
HR-MAS NMR Metabolomics Profile of Vero Cells under the Influence of Virus Infection and nsP2 Inhibitor: A Chikungunya Case Study

 , ,
, ,  , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Results
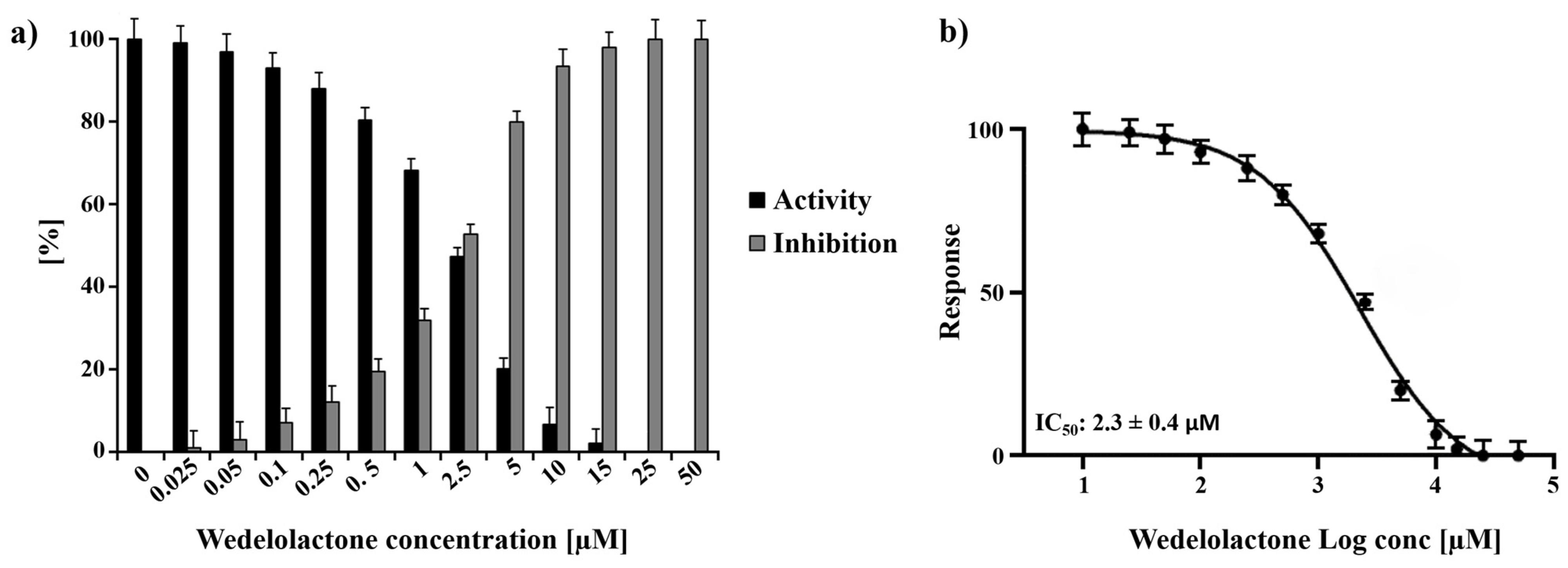
