
Single-Cell and Spatial Multi-Omics Analysis Reveal That Targeting JAG1 in Epithelial Cells Reduces Periodontal Inflammation and Alveolar Bone Loss


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
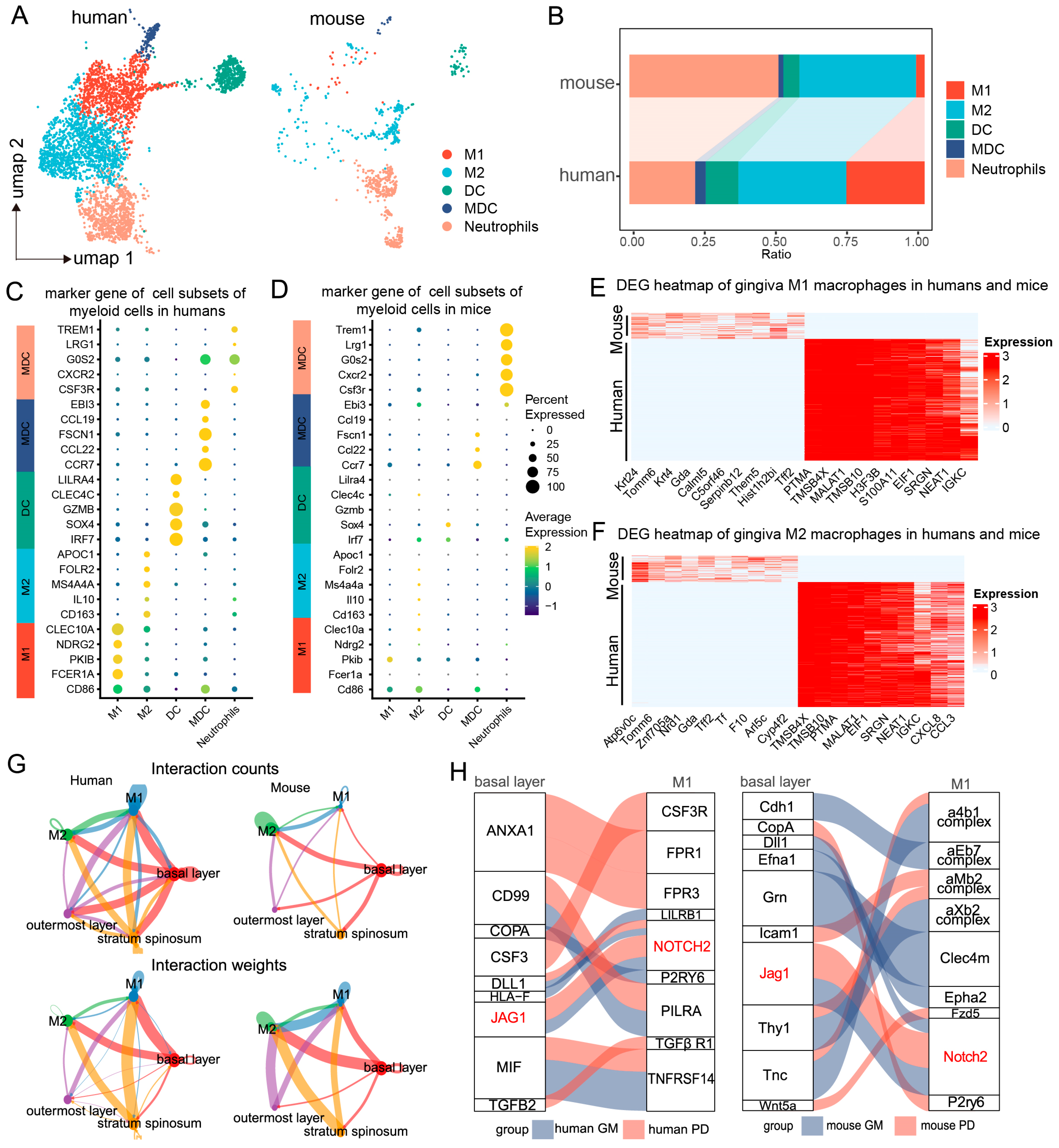
2.1. Single-Cell and Spatial Transcriptomic Analysis Reveals Close Communication Between Epithelial Cells and Macrophages in Periodontitis
2.2. Regulation of Macrophages by Basal Epithelial Cells
2.3. Analysis of Basal Epithelial Cell Activation Pathways and Downstream Genes During Periodontitis
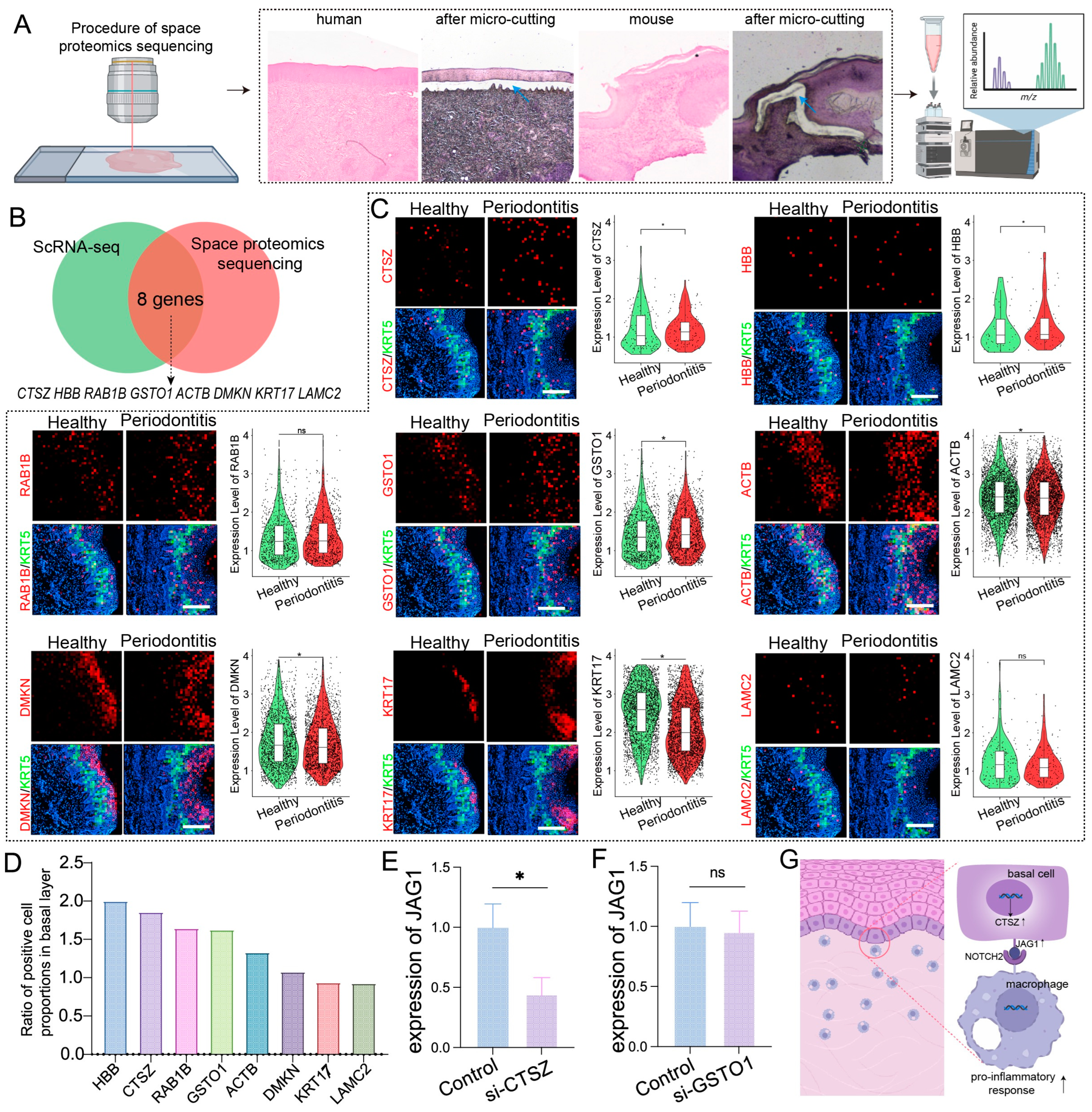
2.4. Spatial Proteomics and Spatial Transcriptomics Verify Genes and Proteins Associated with Basal Layer Inflammation in Periodontitis
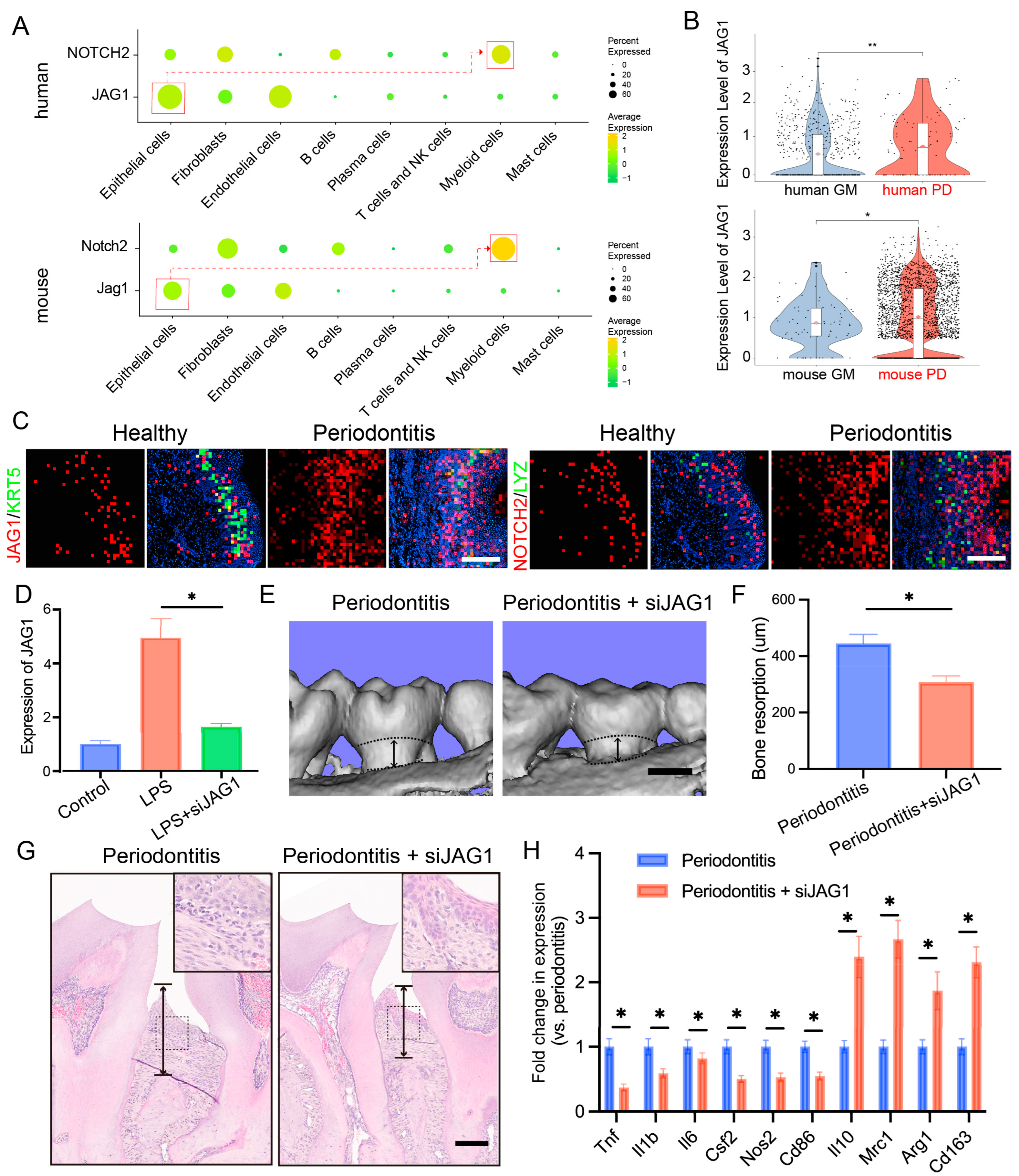
2.5. Targeting the JAG1–NOTCH2 Axis Reduces Periodontal Inflammation and Alveolar Bone Loss in Periodontitis
3. Discussion
4. Materials and Methods
4.1. Data Acquisition
4.2. Patient Recruitment and Tissue Collection
4.3. Spatial Proteomics of the Human Gingival Samples
4.4. Animal Experimentation
4.5. Tissue Collection and Processing for the Animal Study
4.6. Cell Culture
4.7. uCT Analysis
4.8. Data Preprocessing and Quality Control
4.9. Data Integration and Comparative Analysis
4.10. Cell-Cell Communication Analysis
4.11. Immunofluorescence Staining
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hajishengallis, G.; Chavakis, T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.W.; Greenwell-Wild, T.; Brenchley, L.; Dutzan, N.; Overmiller, A.; Sawaya, A.P.; Webb, S.; Martin, D.; Hajishengallis, G.; Divaris, K.; et al. Human oral mucosa cell atlas reveals a stromal-neutrophil axis regulating tissue immunity. Cell 2021, 184, 4090–4104.e15. [Google Scholar] [CrossRef] [PubMed]
