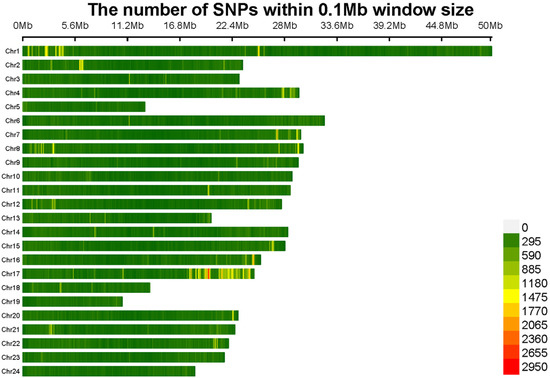
Figure 1.
High-quality SNPs were mapped to 24 C. argus chromosomes, with the gradient color ranging from green to red to indicate increasing SNP density at 1 Mb intervals.
2.2. Sex-Linked Region and Candidate Gene of C. argus
A total of 270 SNPs were found to be significantly associated with sex on chromosome 03 (Chr 03) (Figure 2A, Table S1). These SNPs accounted for 3.97% to 98.97% of the phenotypic variance explained (PVE). Similarly, 31 InDels were identified as significantly associated with sex on Chr 03 (Figure 2C, Table S1). These InDels accounted for 7.30% to 42.61% of the PVE. The Q-Q plots and lambda (λ) values confirm that the GWAS model used is well calibrated and appropriate for analyzing the SNP and InDel data (Figure 2B,D).
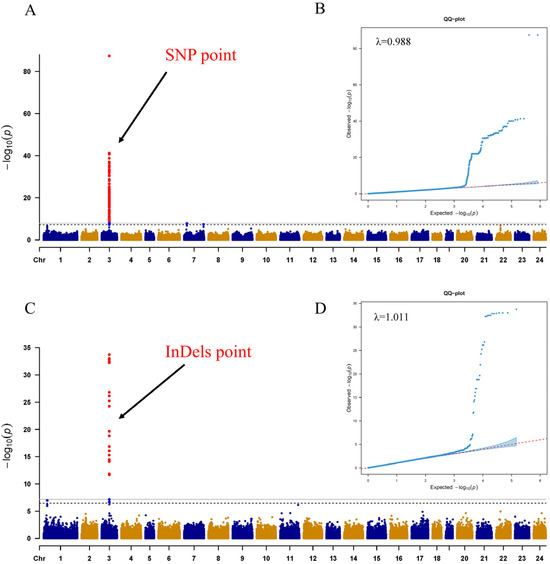
Figure 2.
Results of GWAS analyzing sex characteristics. (A,C) show Manhattan plots of the optimal patterns of SNPs and InDels, along with Q-Q plots for SNPs, InDels, and corresponding λ values. The x-axis represents the chromosome number (Chr) while the y-axis represents the −log10(P) value for each SNP. Red dots indicate SNP signals exceeding the significance threshold, whereas blue and yellow dots fall below the threshold. The red points indicated by the black arrows represent significant loci. (B,D) present Q-Q plots of the optimal patterns of SNPs and InDels.
All SNPs were clustered within a 192 Kb segment on Chr 03 (11,363,773 bp to 11,555,658 bp) (physical location of the start and end in the reference genome). Although two uncharacterized proteins and the rnf144a protein-encoding gene were identified among the 270 SNPs, their lack of involvement highlights the functions of sex determination or differentiation. Additionally, 31 InDels were significantly associated with sex within a 143 Kb region on Chr 03 (11,390,044 bp to 11,534,257 bp), with many loci conforming to the XX/XY pattern. Furthermore, the identification of 21 potential genes through scanning the ±250 Kb regions around genome-wide significant variants, shown in linkage disequilibrium (LD) blocks (Figure 3), is annotated in Table 1. The genes closest to the sex-determining region are EVM0022341 (uncharacterized protein), EVM0015303 (Ring finger protein), and EVM0022102 (uncharacterized protein), located at Table 1. Interestingly, Sox11 and Id2 were located downstream and upstream of the sex-determining region.
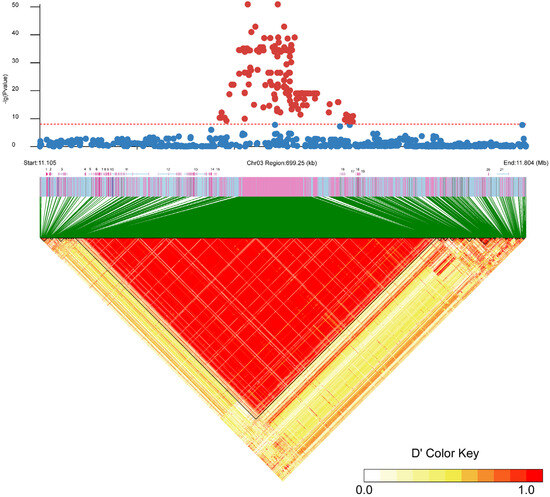
Figure 3.
Fine mapping on Chr 03 revealed the association between SNPs and sex as well as the combined linkage disequilibrium in the sex-related region. Red dots indicate SNP signals exceeding the significance threshold, whereas blue dots fall below the threshold. The axis below the chart represents the region where candidate genes are annotated.

Table 1.
The candidate genes in the sex-determining region on Chr 03 for Northern snakehead.
2.3. Selection and Test of the Sex-Specific Markers
Fst values (Figure 4A,B) and PCA analysis (Figure 5A,B) for each SNP and InDel between males and females confirmed the effectiveness of the primer design site. Subsequently, 10 pairs of male-specific primers were designed by screening 10 sexually dimorphic InDels from 31 identified InDels. After the preliminary PCR amplification screening, we successfully obtained two pairs of primers which effectively distinguished the C. argus of XX-F (genetic sex is XX and phenotypic sex is female), XY-M (genetic sex is XY and phenotypic sex is male), and YY-M (genetic sex is YY and phenotypic sex is male) (Figure 6). For both primers, all samples exhibited clear bands: males displayed two bands, while females and super-males showed a single band, located above and below, respectively.
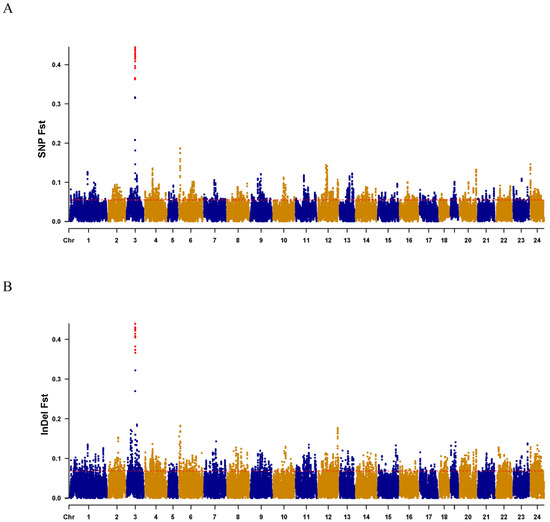
Figure 4.
Result of Fst scan for females and males on chromosomes. (A,B), fine mapping on Chr 3 shows the combination of associations of SNPs and InDels with sex and Fst, respectively. Candidate sign inferred by Fst (>0.3490) are marked by red dots, whereas blue and yellow dots fall below the Fst (>0.3490). The x-axis indicates the Chr numbers. The y-axis is the Fst value of the female and male of the SNPs or InDels.
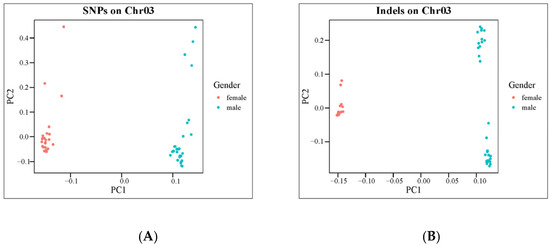
Figure 5.
Result of PCA for females and males with traits. (A,B), the PCA of 59 individuals to distinguish two sexes using SNPs or InDels on Chr03. The red and blue dots represent females and males, respectively.

Figure 6.
Results of PCR products for females, males, and super-males with two primers. The band above the green frame denotes female samples (XX), the red frame represents male samples (XY), and the blue frame signifies super-male samples (YY) within the Northern snakehead population.