2.1. In Vitro Inhibition Assay of the CHIKV ns2 Protease (nsP2) by Wedelolactone
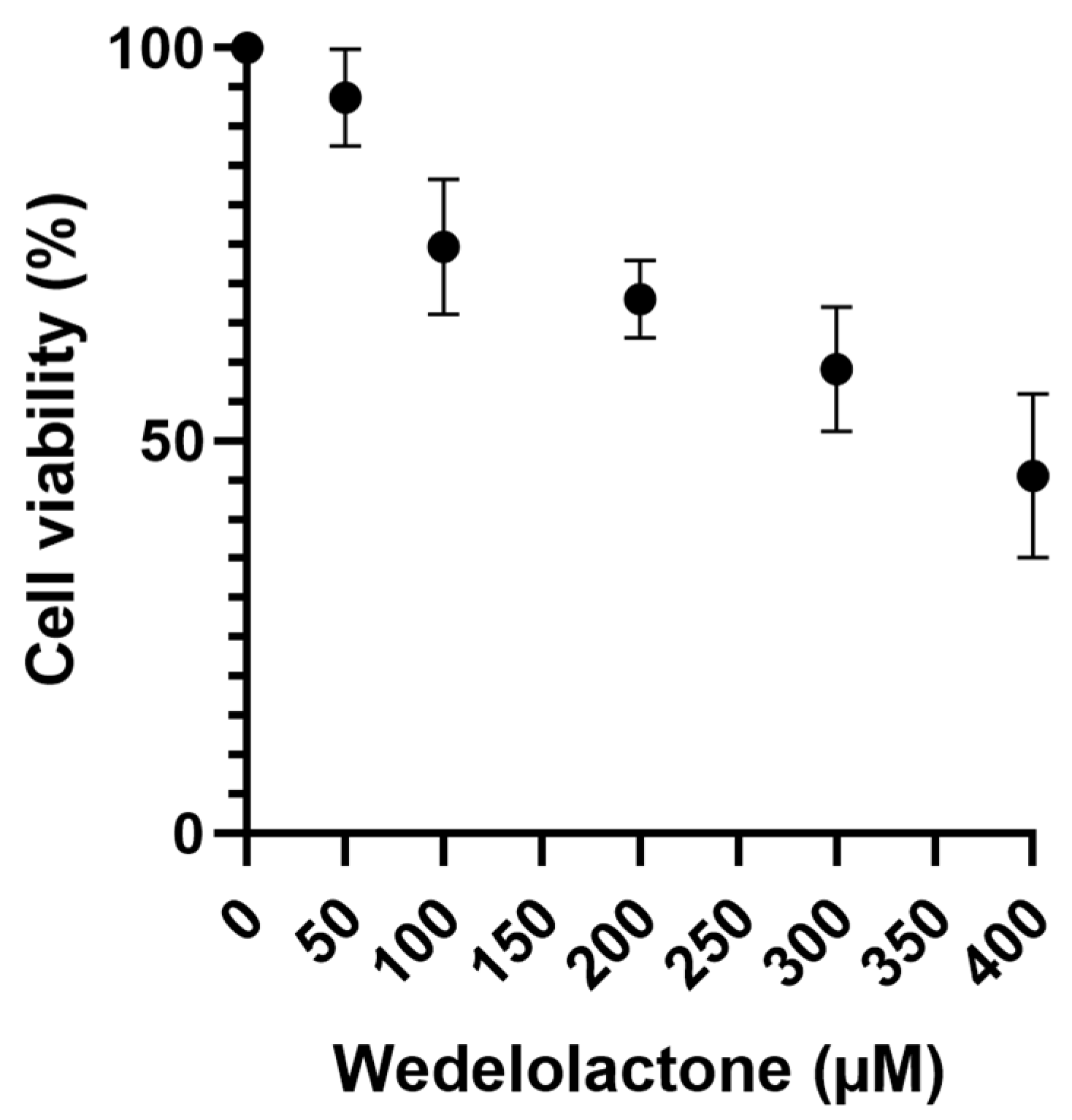