- Manresa, C.; Sanz-Miralles, E.C.; Twigg, J.; Bravo, M. Supportive periodontal therapy (SPT) for maintaining the dentition in adults treated for periodontitis. Cochrane Database Syst. Rev. 2018, 2018, Cd009376. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Kuang, S.; Zhang, Y.; Chen, J.; Wang, S.; Xu, C.; Huang, Y.; Zhang, M.; Huang, S.; Wang, J.; et al. Restoring periodontal tissue homoeostasis prevents cognitive decline by reducing the number of Serpina3n(high) astrocytes in the hippocampus. Innovation 2024, 5, 100547. [Google Scholar] [PubMed]
- Gaffen, S.L.; Moutsopoulos, N.M. Regulation of host-microbe interactions at oral mucosal barriers by type 17 immunity. Sci. Immunol. 2020, 5, eaau4594. [Google Scholar] [CrossRef]
- Silva, L.M.; Doyle, A.D.; Greenwell-Wild, T.; Dutzan, N.; Tran, C.L.; Abusleme, L.; Juang, L.J.; Leung, J.; Chun, E.M.; Lum, A.G.; et al. Fibrin is a critical regulator of neutrophil effector function at the oral mucosal barrier. Science 2021, 374, eabl5450. [Google Scholar] [CrossRef]
- Shen, Z.; Zhang, R.; Huang, Y.; Chen, J.; Yu, M.; Li, C.; Zhang, Y.; Chen, L.; Huang, X.; Yang, J.; et al. The spatial transcriptomic landscape of human gingiva in health and periodontitis. Sci. China Life Sci. 2023, 67, 720–732. [Google Scholar] [CrossRef]
- Huang, F.; Song, Y.; Chen, W.; Liu, Q.; Wang, Q.; Liu, W.; Wang, X.; Wang, W. Effects of Candida albicans infection on defense effector secretion by human oral mucosal epithelial cells. Arch. Oral Biol. 2019, 103, 55–61. [Google Scholar] [CrossRef]
- Dutzan, N.; Abusleme, L.; Bridgeman, H.; Greenwell-Wild, T.; Zangerle-Murray, T.; Fife, M.E.; Bouladoux, N.; Linley, H.; Brenchley, L.; Wemyss, K.; et al. On-going Mechanical Damage from Mastication Drives Homeostatic Th17 Cell Responses at the Oral Barrier. Immunity 2017, 46, 133–147. [Google Scholar] [CrossRef]