2.4. Transcriptome Analysis
A total of 10,672 significant DEGs were identified in XY-M vs. XX-F, 237 were identified in XY-M vs. YY-M, and 12,347 were identified in YY-M vs. XX-F using RNA-Seq. In XY-M vs. XX-F, 5985 genes were upregulated and 7598 were downregulated; in XY-M vs. YY-M, 70 genes were upregulated and 167 were downregulated; in YY-M vs. XX-F, 5558 genes were upregulated and 6789 were downregulated (Figure 7A–C). A total of 158 conserved DEGs in representative glandular tissues were identified by overlapping the differentially expressed genes between groups using Venn diagrams (Figure 7D). These 158 DEGs were collectively enriched in 384 significant GO terms. Figure 8A shows the top GO enrichment, where biological pathways related to signal transduction regulation were significantly enriched, including neutral lipid metabolic process, lipid transport, regulation of monoatomic ion transmembrane transport, signaling receptor regulator activity, monoatomic anion transmembrane transporter activity, protein kinase regulator activity, transcription corepressor activity, and kinase regulator activity. In the KEGG enrichment analysis (Figure 8B), 20 pathways were significantly enriched. Fatty Acid Elongation, Fatty Acid Metabolism, and Fatty Acid Biosynthesis are important for the growth and function of gonadal cells and influence the production of sex hormones. The biosynthesis of Unsaturated Fatty Acids and the Adipocytokine Signaling Pathway play pivotal roles in energy metabolism and reproductive function, regulating gonadal activity and the secretion of sex hormones. Furthermore, the Progesterone-mediated Oocyte Maturation (POM) and mammalian target of rapamycin (mTOR) Signaling Pathways are of particular importance, with POM being directly involved in ovarian function, regulating the process of oocyte maturation and ovulation. The mTOR pathway is critical for cell proliferation and metabolic regulation, significantly impacting gonadal development and germ cell production.
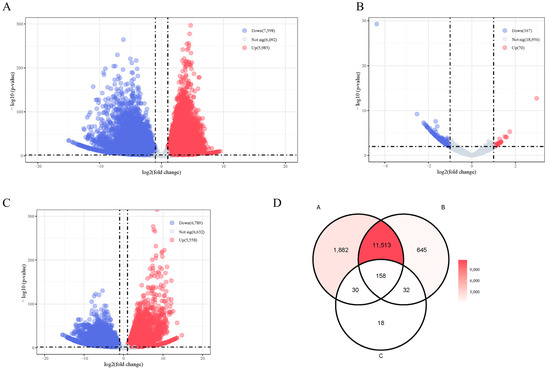
Figure 7.
Transcriptome analyses based on gonads in Northern snakeheads. (A–C) Volcano plot of DEGs shows all the individuals’ gonads within RNA-seq. Red and blue dots indicate significantly upregulated and downregulated genes, respectively (|log2FC| ≥ 1 and p < 0.01). Gray dots indicate genes not significantly differentially expressed between XX vs. XY, XY vs. YY, and XX vs. YY. (D) Venn maps of DEGs obtained from RNA-seq based on gonads, whereas A is XX vs. XY, B is. XY vs. YY, and C is XX vs. YY. The overlapping intersection is compared with the RNA-seq result, and the resulting region of 158 represents the final conservative DEGs.
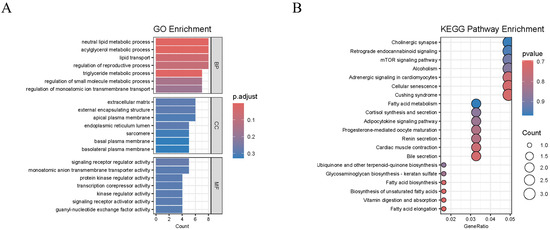
Figure 8.
Functional enrichment analysis of overlapping DEGs. (A) presents the top 21 significantly enriched Gene Ontology (GO) terms for the 158 conserved DEGs. (B) shows the 20 significantly enriched KEGG pathways for the 158 conserved DEGs.
The qPCR results showed that Amh, Amhr2, Ar, Cyp17a, and Dmrt1 were significantly more highly expressed in XY-M and YY-M spermatheca compared to XX-F (Figure 9A), while the ovarian differentiation-related genes Bmp15, Cyp19a1a, Er, Figla, and Foxl2 were significantly under expressed in XY-M and YY-M spermatheca (Figure 9B). These results are consistent with the expression patterns observed in the transcriptomic analysis, which demonstrated clear sexual dimorphism, although the relative expression levels varied slightly. Further correlation analysis was conducted between the qRT-PCR results of DEGs related to spermathecal and ovarian differentiation and the transcriptomic data. Significant correlations were observed for the XY-M vs. XX-F and XY-M vs. YY-M groups, with R² values of 0.8835 and 0.9008, respectively, indicating the high reliability of the transcriptomic data.
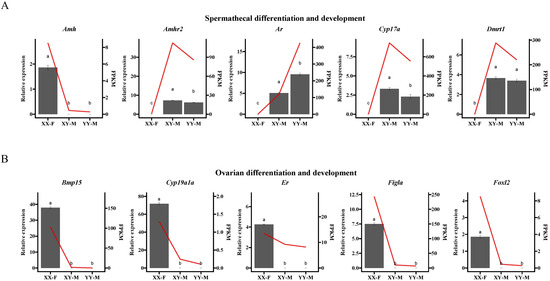
Figure 9.
qPCR validation for (A) five genes within the spermathecal differentiation and development DEGs of gonads RNA-seq and (B) five genes within the ovarian differentiation and development DEGs of gonads RNA-seq. The gray bars represent the relative expression levels of the gene, corresponding to the y-axis on the left side of each figure, and are presented as mean ± standard deviation. The red line indicates the gene expression levels from RNA-seq data, represented in FPKM, and corresponds to the y-axis on the right side of each figure, displayed as the mean. The significance of the qPCR results between the XX-F, XY-M, and YY-M individuals is indicated by letters. The x axis that XX-F (genetic sex is XX and phenotypic sex is female), XY-M (genetic sex is XY and phenotypic sex is male) and YY-M (genetic sex is YY and phenotypic sex is male).
2.5. Identification of Candidate Genes by GWAS and RNA-Seq
A total of 21 potential genes were extracted and annotated in the 250 kb gene candidate region near the locus of significance SNPs. Also, to identify sex-biased genes within the candidate sex-determining regions, we compared these genes with the DEGs in the RNA-Seq dataset of sexually active gonads. The results showed that it contained 10 female-biased transcripts and seven male-biased transcripts (Figure 10B, Table S2). The female-biased protein-coding genes identified were ywhaq, iah1, asap2, id2, sox11, dnajc30 and grhl1. The male-biased genes included klf11, cmpk2, rsad2, dnaaf2, itgb1bp1, and kidins220, along with other uncharacterized genes. The finding of these genes will assist in the foundation of future in-depth studies of the molecular mechanisms of Northern snakehead populations. We conducted a functional exploration of candidate genes by GO enrichment and a Kegg pathway enrichment analysis (Figure 10A,C). The results revealed that several GO enrichment terms might be associated with sex differentiation or development, including DP-dissociation Inhibitor Activity, Lipid Droplet, Positive Regulation of Transmembrane Receptor Protein Serine/Threonine Kinase Signaling Pathways, 2-acylglycerol-3-phosphate O-acyltransferase Activity and mRNA Cleavage Factor Complexes and mRNA Cleavage and Polyadenylation Specificity Factor Complexes. Additionally, five KEGG pathways were significantly enriched among these candidate genes, with Hippo Signaling Pathway—Fly, Oocyte Meiosis, TGF-beta Signaling Pathway, and Glycerophospholipid Metabolism being potentially associated with sex regulation.
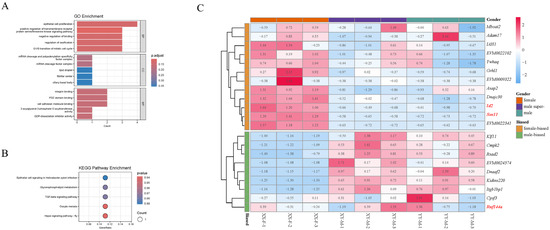
Figure 10.
Final candidate genes and their functional enrichment analysis. (A,B) Enriched GO terms and KEGG pathways for candidate genes. (C) Heatmap analysis of candidate genes differentially expressed between XX-F, XY-M and YY-M Northern snakeheads. XX-F, female gonad individuals. XY-M, male gonad individuals. YY-M, super-male gonad individuals. Matrix blocks in red are upregulated while those in blue are downregulated in gens. The red and bold name is interested in candidate gene.