2.2. Assessment of Wedelolactone Cytotoxicity In Vero E6 Cells
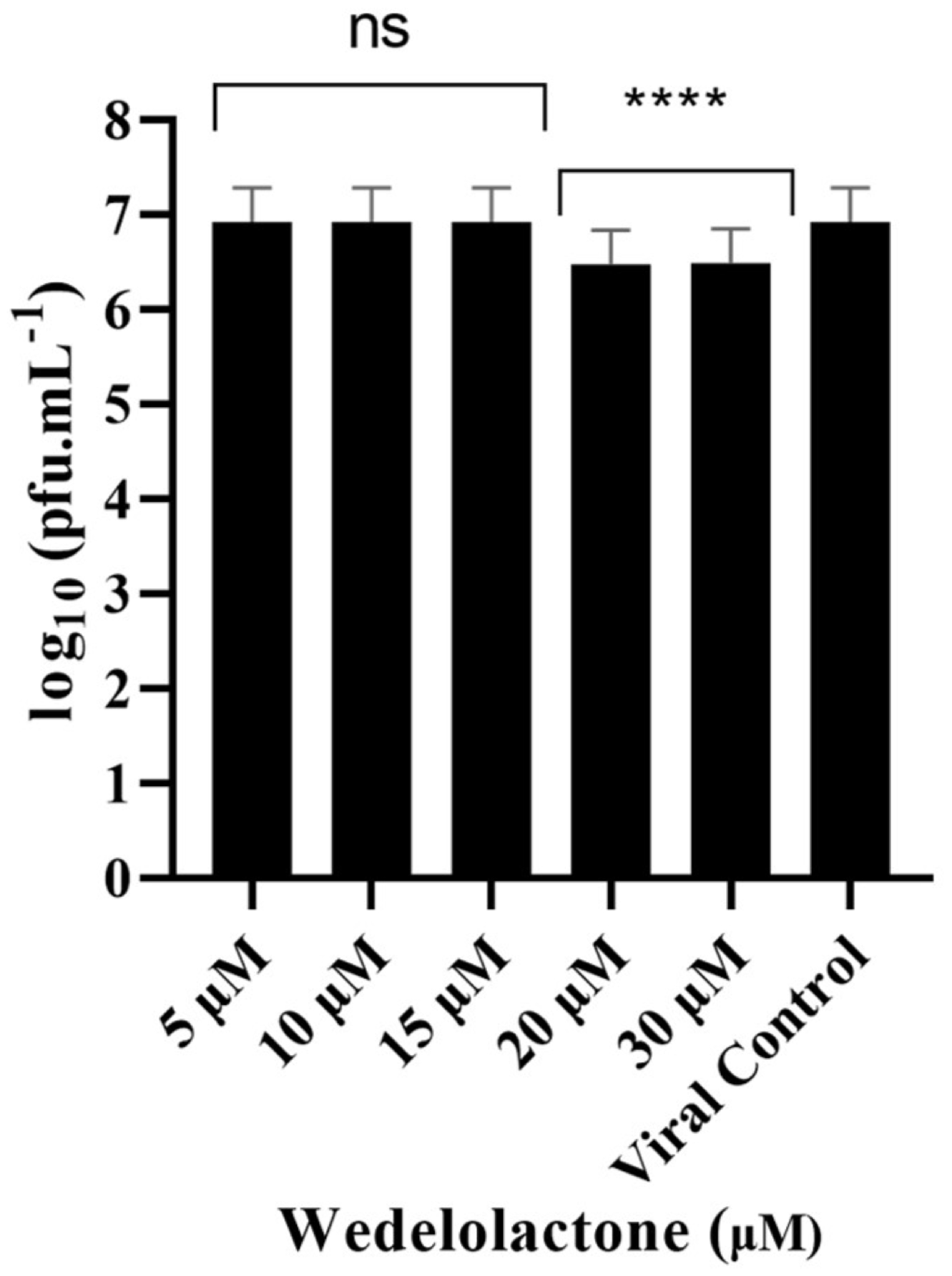