- Bai, L.; Chen, B.-Y.; Liu, Y.; Zhang, W.-C.; Duan, S.-Z. A Mouse Periodontitis Model with Humanized Oral Bacterial Community. Front. Cell. Infect. Microbiol. 2022, 12, 842845. [Google Scholar] [CrossRef]
- Marchesan, J.; Girnary, M.S.; Jing, L.; Miao, M.Z.; Zhang, S.; Sun, L.; Morelli, T.; Schoenfisch, M.H.; Inohara, N.; Offenbacher, S.; et al. An experimental murine model to study periodontitis. Nat. Protoc. 2018, 13, 2247–2267. [Google Scholar] [CrossRef] [PubMed]
- Hackert, N.S.; Radtke, F.A.; Exner, T.; Lorenz, H.-M.; Müller-Tidow, C.; Nigrovic, P.A.; Wabnitz, G.; Grieshaber-Bouyer, R. Human and mouse neutrophils share core transcriptional programs in both homeostatic and inflamed contexts. Nat. Commun. 2023, 14, 8133. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Lin, Y.; Nery, J.R.; Urich, M.A.; Breschi, A.; Davis, C.A.; Dobin, A.; Zaleski, C.; Beer, M.A.; Chapman, W.C.; et al. Comparison of the transcriptional landscapes between human and mouse tissues. Proc. Natl. Acad. Sci. USA 2014, 111, 17224–17229. [Google Scholar] [CrossRef] [PubMed]
- Jiao, A.; Zhang, C.; Wang, X.; Sun, L.; Liu, H.; Su, Y.; Lei, L.; Li, W.; Ding, R.; Ding, C.; et al. Single-cell sequencing reveals the evolution of immune molecules across multiple vertebrate species. J. Adv. Res. 2024, 55, 73–87. [Google Scholar] [CrossRef]
- Li, J.; Ma, J.; Zhang, Q.; Gong, H.; Gao, D.; Wang, Y.; Li, B.; Li, X.; Zheng, H.; Wu, Z.; et al. Spatially resolved proteomic map shows that extracellular matrix regulates epidermal growth. Nat. Commun. 2022, 13, 4012. [Google Scholar] [CrossRef]
- Kim, T.S.; Ikeuchi, T.; Theofilou, V.I.; Williams, D.W.; Greenwell-Wild, T.; June, A.; Adade, E.E.; Li, L.; Abusleme, L.; Dutzan, N.; et al. Epithelial-derived interleukin-23 promotes oral mucosal immunopathology. Immunity 2024, 57, 859–875.e11. [Google Scholar] [CrossRef]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef]