3. Discussion
In recent years, WGS has become a powerful tool in the development of sex-specific markers due to its broad coverage, ability to detect large sex-linked fragments, and capability of screening vast numbers of genetic markers, and it has been increasingly applied to the development of sex-specific markers in a growing number of fish species [45,46,49]. Based on resequencing, several new analytical strategies have been developed for sex-specific sequence screening, including the investigation of sexually dimorphic SNPs and the comparing of sequencing depth, GWAS, and Fst analyses [10,58,59]. We conducted GWAS on sex traits in Northern snakeheads and identified a total of 270 SNPs and 31 InDels associated with the sex phenotype. All these phenotype-associated SNPs and InDels exceeded the significance threshold. Among the genes associated with these molecular markers, three were associated with SNPs, two with InDels, and two genes were found to be associated with both SNPs and InDels. Subsequent studies have shown that the genes EVM0022341 and rnf144a are not directly related to sex determination or differentiation functions. Interestingly, sox11 and id2 were found within their sex determination candidate window.
Clarifying the sex determination mechanism in Northern snakeheads, including the confirmation of its XX/XY system and the identification of a sex-determining region through QTL mapping, lays the groundwork for mono-sex breeding strategies [53,55,56,57]. The development of sex-specific markers using AFLP [60] and SSR [61] methods presents several disadvantages: they are costly, labor-intensive, and time-consuming. First, while SSR markers have historically been useful for identifying sex-specific markers, the process of isolating and screening them is time-consuming and labor-intensive. SNP, however, is a powerful, versatile, and efficient tool that can better facilitate the identification and utilization of snakeheads [62]. The use of specific primers designed from GWAS results for PCR validation is a key step in confirming the sex-specificity of identified genetic fragments to ensure the accuracy of sex-linked markers and their applicability in breeding programs, which ultimately contributes to more efficient and precise sex control in aquaculture species. Based on 31 sex-related InDels, 10 pairs of primers were designed. Finally, two InDel sex-linked markers were obtained that can successfully distinguish XX, XY, and YY fish. The results suggest that this method offers advantages in terms of precision and lower sample size requirements. This method has been gradually applied to assist in mapping sex-specific sequences in aquatic animals, such as largemouth bass (Micropterus Salmoides L.) [47], blunt-snout bream (Megalobrama amblycephala) [46], army fish (Spinibarbus hollandi) [45], Spinibarbus hollandi [23], Takifugu bimaculatus [49], Pacific abalone (Haliotis discus) [48], and Takifugu bimaculatus [49]. Sun-developed universal sex markers (C. maculata ♀ × C. argus ♂) are applicable to two species, C. argus and C. maculata, and their hybrids [56]. Although these markers are effective in distinguishing genetic males and females in snakehead species, they cannot distinguish YY fish. Additionally, Dai [63] have developed a molecular marker for sex identification in albino snakeheads which can differentiate YY fish from albino snakeheads and also distinguish between male and female wild snakeheads. However, there is no direct evidence for distinguishing XX, XY, and YY fish from non-albino snakeheads. Sex markers in these species usually offer technical advantages. Wen employed a single analytical method to display the Fst values between males and females across the entire genome, identifying a substantial potential sex differentiation region spanning 6 Mb and encompassing 275 genes, representing a relatively broad research scope [46]. In the present study, we used both GWAS and Fst to identify sex-linked genomic segments. A total of 270 SNPs and 31 InDels were detected as sexually dimorphic regions spanning a narrow 115 Kb segment on Chr03, providing a valuable resource for developing diagnostic markers of genetic sex in Northern snakeheads. In general, these new sex markers establish a strong foundation for identifying the genetic sex and sex chromosomes of Northern snakeheads and provide a highly efficient technological approach for mono-sex breeding in Northern snakehead aquaculture, which holds significant potential for enhancing efficiency and productivity.
Among the numerous sex-specific markers identified in fish, some are located on sex-determining genes. For example, the domain gene on the Y chromosome (DMY) gene was found in the sex-determining region of Medaka (Oryzias latipes) [64], the dmrt1 gene was identified based on a male-specific 15 bp deletion in large yellow croaker (Larimichthys crocea) [65], the dmrt1 gene in spotted scat (Scatophagus argus) [66], and the amh gene in mandarin fish (Siniperca chuatsi) [67]. However, in some fish species, sex-determining genes have not been identified despite the presence of sex markers, such as in spotted knifejaw [68], blotched snakehead [69], blunt-snout bream (Megalobrama amblycephala) [46], albino snakeheads [63] and largemouth bass [51]. In our previous study, several genes, including sox11, were identified in the sex QTL region using a high-density genetic map and QTL analysis [53]. In this study, based on a high-quality reference genome and GWS combined with GWAS analysis, we accurately identified the sex-determining region on Chr 03 within a 115 Kb segment, facilitating the screening of candidate genes. However, no previously identified sex-determination genes were found in the sex-determining region based on the annotation results. The genes closest to the sex-determining region are EVM0022341 (uncharacterized protein), EVM0015303 (Ring finger protein), and EVM0022102 (uncharacterized protein). Additionally, sox11 and id2 were near the window of sexual differentiation.
Recently, fish have been observed to exhibit sex plasticity, displaying a diverse array of sex determination mechanisms. As high-throughput sequencing technology has become more sophisticated, numerous candidate genes have been identified across various teleost fish species. For example, the amh in Sebastes rockfish [70], the dmrt1 in Scatophagus argus [66], the sox11a gene in Oreochromis niloticus [71], and five sex-related genes in largemouth bass [72]. In this study, the sox11 protein-coding gene was annotated from the ~115 Kb candidate SD region on Chr 03. Sox11 is a transcriptional activator that belongs to the SOX gene family and is crucial for several developmental processes, including gonadal differentiation, spermatogenesis, vitellogenesis, and oogenesis in fish [71,73,74]. This gene corresponds to the candidate gene sox11 identified in the fine mapping of the sex QTL [53]. Additionally, RNA-Seq analysis revealed that the sox11 gene is the most highly expressed in females among the three gonadal tissues, indicating that the sox11 gene may play a complex role in the sexual differentiation of Northern snakeheads. Similar results were obtained in Collichthys lucidus [75].
To date, several new candidate master sex-determining (MSD) genes have been discovered that are not directly related to gonadal development. For example, the Y-chromosome (sdY) gene in Atlantic Salmon [76,77,78,79], the bcar1 gene in channel catfish [80], the rnf183 gene in L. polyactis [81], and the id2bbY gene in Arapaima gigas [82], Interestingly, we characterized two protein-coding genes, rnf144a and id2, consistent with findings from a recent study [56], in candidate sex-determining regions that have not yet been functionally confirmed. id2 proteins belong to the ID family and primarily regulate cell differentiation and proliferation by inhibiting the function of bHLH (basic Helix–Loop–Helix) transcription factors. Evidence has demonstrated that the id2 gene plays a crucial role in male meiosis and spermatid development in mice by regulating the differentiation of testicular supporting cells through the inhibition of bHLH transcription factors [83]. The id2bbY gene, a homolog of id2 in Arapaima gigas, is thought to be involved in sex determination [82]. rnf144a, an E3 ubiquitin ligase for DNA-PKcs, promotes apoptosis during DNA damage and is strongly correlated with body conformation traits in goats [84]. Although it has rarely been reported in teleost fish, a recent study on RNF family members showed that rnf183 is a sex-determining gene in little yellow croaker [81]. In our study, the id2 genes showed biased expression in both females and males in RNA-seq, whereas rnf144a exhibited inconsistent expression trends. Therefore, it is reasonable to assume that the id2 or rnf144a gene might be a candidate MSD gene in C. argus. These genes represent valuable genetic resources for studying the sexual evolution of the Northern snakehead and other teleost fish. By focusing on these genes, researchers can gain insights into the origin and evolution of primitive sex chromosomes, the genetic mechanisms driving sex determination, and how these systems have evolved over time. This knowledge has important implications for both evolutionary biology and practical applications in aquaculture, particularly in the development of sustainable and efficient mono-sex breeding strategies.
By combining the RNA-seq data from three gonadal tissues of C. argus, the candidate genes id2 and rnf144a were analyzed in the sex determination region, and it was found that they exhibited biased expression related to sex determination in C. argus. This finding warrants further investigation, particularly focusing on the roles and regulatory mechanisms within the sex determination (SD) system. By employing gene-editing technologies and gene expression profiling in gonads during the critical stages of sex determination, we can uncover the precise roles of the master sex-determining genes in Northern snakeheads in future works. This will not only enhance our understanding of sex differentiation but also pave the way for developing targeted genetic tools to control sex ratios in aquaculture, leading to more sustainable and productive breeding programs.
4. Materials and Methods
4.1. Ethics Statement
The sampling process of Channa argus adhered to the experimental animal welfare guidelines and was approved by the Experimental Animal Welfare Ethics Committee of the Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences.