2.3. Effects of Wedelolactone on CHIKV-Infected Cells
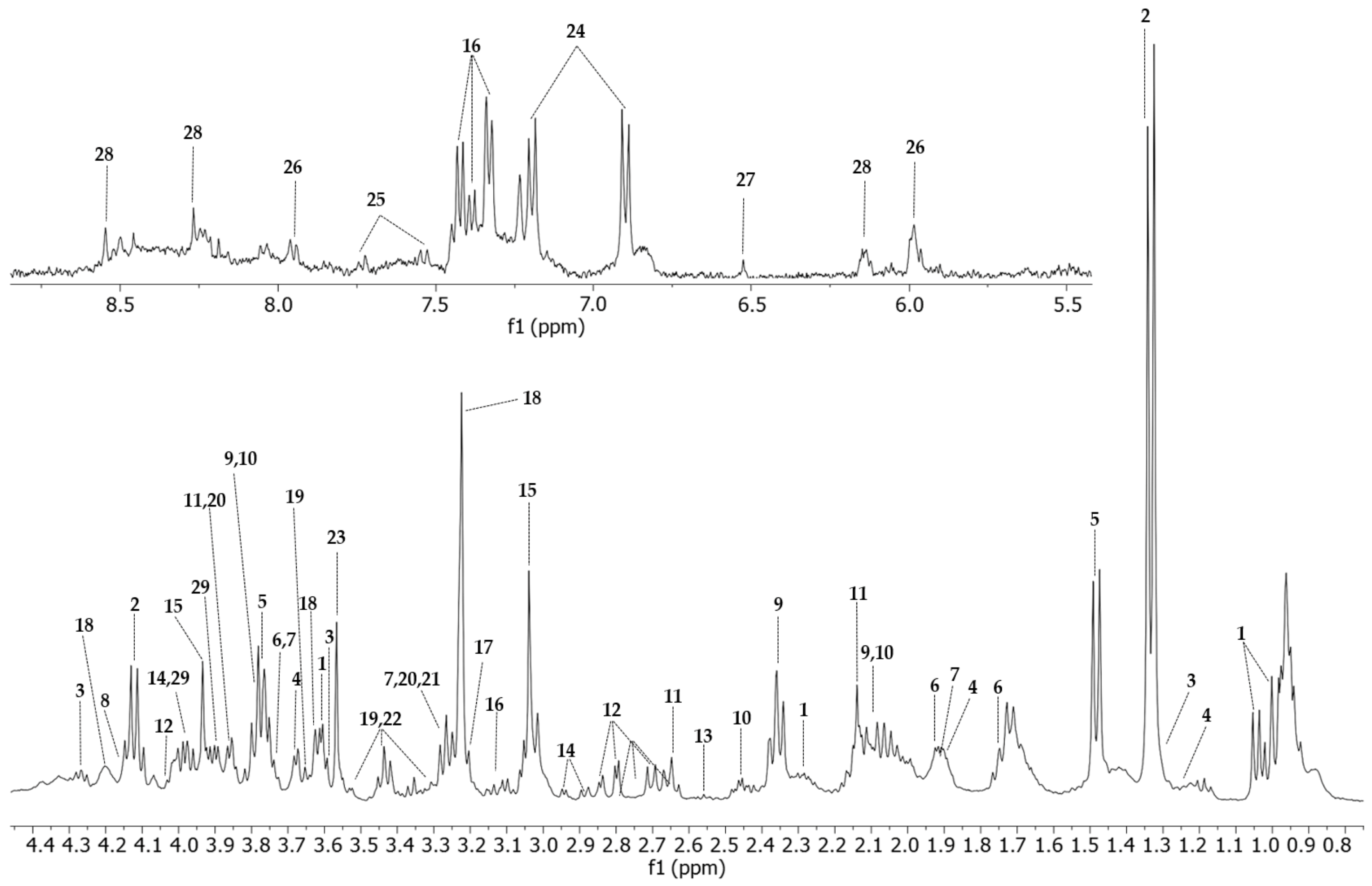
2.4. HR-MAS NMR Spectroscopy Applied to the