- Kong, L.; Pokatayev, V.; Lefkovith, A.; Carter, G.T.; Creasey, E.A.; Krishna, C.; Subramanian, S.; Kochar, B.; Ashenberg, O.; Lau, H.; et al. The landscape of immune dysregulation in Crohn’s disease revealed through single-cell transcriptomic profiling in the ileum and colon. Immunity 2023, 56, 444–458.e5. [Google Scholar] [CrossRef]
- Fawkner-Corbett, D.; Antanaviciute, A.; Parikh, K.; Jagielowicz, M.; Gerós, A.S.; Gupta, T.; Ashley, N.; Khamis, D.; Fowler, D.; Morrissey, E.; et al. Spatiotemporal analysis of human intestinal development at single-cell resolution. Cell 2021, 184, 810–826.e23. [Google Scholar] [CrossRef]
- Bosshardt, D.D. The periodontal pocket: Pathogenesis, histopathology and consequences. Periodontology 2000 2018, 76, 43–50. [Google Scholar] [CrossRef]
- Kayama, H.; Okumura, R.; Takeda, K. Interaction Between the Microbiota, Epithelia, and Immune Cells in the Intestine. Annu. Rev. Immunol. 2020, 38, 23–48. [Google Scholar] [CrossRef] [PubMed]
- Loomba, S.; Straehle, J.; Gangadharan, V.; Heike, N.; Khalifa, A.; Motta, A.; Ju, N.; Sievers, M.; Gempt, J.; Meyer, H.S.; et al. Connectomic comparison of mouse and human cortex. Science 2022, 377, eabo0924. [Google Scholar] [CrossRef] [PubMed]
- Shami, A.N.; Zheng, X.; Munyoki, S.K.; Ma, Q.; Manske, G.L.; Green, C.D.; Sukhwani, M.; Orwig, K.E.; Li, J.Z.; Hammoud, S.S. Single-Cell RNA Sequencing of Human, Macaque, and Mouse Testes Uncovers Conserved and Divergent Features of Mammalian Spermatogenesis. Dev. Cell 2020, 54, 529–547.e12. [Google Scholar] [CrossRef]
- Mao, C.; Ma, Z.; Jia, Y.; Li, W.; Xie, N.; Zhao, G.; Ma, B.; Yu, F.; Sun, J.; Zhou, Y.; et al. Nidogen-2 Maintains the Contractile Phenotype of Vascular Smooth Muscle Cells and Prevents Neointima Formation via Bridging Jagged1-Notch3 Signaling. Circulation 2021, 144, 1244–1261. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhu, C.; Wang, X.; Kim, K.; Bartolome, A.; Dongiovanni, P.; Yates, K.P.; Valenti, L.; Carrer, M.; Sadowski, T.; et al. Hepatocyte TLR4 triggers inter-hepatocyte Jagged1/Notch signaling to determine NASH-induced fibrosis. Sci. Transl. Med. 2021, 13, eabe1692. [Google Scholar] [CrossRef]