4.2. Sample Sources and DNA Extraction
Adult Northern snakehead (Figure 11) samples were originally collected from Weishanhu Lake (Shandong Province) and Fuyuan City (Heilongjiang Province) in China (Figure 12). These fish were cultivated at the Nanhai Bai-rong Aquatic Varieties Limited Company in Foshan City, Guangdong Province, China. The phenotypic sex of all individuals was determined by examining the gonads. A total of 59 fish (26 females and 33 males) were randomly selected, and fin tissue was collected from each individual, immediately immersed in absolute ethanol, and then preserved at −20 °C. Genomic DNA was extracted from the fin clips using a Qiagen DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA, USA) according to the manufacturer’s protocol. DNA quality and concentration were assessed using a NanoDrop One spectrophotometer (Thermo Scientific, 168 Third Avenue, Waltham, MA, USA) and 1% agarose gel electrophoresis.

Figure 11.
Individuals in C. argus.
Figure 12.
Map of sampling sites for C. argus. The red dot shows the specific sampling locations.
4.3. Whole Genome Resequencing and Variation Detection
Genomic DNA from 59 individuals was used for whole-genome resequencing, which was performed on the Illumina NovaSeq platform in PE150 mode. The raw reads obtained from the high-throughput sequencing pipelines were quality controlled using FastQC software (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/, accessed on 6 October 2024). Paired-end data with adapter contamination and low-quality bases were trimmed to obtain clean reads using fastp (v. 0.21.0) with default parameters [85]. The clean reads were aligned to the Northern snakehead reference genome (ASM1899790v1) using BWA (v. 0.7.17) [86]. The resulting SAM files were converted to BAM files and sorted using SAMtools (v1.15.1) [87]. Duplicates were removed with the MarkDuplicates tool from the Picard toolkit v2.27.5. SNPs and InDels were identified using the Genome Analysis Toolkit (GATK) package (v. 4.3.0.0), following handle scripts, including the merging of each sample gVCF files and combining them into a multi-sample gVCF file by HaplotypeCaller and CombineGVCFs tools; Genotyping multi-sample gVCF file with GenotypeGVCFs methods, finally the SNP and InDel after filtering the variants following the parameters “QD < 2.0 || MQ < 40.0 || FS > 60.0 || SOR > 3.0 || MQRankSum < −12.5 || ReadPosRankSum < −8.0”. The resulting SNPs and InDels were further filtered using PLINK (v. 1.90) [88], excluding sites with missingness per individual > 0.1 and missingness per marker > 0.05.
4.4. Sex-Determining Region and Loci Screening Using GWAS
A genome-wide association analysis of the sex phenotype data and genotype data was conducted using a mixed linear model (MLM) in GEMMA (v. 0.98.5) software, based on the filtered SNPs and InDels [89]. The MLM is represented by the following equation: . Here, Y represents the observation vector of phenotypes, X is the design matrix of fixed effects, including the first three principal components and sex, and β is the vector of fixed-effect coefficients, often used to explain global, systemic impacts. Z is the design matrix of random effects, μ is the vector of random-effect coefficients (reflecting random differences between individuals as well as genetic background or population structures), and e represents the residual effects [90].
In the present study, we used the phenotypic characteristics of males and females as a basis for identifying genes in sex-determining regions. To ensure the validity of the optimal GWAS mode analysis, we first used a genomic inflation factor (λ) value close to 1.00. [91]. Quantile-quantile (Q-Q) plots and a Manhattan plot of significant SNPs or InDels were visualized by CMplot [92]. The genome-wide significance and suggestive association thresholds were set at 0.01/N and 0.05/N, respectively, where N represents the number of SNPs and InDels used in the GWAS analysis. These thresholds were established according to the Bonferroni correction method [93].
4.5. Sex Candidate Gene and Region Identify by Fst
The sex-linked genomic segment in Northern snakeheads was pinpointed through GWAS utilizing filtered SNPs. The Fst [94] between females and males was also calculated at each SNP, and the results were plotted using a CMplot to investigate sex-linked genomic regions. The significance of the association between loci and phenotypic sex was determined using the resulting threshold values, which were then plotted to further examine sex-linked genomic regions. To identify candidate genes, LDBlockShow (v. 1.40) was employed to generate linkage disequilibrium (LD) blocks and construct haplotype maps for the 250 kb regions upstream and downstream of each significantly associated SNP [95]. Subsequently, candidate genes within the delineated haplotype block regions were precisely annotated using BLAST against the SwissProt protein database.
4.6. Male-Specific Markers Development and Validation through InDel Region
We used a script to identify samples with sexually dimorphic InDels sites larger than 30 bp, and their validation was performed by aligning the resequencing reads from female and male samples to the reference genome. The selected data were identified as candidate regions using IGV (http://www.broadinstitute.org/software/igv/, accessed on 6 October 2024) for following PCR primers (Tables S3 and S4). These primers were designed based on the genomic sequences using Primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi, accessed on 6 October 2024), and these primers were then synthesized by Tianyi Huiyuan Biotech Co., Ltd. (Guangzhou, China). To verify the specificity of the primers, routine PCR amplification was conducted using the primer pairs synthesized in bulk by the aforementioned company. The PCR reaction was conducted in a total volume of 20 μL, consisting of 10 μL of Taq Mix, 0.5 μL of DNA template, 0.8 μL of each forward and reverse primer, and 7.9 μL of deionized water. The PCR conditions included an initial denaturation at 95 °C for 2 min, followed by 39 cycles of 35 s at 95 °C for denaturation, 35 s for annealing at the Tm, and 40 s at 72 °C for extension, with a final extension at 72 °C for 10 min. The amplified product length was evaluated by separate 8% polyacrylamide gels on 100 samples, including 43 females and 57 males, from randomly selected groups.
4.7. RNA Extraction, Sequencing, and Analysis of Sex-Biased Genes
RNA was extracted from the gonads of three Channa argus genotypes (XY-M, XX-F, and YY-M) using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The quality and concentration of RNA from each sample were assessed before being sent to Beijing Novogene (Beijing, China) for library construction and paired-end sequencing using the Illumina HiSeq 2000 platform (Illumina, San Diego, CA, USA). All raw sequencing data have been deposited in the NCBI database under accession number PRJNA895982.
Raw data were processed using FastQC (v. 0.11.8) to remove adapters, bad-quality reads, and reads that carried ambiguous bases (N), resulting in high-quality clean data. The clean reads were then aligned to the Northern snakehead reference genome (GenBank assembly accession No. GCA_003845795.1) using HISAT2 (v. 2.1.0) [96]. Transcript expression levels were quantified using the fragments per kilobase of transcript per million mapped reads (FPKM) method [97]. Differential expression analysis between sample groups was performed using DESeq2 (v. 1.36.0) to identify differentially expressed genes (DEGs) [98]. The false discovery rate (FDR) was controlled by adjusting p-values, with significance thresholds being set at |log2 fold change| ≥ 1 and p < 0.01 [98]. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were conducted using the ClusterProfiler package (v. 3.18.1) in R [99,100,101]. Candidate genes identified through GWAS were compared with the DEGs to identify overlapping genes. These overlapping genes were clustered based on Pearson’s correlation and Ward’s method using the R function hclust and subsequently visualized as heatmaps with the pheatmap package (v. 1.0.12) [102].
4.8. Validation of the Candidate Genes in RNA-Seq by qPCR
We designed primer pairs targeting 10 sex-related genes and employed quantitative PCR (qPCR) to validate the reliability of the RNA sequencing results. The reaction mixture (total volume of 20 μL) was prepared following the manufacturer’s instructions (ABI, Tampa, FL, USA) and included 10 μL of 2×SYBR Green® Real-Time PCR Master Mix (Toyobo, Osaka, Japan), 0.8 μL of paired primers, 0.5 μL of cDNA template, and 7.9 μL of sterile water. The following PCR conditions were employed for amplification: three biological replicates were performed and then standardized using C. argus β-actin as an internal reference [103]. Relative gene expression levels were calculated using the 2−ΔΔCt method [104]. One-way ANOVA was conducted using SPSS 22.0 to assess the significance of gene expression differences between groups, with p < 0.05 being considered statistically significant.
5. Conclusions
In the present study, we identified sexually linked gene fragments on Chr03 and developed two sex-specific markers. Additionally, we observed many growth-related candidate genes located near the associated SNPs. These findings provide a foundation and essential tools for further exploration of the molecular mechanisms of sex determination, ultimately facilitating advancements in the sex control breeding of Northern snakehead populations.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms252010889/s1.