Analysis of Vero E6 Cells
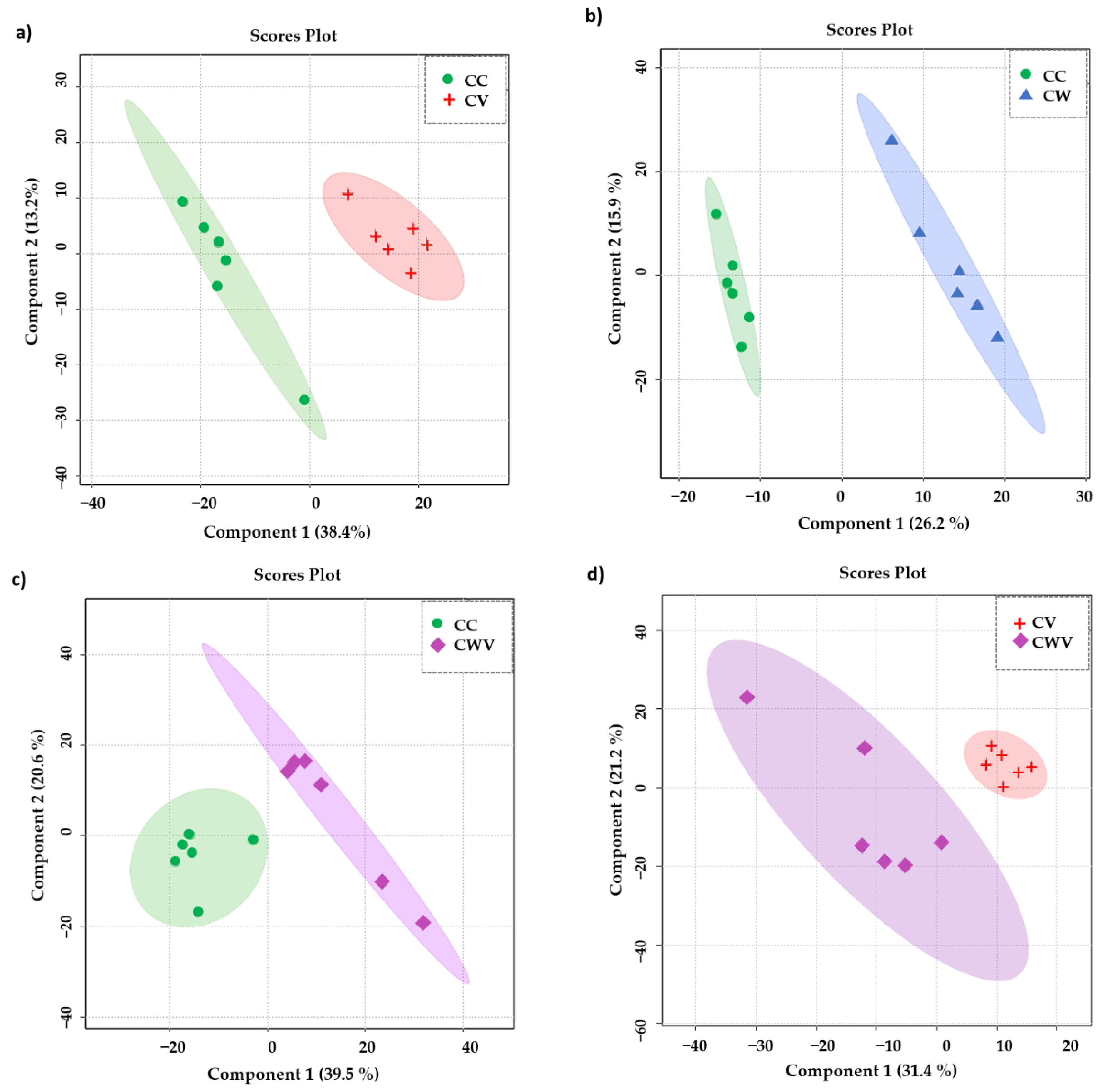
2.5. Chemometric Analysis
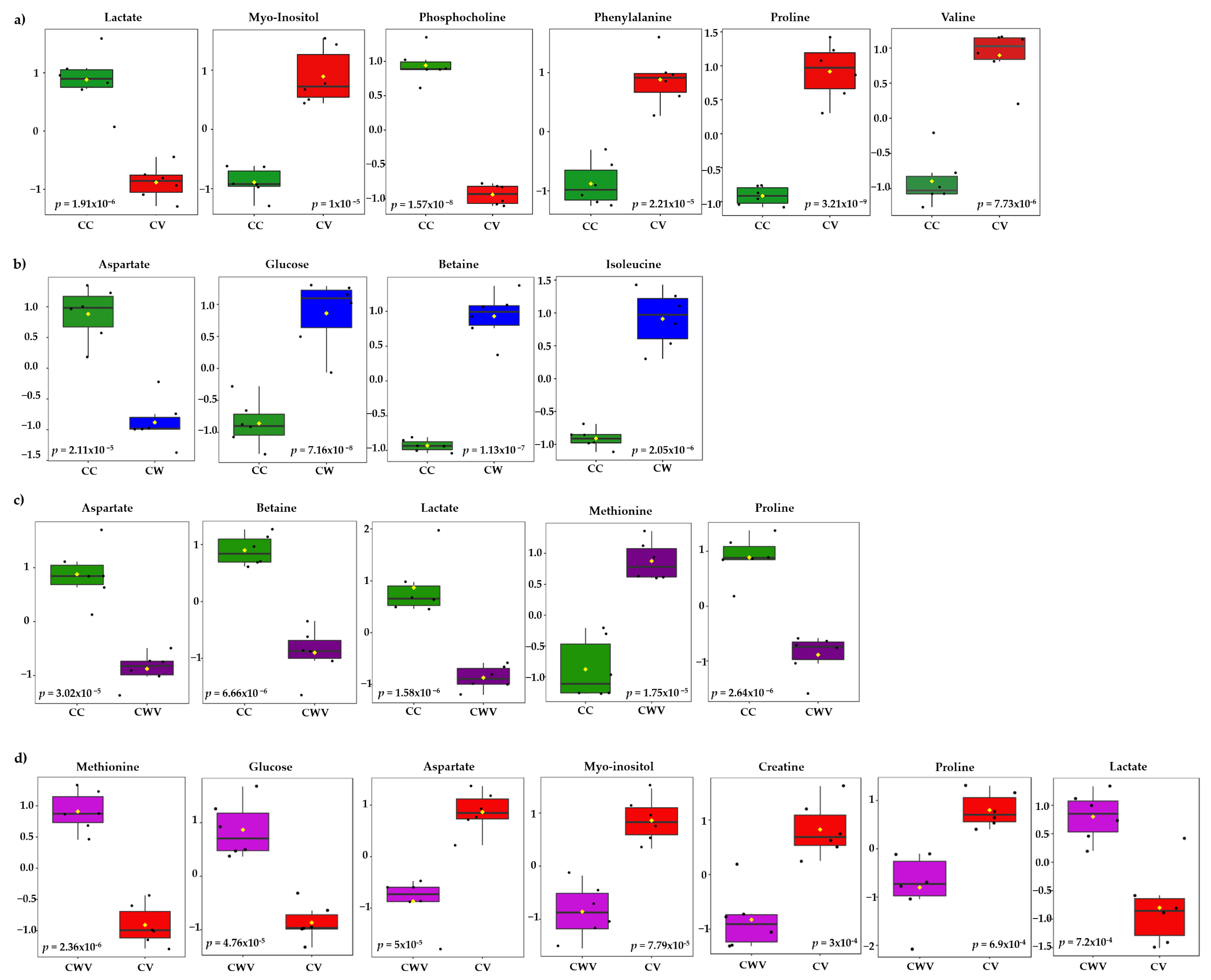