- Siouti, E.; Salagianni, M.; Manioudaki, M.; Pavlos, E.; Klinakis, A.; Galani, I.; Andreakos, E. Notch signaling in adipose tissue macrophages prevents diet-induced inflammation and metabolic dysregulation. Eur. J. Immunol. 2024, 54, e2350669. [Google Scholar] [CrossRef]
- López-López, S.; de Ávila, M.J.R.; de León, N.C.H.; Ruiz-Marcos, F.; Baladrón, V.; Nueda, M.L.; Laborda, J.; García-Ramírez, J.J.; Monsalve, E.M.; Díaz-Guerra, M.J.M. NOTCH4 Exhibits Anti-Inflammatory Activity in Activated Macrophages by Interfering with Interferon-γ and TLR4 Signaling. Front. Immunol. 2021, 12, 734966. [Google Scholar] [CrossRef]
- Ayuthaya, B.I.N.; Lertpimonchai, A.; Samaranayake, L.; Vathesatogkit, P.; Thienpramuk, L.; Wisitrasameewong, W.; Tamsailom, S. The Potential Effect of Periodontal Disease on the Development of Metabolic Syndrome: A 10-Year Observational Study in a Thai Adult Cohort. J. Clin. Periodontol. 2024. [Google Scholar] [CrossRef]
- Pirih, F.Q.; Monajemzadeh, S.; Singh, N.; Sinacola, R.S.; Shin, J.M.; Chen, T.; Fenno, J.C.; Kamarajan, P.; Rickard, A.H.; Travan, S.; et al. Association between metabolic syndrome and periodontitis: The role of lipids, inflammatory cytokines, altered host response, and the microbiome. Periodontology 2000 2021, 87, 50–75. [Google Scholar] [CrossRef]
- Groeger, S.; Meyle, J. Oral Mucosal Epithelial Cells. Front. Immunol. 2019, 10, 208. [Google Scholar] [CrossRef]