Author Contributions
Conceptualization, H.L., J.Z. (Jin Zhang), X.Z., K.C. and J.Z. (Jian Zhao); methodology, H.L. and J.Z. (Jin Zhang); formal analysis, H.L.; investigation, T.C., W.X., M.O., Q.L. and S.F.; data curation, T.C., W.X., M.O., Q.L. and S.F.; writing—original draft preparation, H.L.; writing—review and editing, J.Z. (Jin Zhang); visualization, J.Z. (Jin Zhang); supervision, K.C., X.Z., M.O. and J.Z. (Jian Zhao); project administration, M.O.; funding acquisition, M.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the China Agriculture Research System of MOF and MARA (CARS-46); the Central Public-interest Scientific Institution Basal Research Fund, CAFS (2023TD37, 2023SJHX2); the National Natural Science Foundation of China (32373127); and the Guangdong Province Rural Revitalization Strategy Special Fund (2022-SPY-00-016, 2023-SJS-00-001); The Special Financial Fund of Foshan in 2024—Cooperation project for high-level agricultural science and technology demonstration city construction in Guangdong Province, the Basic and Applied Basic Research Foundation of Guangdong Province (2024A1515030165).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Ethics Committee of the Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences (Approval ID: LAEC-PRFRI-2022-06-01; Date: 1 June 2022). All fish were humanely sacrificed according to the relevant guidelines.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data analyzed in this study can be downloaded from the National Center for Biotechnology Information (NCBI) databases (PRJNA895982).
Acknowledgments
The authors sincerely thank Boxiang Chen for his help in our fish provision and storage.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Guiguen, Y.; Fostier, A.; Herpin, A. Sex Determination and Differentiation in Fish. In Sex Control in Aquaculture; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 35–63. ISBN 978-1-119-12729-1. [Google Scholar]
- Kobayashi, Y.; Nagahama, Y.; Nakamura, M. Diversity and Plasticity of Sex Determination and Differentiation in Fishes. Sex. Dev. 2012, 7, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.-L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex Determination: Why so Many Ways of Doing It? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed]
- Sember, A.; Nguyen, P.; Perez, M.F.; Altmanová, M.; Ráb, P.; Cioffi, M.d.B. Multiple Sex Chromosomes in Teleost Fishes from a Cytogenetic Perspective: State of the Art and Future Challenges. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200098. [Google Scholar] [CrossRef] [PubMed]
- Kitano, J.; Ansai, S.; Takehana, Y.; Yamamoto, Y. Diversity and Convergence of Sex-Determination Mechanisms in Teleost Fish. Annu. Rev. Anim. Biosci. 2024, 12, 233–259. [Google Scholar] [CrossRef]
- de Freitas, N.L.; Al-Rikabi, A.B.H.; Bertollo, L.A.C.; Ezaz, T.; Yano, C.F.; de Oliveira, E.A.; Hatanaka, T.; de Bello Cioffi, M. Early Stages of XY Sex Chromosomes Differentiation in the Fish Hoplias malabaricus (Characiformes, Erythrinidae) Revealed by DNA Repeats Accumulation. Curr. Genom. 2018, 19, 216–226. [Google Scholar] [CrossRef]
- Xue, L.; Gao, Y.; Wu, M.; Tian, T.; Fan, H.; Huang, Y.; Huang, Z.; Li, D.; Xu, L. Telomere-to-Telomere Assembly of a Fish Y Chromosome Reveals the Origin of a Young Sex Chromosome Pair. Genome Biol. 2021, 22, 203. [Google Scholar] [CrossRef]
- Nguyen, D.H.M.; Ponjarat, J.; Laopichienpong, N.; Panthum, T.; Singchat, W.; Ahmad, S.F.; Kraichak, E.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; et al. Genome-Wide SNP Analysis of Hybrid Clariid Fish Reflects the Existence of Polygenic Sex-Determination in the Lineage. Front. Genet. 2022, 13, 789573. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, G.; Shao, C.; Huang, Q.; Liu, G.; Zhang, P.; Song, W.; An, N.; Chalopin, D.; Volff, J.-N.; et al. Whole-Genome Sequence of a Flatfish Provides Insights into ZW Sex Chromosome Evolution and Adaptation to a Benthic Lifestyle. Nat. Genet. 2014, 46, 253–260. [Google Scholar] [CrossRef]
- Li, Y.-L.; Xing, T.-F.; Liu, J.-X. Genome-Wide Association Analyses Based on Whole-Genome Sequencing of Protosalanx Hyalocranius Provide Insights into Sex Determination of Salangid Fishes. Mol. Ecol. Resour. 2020, 20, 1038–1049. [Google Scholar] [CrossRef]
- de Oliveira, E.A.; Sember, A.; Bertollo, L.A.C.; Yano, C.F.; Ezaz, T.; Moreira-Filho, O.; Hatanaka, T.; Trifonov, V.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Tracking the Evolutionary Pathway of Sex Chromosomes among Fishes: Characterizing the Unique XX/XY1Y2 System in Hoplias malabaricus (Teleostei, Characiformes). Chromosoma 2018, 127, 115–128. [Google Scholar] [CrossRef]
- Li, M.; Zhang, R.; Fan, G.; Xu, W.; Zhou, Q.; Wang, L.; Li, W.; Pang, Z.; Yu, M.; Liu, Q.; et al. Reconstruction of the Origin of a Neo-Y Sex Chromosome and Its Evolution in the Spotted Knifejaw, Oplegnathus Punctatus. Mol. Biol. Evol. 2021, 38, 2615–2626. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Zou, Y.; Xiao, S.; Li, W.; Han, Z.; Han, F.; Xiao, J.; Liu, F.; Wang, Z. Chromosome Assembly of Collichthys lucidus, a Fish of Sciaenidae with a Multiple Sex Chromosome System. Sci. Data 2019, 6, 132. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zou, Y.; Xiao, S.; Chen, J.; Wang, Z.; Wang, Y.; Jie, X.; Cai, M. Development of a PCR-Based Genetic Sex Identification Method in Spinyhead Croaker (Collichthys lucidus). Aquaculture 2020, 522, 735130. [Google Scholar] [CrossRef]
- Araya-Jaime, C.; Mateussi, N.T.B.; Utsunomia, R.; Costa-Silva, G.J.; Oliveira, C.; Foresti, F. ZZ/Z0: The New System of Sex Chromosomes in Eigenmannia Aff. Trilineata (Teleostei: Gymnotiformes: Sternopygidae) Characterized by Molecular Cytogenetics and DNA Barcoding. Zebrafish 2017, 14, 464–470. [Google Scholar] [CrossRef]
- Mei, J.; Gui, J.-F. Genetic Basis and Biotechnological Manipulation of Sexual Dimorphism and Sex Determination in Fish. Sci. China Life Sci. 2015, 58, 124–136. [Google Scholar] [CrossRef]
- Liu, H.; Pang, M.; Yu, X.; Zhou, Y.; Tong, J.; Fu, B. Sex-Specific Markers Developed by next-Generation Sequencing Confirmed an XX/XY Sex Determination System in Bighead Carp (Hypophthalmichthys nobilis) and Silver Carp (Hypophthalmichthys molitrix). DNA Res. 2018, 25, 257–264. [Google Scholar] [CrossRef]
- Mu, J.; Hu, W.; Chen, R.; Yang, Y.; Li, H.; Li, W.; Yin, X.; Xu, D. Production of Neomale and Neofemale Large Yellow Croaker (Larimichthys Crocea) and Establishment of All-Female Populations. Aquaculture 2024, 590, 741010. [Google Scholar] [CrossRef]
- Niu, J.-S.; Wang, T.; Li, Z.; Wang, Z.-W.; Ding, M.; Wang, M.-T.; Lian, Z.-Q.; Mei, J.; Wang, Y.; Zhou, L.; et al. Efficient Breeding and Growth Advantage of All-Male Population in Lanzhou Catfish (Silurus Lanzhouensis). Aquaculture 2024, 578, 740023. [Google Scholar] [CrossRef]
- Zhao, J.; Ou, M.; Wang, Y.; Liu, H.; Luo, Q.; Zhu, X.; Chen, B.; Chen, K. Breeding of YY Super-Male of Blotched Snakehead (Channa maculata) and Production of All-Male Hybrid (Channa Argus ♀ × C. Maculata ♂). Aquaculture 2021, 538, 736450. [Google Scholar] [CrossRef]
- Ortega-Recalde, O.; Goikoetxea, A.; Hore, T.A.; Todd, E.V.; Gemmell, N.J. The Genetics and Epigenetics of Sex Change in Fish. Annu. Rev. Anim. Biosci. 2020, 8, 47–69. [Google Scholar] [CrossRef]
- Balogh, R.E.; Csorbai, B.; Guti, C.; Keszte, S.; Urbányi, B.; Orbán, L.; Kovács, B. Validation of a Male-Specific DNA Marker Confirms XX/XY-Type Sex Determination in Several Hungarian Strains of African Catfish (Clarias Gariepinus). Theriogenology 2023, 205, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Huang, W.; Peng, S.; Zhou, J.; Zhan, H.; Gui, L.; Li, W.; Li, Q. Screening and Characterization of Sex-Specific Markers by NGS Sequencing in Spinibarbus hollandi with Implication of XY Sex Determination System. Aquaculture 2023, 565, 739147. [Google Scholar] [CrossRef]
- Liu, H.; Xia, W.; Li, B.; Liu, L.; Wang, Y.; Luo, Q.; Ou, M.; Zhu, X.; Chen, K.; Zhao, J. Sex-Specific Markers Developed by 2b-RAD and Genome Sequencing Reveal an XX/XY Sex-Determination System in Mud Carp (Cirrhinus molitorella). Aquaculture 2023, 565, 739131. [Google Scholar] [CrossRef]
- Peng, Y.-X.; Huang, Y.-Q.; Zhong, J.; Jiang, Z.-T.; Fan, S.; Shi, H.-J.; Chen, H.-P.; Deng, S.-P.; Li, G.-L.; Jiang, D.-N. Identification of Sex-Linked Marker and Candidate Sex Determination Gene in Ornamental Fish, African Scat (Scatophagus Tetracanthus). Aquaculture 2023, 563, 739023. [Google Scholar] [CrossRef]
- Degani, G. Russian Sturgeon Acclimatization in Northern Israel: Nutrition, Reproduction, and Growth. Mich. J. Anim. Sci. Technol. 2023, 1, 1–36. [Google Scholar]
- Colihueque, N.; Parraguez, M. Assessing the Effectiveness of Sex-Linked Molecular Markers to Identify Neomale Breeders for the Production of All-Female Progenies of Rainbow Trout. Mar. Biotechnol. 2024, 26, 199–204. [Google Scholar] [CrossRef]
- Tao, W.; Zhu, X.; Cao, J.; Xiao, H.; Dong, J.; Kocher, T.D.; Lu, M.; Wang, D. Screening and Characterization of Sex-Linked DNA Markers in Mozambique Tilapia (Oreochromis mossambicus). Aquaculture 2022, 557, 738331. [Google Scholar] [CrossRef]
- Beardmore, J.A.; Mair, G.C.; Lewis, R.I. Monosex Male Production in Finfish as Exemplified by Tilapia: Applications, Problems, and Prospects. In Reproductive Biotechnology in Finfish Aquaculture; Lee, C.-S., Donaldson, E.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 283–301. ISBN 978-0-444-50913-0. [Google Scholar]
- Simco, B.A.; Goudie, C.A.; Klar, G.T.; Parker, N.C.; Davis, K.B. Influence of Sex on Growth of Channel Catfish. Trans. Am. Fish. Soc. 1989, 118, 427–434. [Google Scholar] [CrossRef]
- Liu, H.; Guan, B.; Xu, J.; Hou, C.; Tian, H.; Chen, H. Genetic Manipulation of Sex Ratio for the Large-Scale Breeding of YY Super-Male and XY All-Male Yellow Catfish (Pelteobagrus fulvidraco (Richardson)). Mar Biotechnol 2013, 15, 321–328. [Google Scholar] [CrossRef]
- Wang, D.-W.; Li, Y.; Li, Z.-Q. Identification of a Male-Specific Amplified Fragment Length Polymorphism (AFLP) and a Sequence Characterized Amplified Region (SCAR) Marker in Eucommia ulmoides Oliv. Int. J. Mol. Sci. 2011, 12, 857–864. [Google Scholar] [CrossRef]
- Wang, L.; Sun, F.; Wan, Z.Y.; Ye, B.; Wen, Y.; Liu, H.; Yang, Z.; Pang, H.; Meng, Z.; Fan, B.; et al. Genomic Basis of Striking Fin Shapes and Colors in the Fighting Fish. Mol. Biol. Evol. 2021, 38, 3383–3396. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-L.; Li, J.; Deng, S.-P.; Tian, Y.-S.; Wang, Q.-Y.; Zhuang, Z.-M.; Sha, Z.-X.; Xu, J.-Y. Isolation of Female-Specific AFLP Markers and Molecular Identification of Genetic Sex in Half-Smooth Tongue Sole (Cynoglossus semilaevis). Mar. Biotechnol. 2007, 9, 273–280. [Google Scholar] [CrossRef]
- Bye, V.J.; Lincoln, R.F. Commercial Methods for the Control of Sexual Maturation in Rainbow Trout (Salmo gairdneri R.). Aquaculture 1986, 57, 299–309. [Google Scholar] [CrossRef]
- Saillant, E.; Fostier, A.; Menu, B.; Haffray, P.; Chatain, B. Sexual Growth Dimorphism in Sea Bass Dicentrarchus Labrax. Aquaculture 2001, 202, 371–387. [Google Scholar] [CrossRef]
- Beullens, K.; Eding, E.H.; Ollevier, F.; Komen, J.; Richter, C.J.J. Sex Differentiation, Changes in Length, Weight and Eye Size before and after Metamorphosis of European Eel (Anguilla anguilla L.) Maintained in Captivity. Aquaculture 1997, 153, 151–162. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, N.; Peng, K.; He, X.; Chen, C.X.; Liu, H.; Liu, K.; Jia, L.; Bao, B. A Combination of Genome-Wide Association Study Screening and SNaPshot for Detecting Sex-Related SNPs and Genes in Cynoglossus Semilaevis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 35, 100711. [Google Scholar] [CrossRef]
- Martínez, P.; Viñas, A.M.; Sánchez, L.; Díaz, N.; Ribas, L.; Piferrer, F. Genetic Architecture of Sex Determination in Fish: Applications to Sex Ratio Control in Aquaculture. Front. Genet. 2014, 5, 340. [Google Scholar] [CrossRef]
- Zheng, S.; Wang, X.; Zhang, S.; Long, J.; Tao, W.; Li, M.; Wang, D. Screening and Characterization of Sex-Linked DNA Markers and Marker-Assisted Selection in the Southern Catfish (Silurus meridionalis). Aquaculture 2020, 517, 734783. [Google Scholar] [CrossRef]
- You, X.; Shan, X.; Shi, Q. Research Advances in the Genomics and Applications for Molecular Breeding of Aquaculture Animals. Aquaculture 2020, 526, 735357. [Google Scholar] [CrossRef]
- Peralta, D.M.; Túnez, J.I.; Cruz, U.E.R.; Ceballos, S.G. A Rapid Approach for Sex Assignment by RAD-Seq Using a Reference Genome. PLoS ONE 2024, 19, e0297987. [Google Scholar] [CrossRef]
- Teal, C.N.; Coykendall, D.K.; Campbell, M.R.; Eardley, D.L.; Delomas, T.A.; Shira, J.T.; Schill, D.J.; Bonar, S.A.; Culver, M. Sex-Specific Markers Undetected in Green Sunfish Lepomis Cyanellus Using Restriction-Site Associated DNA Sequencing. J. Fish Biol. 2022, 100, 1528–1540. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Liu, H.; Cheng, L.; Pan, Z.; Chang, G.; Wu, N.; Wang, H.; Ding, H.; Fang, Y.; Wang, L. Identification of Sex-Specific Sequences through 2b-RAD Sequencing in Pseudobagrus ussuriensis. Aquaculture 2021, 539, 736639. [Google Scholar] [CrossRef]
- Huang, W.; Lai, J.; Liang, W.; Ye, S.; Li, J.; Zhou, J.; Zhang, Y.; Peng, S.; Zhan, H.; Zheng, P.; et al. Identification of Sex-Specific DNA Markers in the Army Fish (Spinibarbus hollandi) by Whole Genome Re-Sequencing Method. Aquaculture 2024, 583, 740605. [Google Scholar] [CrossRef]