- Moutsopoulos, N.M.; Konkel, J.E. Tissue-Specific Immunity at the Oral Mucosal Barrier. Trends Immunol. 2018, 39, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.; García, M.P.; Polanco, A.F.; González-Osuna, L.; Sierra-Cristancho, A.; Melgar-Rodríguez, S.; Cafferata, E.A.; Vernal, R. Humanized Mouse Models for the Study of Periodontitis: An Opportunity to Elucidate Unresolved Aspects of Its Immunopathogenesis and Analyze New Immunotherapeutic Strategies. Front. Immunol. 2021, 12, 663328. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Niimi, H.; Ohsugi, Y.; Tsuchiya, Y.; Shimohira, T.; Komatsu, K.; Liu, A.; Shiba, T.; Aoki, A.; Iwata, T.; et al. Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease. Int. J. Mol. Sci. 2021, 22, 8900. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuang, S.; Yang, J.; Shen, Z.; Xia, J.; Lin, Z. Single-Cell and Spatial Multi-Omics Analysis Reveal That Targeting JAG1 in Epithelial Cells Reduces Periodontal Inflammation and Alveolar Bone Loss. Int. J. Mol. Sci. 2024, 25, 13255. https://doi.org/10.3390/ijms252413255
Kuang S, Yang J, Shen Z, Xia J, Lin Z. Single-Cell and Spatial Multi-Omics Analysis Reveal That Targeting JAG1 in Epithelial Cells Reduces Periodontal Inflammation and Alveolar Bone Loss. International Journal of Molecular Sciences. 2024; 25(24):13255. https://doi.org/10.3390/ijms252413255
Chicago/Turabian StyleKuang, Shuhong, Jiayu Yang, Zongshan Shen, Juan Xia, and Zhengmei Lin. 2024. "Single-Cell and Spatial Multi-Omics Analysis Reveal That Targeting JAG1 in Epithelial Cells Reduces Periodontal Inflammation and Alveolar Bone Loss" International Journal of Molecular Sciences 25, no. 24: 13255. https://doi.org/10.3390/ijms252413255
APA StyleKuang, S., Yang, J., Shen, Z., Xia, J., & Lin, Z. (2024). Single-Cell and Spatial Multi-Omics Analysis Reveal That Targeting JAG1 in Epithelial Cells Reduces Periodontal Inflammation and Alveolar Bone Loss. International Journal of Molecular Sciences, 25(24), 13255. https://doi.org/10.3390/ijms252413255