- Wen, M.; Wang, S.; Zhu, C.; Zhang, Y.; Liu, Z.; Wu, C.; Wang, S.; Wang, Y.; Ren, L.; Tao, M.; et al. Identification of Sex Locus and a Male-Specific Marker in Blunt-Snout Bream (Megalobrama amblycephala) Using a Whole Genome Resequencing Method. Aquaculture 2024, 582, 740559. [Google Scholar] [CrossRef]
- Wen, M.; Zhang, Y.; Wang, S.; Hu, F.; Tang, C.; Li, Q.; Qin, Q.; Tao, M.; Zhang, C.; Zhao, R.; et al. Sex Locus and Sex Markers Identification Using Whole Genome Pool-Sequencing Approach in the Largemouth Bass (Micropterus salmoides L.). Aquaculture 2022, 559, 738375. [Google Scholar] [CrossRef]
- Luo, H.; Xiao, J.; Jiang, Y.; Ke, Y.; Ke, C.; Cai, M. Mapping and Marker Identification for Sex-Determining in the Pacific Abalone, Haliotis Discus Hannai Ino. Aquaculture 2021, 530, 735810. [Google Scholar] [CrossRef]
- Wang, M.; Li, L.; Lin, H.; Zhou, Z.; Liu, B.; Zhong, J.; Pu, F.; Shi, Y.; Zhou, T.; Xu, P. Genome-Wide Association Study Identifies Genomic Loci of Sex Determination, Gonadal Weight and Gonadosomatic Index Traits in Takifugu Bimaculatus. Aquaculture 2022, 546, 737389. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, L.; Fan, Z.; Lu, B.; Chen, J.; Tan, D.; Jiang, D.; Tao, W.; Wang, D. Screening and Characterization of Sex-Linked DNA Markers and Marker-Assisted Selection in Blue Tilapia (Oreochromis aureus). Aquaculture 2021, 530, 735934. [Google Scholar] [CrossRef]
- Du, J.; Zhou, J.; Li, S.; Shao, J.; Jiang, P.; Dong, C.; Bai, J. A PCR-Based Method for Genetic Sex Identification and Evidence of the XX/XY Sex Determination System in Largemouth Bass (Micropterus salmoides L.). Aquaculture 2021, 545, 737220. [Google Scholar] [CrossRef]
- Martínez, P.; Robledo, D.; Taboada, X.; Blanco, A.; Moser, M.; Maroso, F.; Hermida, M.; Gómez-Tato, A.; Álvarez-Blázquez, B.; Cabaleiro, S.; et al. A Genome-Wide Association Study, Supported by a New Chromosome-Level Genome Assembly, Suggests Sox2 as a Main Driver of the Undifferentiatiated ZZ/ZW Sex Determination of Turbot (Scophthalmus maximus). Genomics 2021, 113, 1705–1718. [Google Scholar] [CrossRef]
- Liu, H.; Luo, Q.; Ou, M.; Zhu, X.; Zhao, J.; Chen, K. High-Density Genetic Linkage Map and QTL Fine Mapping of Growth and Sex in Snakehead (Channa argus). Aquaculture 2020, 519, 734760. [Google Scholar] [CrossRef]
- Ou, M.; Chen, K.-C.; Luo, Q.; Liu, H.-Y.; Wang, Y.-P.; Chen, B.-X.; Liang, X.-Q.; Zhao, J. Performance Evaluation of XY All-Male Hybrids Derived from XX Female Channa Argus and YY Super-Males Channa Maculate. Aquacult. Rep. 2021, 20, 100768. [Google Scholar] [CrossRef]
- Ou, M.; Yang, C.; Luo, Q.; Huang, R.; Zhang, A.-D.; Liao, L.-J.; Li, Y.-M.; He, L.-B.; Zhu, Z.-Y.; Chen, K.-C.; et al. An NGS-Based Approach for the Identification of Sex-Specific Markers in Snakehead (Channa argus). Oncotarget 2017, 8, 98733–98744. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wen, H.; Qi, X.; Li, C.; Sun, C.; Wang, L.; Zhu, M.; Jiang, T.; Zhang, X.; Li, Y. Comparative Study of Candidate Sex Determination Regions in Snakeheads (Channa Argus and C. Maculata) and Development of Novel Sex Markers. Aquaculture 2023, 575, 739771. [Google Scholar] [CrossRef]
- Wang, L.; Xie, N.; Shen, Y.; Ye, B.; Yue, G.H.; Feng, X. Constructing High-Density Genetic Maps and Developing Sexing Markers in Northern Snakehead (Channa argus). Mar. Biotechnol. 2019, 21, 348–358. [Google Scholar] [CrossRef]
- Fu, B.; Zhou, Y.; Liu, H.; Yu, X.; Tong, J. Updated Genome Assembly of Bighead Carp (Hypophthalmichthys nobilis) and Its Differences Between Male and Female on Genomic, Transcriptomic, and Methylation Level. Front. Genet. 2021, 12, 728177. [Google Scholar] [CrossRef]
- Li, S.; Xu, L.; Shi, Y.; Chen, J. Male-Specific Markers Developed by next-Generation Sequencing Confirmed an XX/XY Sex-Determination System in Farmed Ayu (Plecoglossus altivelis). Aquaculture 2021, 541, 736822. [Google Scholar] [CrossRef]
- Felip, A.; Young, W.P.; Wheeler, P.A.; Thorgaard, G.H. An AFLP-Based Approach for the Identification of Sex-Linked Markers in Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2005, 247, 35–43. [Google Scholar] [CrossRef]
- Chen, S.-L.; Ji, X.-S.; Shao, C.-W.; Li, W.-L.; Yang, J.-F.; Liang, Z.; Liao, X.-L.; Xu, G.-B.; Xu, Y.; Song, W.-T. Induction of Mitogynogenetic Diploids and Identification of WW Super-Female Using Sex-Specific SSR Markers in Half-Smooth Tongue Sole (Cynoglossus semilaevis). Mar. Biotechnol. 2012, 14, 120–128. [Google Scholar] [CrossRef]
- Liu, Z.J.; Cordes, J.F. DNA Marker Technologies and Their Applications in Aquaculture Genetics. Aquaculture 2004, 238, 1–37. [Google Scholar] [CrossRef]
- Dai, S.; Chen, M.; Zheng, S.; Su, J.; Wu, J.; Han, L.; Zhou, C.; Zou, Y.; Wang, D.; Li, M. Sex-Linked DNA Marker Screening and Characterization in Albino Northern Snakehead (Channa argus Var.) via Third-Generation Sequencing and Pool Resequencing. Aquaculture 2025, 594, 741449. [Google Scholar] [CrossRef]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY Is a Y-Specific DM-Domain Gene Required for Male Development in the Medaka Fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Xiao, S.; Xu, S.; Ye, K.; Lin, X.; Sun, S.; Wang, Z. Identification of a Male-Specific DNA Marker in the Large Yellow Croaker (Larimichthys crocea). Aquaculture 2017, 480, 116–122. [Google Scholar] [CrossRef]
- Mustapha, U.F.; Jiang, D.-N.; Liang, Z.-H.; Gu, H.-T.; Yang, W.; Chen, H.-P.; Deng, S.-P.; Wu, T.-L.; Tian, C.-X.; Zhu, C.-H.; et al. Male-Specific Dmrt1 Is a Candidate Sex Determination Gene in Spotted Scat (Scatophagus argus). Aquaculture 2018, 495, 351–358. [Google Scholar] [CrossRef]
- Han, C.; Zhu, Q.; Lu, H.; Wang, C.; Zhou, X.; Peng, C.; Tang, L.; Han, L.; Chen, J.; Li, S.; et al. Screening and Characterization of Sex-Specific Markers Developed by a Simple NGS Method in Mandarin Fish (Siniperca chuatsi). Aquaculture 2020, 527, 735495. [Google Scholar] [CrossRef]
- Li, M.; Xu, H.; Xu, W.; Zhou, Q.; Xu, X.; Zhu, Y.; Zheng, W.; Li, W.; Pang, Z.; Chen, S. Isolation of a Male-Specific Molecular Marker and Development of a Genetic Sex Identification Technique in Spotted Knifejaw (Oplegnathus punctatus). Mar. Biotechnol. 2020, 22, 467–474. [Google Scholar] [CrossRef]
- Han, C.; Zhou, X.; Lu, H.; Zhu, Q.; Han, L.; Li, S.; Lin, H.; Zhang, Y. A Simple PCR-Based Genetic Sex Identification Method in the Blotched Snakehead (Channa maculata) Developed by High-Throughput Sequencing. Aquaculture 2021, 538, 736579. [Google Scholar] [CrossRef]
- Song, W.; Xie, Y.; Sun, M.; Li, X.; Fitzpatrick, C.K.; Vaux, F.; O’Malley, K.G.; Zhang, Q.; Qi, J.; He, Y. A Duplicated Amh Is the Master Sex-Determining Gene for Sebastes Rockfish in the Northwest Pacific. Open Biol. 2021, 11, 210063. [Google Scholar] [CrossRef]
- Wei, L.; Yang, C.; Tao, W.; Wang, D. Genome-Wide Identification and Transcriptome-Based Expression Profiling of the Sox Gene Family in the Nile Tilapia (Oreochromis niloticus). Int. J. Mol. Sci. 2016, 17, 270. [Google Scholar] [CrossRef]
- Zhang, X.; Ruan, Z.; Sun, C.; Hu, C.; Huang, Y.; You, X.; Wang, X.; Xu, J.; Liu, H.; Liu, X.; et al. Genome-Wide Identification, Sequence Alignment, and Transcription of Five Sex-Related Genes in Largemouth Bass (Micropterus salmoides). FBL 2024, 29, 63. [Google Scholar] [CrossRef]
- Yang, J.; Hu, Y.; Han, J.; Xiao, K.; Liu, X.; Tan, C.; Zeng, Q.; Du, H. Genome-Wide Analysis of the Chinese Sturgeon Sox Gene Family: Identification, Characterisation and Expression Profiles of Different Tissues. J. Fish Biol. 2020, 96, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lin, D.; Zhang, Y.; Ma, G.; Zhang, W. A Homologue of Sox11 Predominantly Expressed in the Ovary of the Orange-Spotted Grouper Epinephelus Coioides. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 149, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhou, L.; Zhang, J.; Wang, Y.; Wang, Z.; Liu, X.; Cai, M. Genome-Wide Identification and Transcriptome-Based Expression Profiling of the Sox Gene Family in the Spinyhead Croaker (Collichthys lucidus). J. Fish Biol. 2022, 100, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Bertho, S.; Herpin, A.; Branthonne, A.; Jouanno, E.; Yano, A.; Nicol, B.; Muller, T.; Pannetier, M.; Pailhoux, E.; Miwa, M.; et al. The Unusual Rainbow Trout Sex Determination Gene Hijacked the Canonical Vertebrate Gonadal Differentiation Pathway. Proc. Natl. Acad. Sci. USA 2018, 115, 12781–12786. [Google Scholar] [CrossRef] [PubMed]
- Ayllon, F.; Solberg, M.F.; Besnier, F.; Fjelldal, P.G.; Hansen, T.J.; Wargelius, A.; Edvardsen, R.B.; Glover, K.A. Autosomal sdY Pseudogenes Explain Discordances Between Phenotypic Sex and DNA Marker for Sex Identification in Atlantic Salmon. Front. Genet. 2020, 11, 544207. [Google Scholar] [CrossRef]
- Bertho, S.; Herpin, A.; Schartl, M.; Guiguen, Y. Lessons from an Unusual Vertebrate Sex-Determining Gene. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200092. [Google Scholar] [CrossRef]
- Yano, A.; Nicol, B.; Jouanno, E.; Quillet, E.; Fostier, A.; Guyomard, R.; Guiguen, Y. The Sexually Dimorphic on the Y-Chromosome Gene (sdY) Is a Conserved Male-Specific Y-Chromosome Sequence in Many Salmonids. Evol. Appl. 2013, 6, 486–496. [Google Scholar] [CrossRef]
- Bao, L.; Tian, C.; Liu, S.; Zhang, Y.; Elaswad, A.; Yuan, Z.; Khalil, K.; Sun, F.; Yang, Y.; Zhou, T.; et al. The Y Chromosome Sequence of the Channel Catfish Suggests Novel Sex Determination Mechanisms in Teleost Fish. BMC Biol. 2019, 17, 6. [Google Scholar] [CrossRef]
- Xie, Q.-P.; Zhan, W.; Shi, J.-Z.; Liu, F.; Niu, B.-L.; He, X.; Liu, M.; Wang, J.; Liang, Q.-Q.; Xie, Y.; et al. Whole-Genome Assembly and Annotation for the Little Yellow Croaker (Larimichthys polyactis) Provide Insights into the Evolution of Hermaphroditism and Gonochorism. Mol. Ecol. Resour. 2023, 23, 632–658. [Google Scholar] [CrossRef]
- Adolfi, M.C.; Du, K.; Kneitz, S.; Cabau, C.; Zahm, M.; Klopp, C.; Feron, R.; Paixão, R.V.; Varela, E.S.; de Almeida, F.L.; et al. A Duplicated Copy of Id2b Is an Unusual Sex-Determining Candidate Gene on the Y Chromosome of Arapaima (Arapaima gigas). Sci. Rep. 2021, 11, 21544. [Google Scholar] [CrossRef]
- He, Z.; Yan, R.-G.; Shang, Q.-B.; Yang, Q.-E. Elevated Id2 Expression Causes Defective Meiosis and Spermatogenesis in Mice. Dev. Dyn. 2024, 253, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.; Zhou, Z.; Zhou, T.; Zhao, J.; Chen, L.; Lin, H.; Pu, F.; Ke, Q.; Bai, H.; Xu, P. Genome-Wide Association Study of Body Shape-Related Traits in Large Yellow Croaker (Larimichthys crocea). Mar. Biotechnol. 2020, 22, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows–Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Zhou, X.; Stephens, M. Genome-Wide Efficient Mixed-Model Analysis for Association Studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef]
- Zhang, Z.; Ersoz, E.; Lai, C.-Q.; Todhunter, R.J.; Tiwari, H.K.; Gore, M.A.; Bradbury, P.J.; Yu, J.; Arnett, D.K.; Ordovas, J.M.; et al. Mixed Linear Model Approach Adapted for Genome-Wide Association Studies. Nat. Genet. 2010, 42, 355–360. [Google Scholar] [CrossRef]
- Ai, C.H.; Zhu, Z.X.; Huang, D.D.; Yang, G.; De Liu, T.; Bai, Y.; Liang, X.Y.; Xiong, Y.Y.; Lin, Y.L.; Lin, H.R.; et al. Identification of SNPs and Candidate Genes Associated with Early Growth in Orange-Spotted Grouper (Epinephelus coioides) by a Genome-Wide Association Study. Aquaculture 2023, 565, 739129. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X.; et al. rMVP: A Memory-Efficient, Visualization-Enhanced, and Parallel-Accelerated Tool for Genome-Wide Association Study. Genom. Proteom. Bioinform. 2020, 19, 619–628. [Google Scholar] [CrossRef]
- Tanner, M.; Sharrard, M.; Rigby, A. Gene Polymorphisms and the Use of the Bonferroni Correction Factor: When and When Not to Apply? Arch. Dis. Child. 1997, 76, 385. [Google Scholar] [CrossRef][Green Version]
- Nei, M. Definition and Estimation of Fixation Indices. Evolution 1986, 40, 643–645. [Google Scholar] [CrossRef]
- Dong, S.-S.; He, W.-M.; Ji, J.-J.; Zhang, C.; Guo, Y.; Yang, T.-L. LDBlockShow: A Fast and Convenient Tool for Visualizing Linkage Disequilibrium and Haplotype Blocks Based on Variant Call Format Files. Brief. Bioinform. 2021, 22, bbaa227. [Google Scholar] [CrossRef]
- Ou, M.; Huang, R.; Yang, C.; Gui, B.; Luo, Q.; Zhao, J.; Li, Y.; Liao, L.; Zhu, Z.; Wang, Y.; et al. Chromosome-Level Genome Assemblies of Channa Argus and Channa Maculata and Comparative Analysis of Their Temperature Adaptability. GigaScience 2021, 10, giab070. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.-C.; Konaté, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or Normalized Counts? A Comparative Study of Quantification Measures for the Analysis of RNA-Seq Data from the NCI Patient-Derived Models Repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated Genome Annotation and Pathway Identification Using the KEGG Orthology (KO) as a Controlled Vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, J.; Liu, C.; Luo, S.; Gao, X.; Wu, Y.; Wang, J.; Wang, X.; Wu, X.; Shen, W.; et al. Genetics Responses to Hypoxia and Reoxygenation Stress in Larimichthys Crocea Revealed via Transcriptome Analysis and Weighted Gene Co-Expression Network. Animals 2021, 11, 3021. [Google Scholar] [CrossRef]
- Raivo Kolde Pheatmap: Pretty Heatmaps 2010, 1.0.12. Available online: https://cran.r-project.org/web/packages/pheatmap/pheatmap.pdf (accessed on 6 October 2024).
- Mao, H.; Chen, K.; Zhu, X.; Luo, Q.; Zhao, J.; Li, W.; Wu, X.; Xu, H. Identification of Suitable Reference Genes for Quantitative Real-Time PCR Normalization in Blotched Snakehead Channa Maculata. J. Fish Biol. 2017, 90, 2312–2322. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).