2.6. Metabolite Discrimination
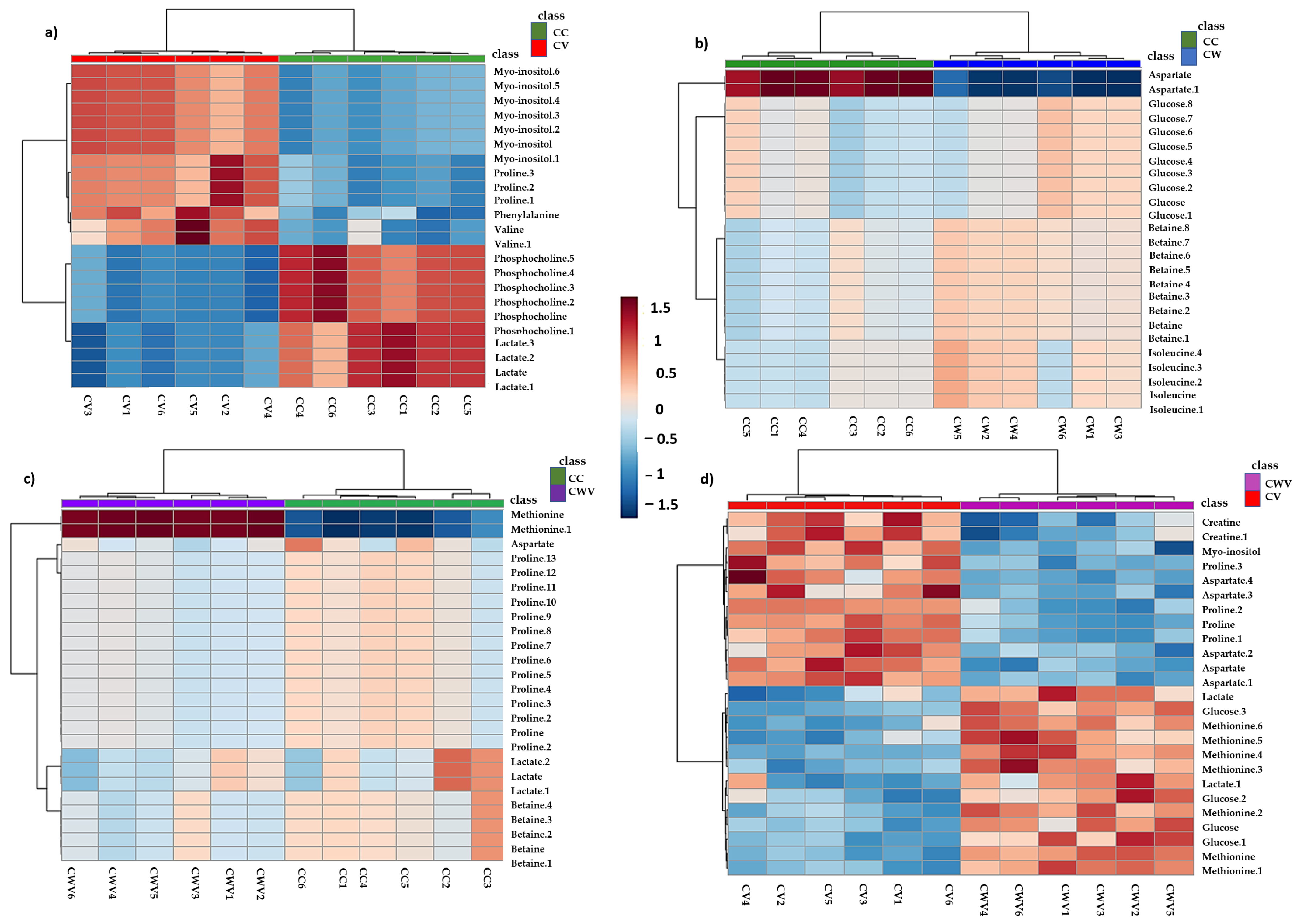
2.7. Heat Map Analysis of Metabolites
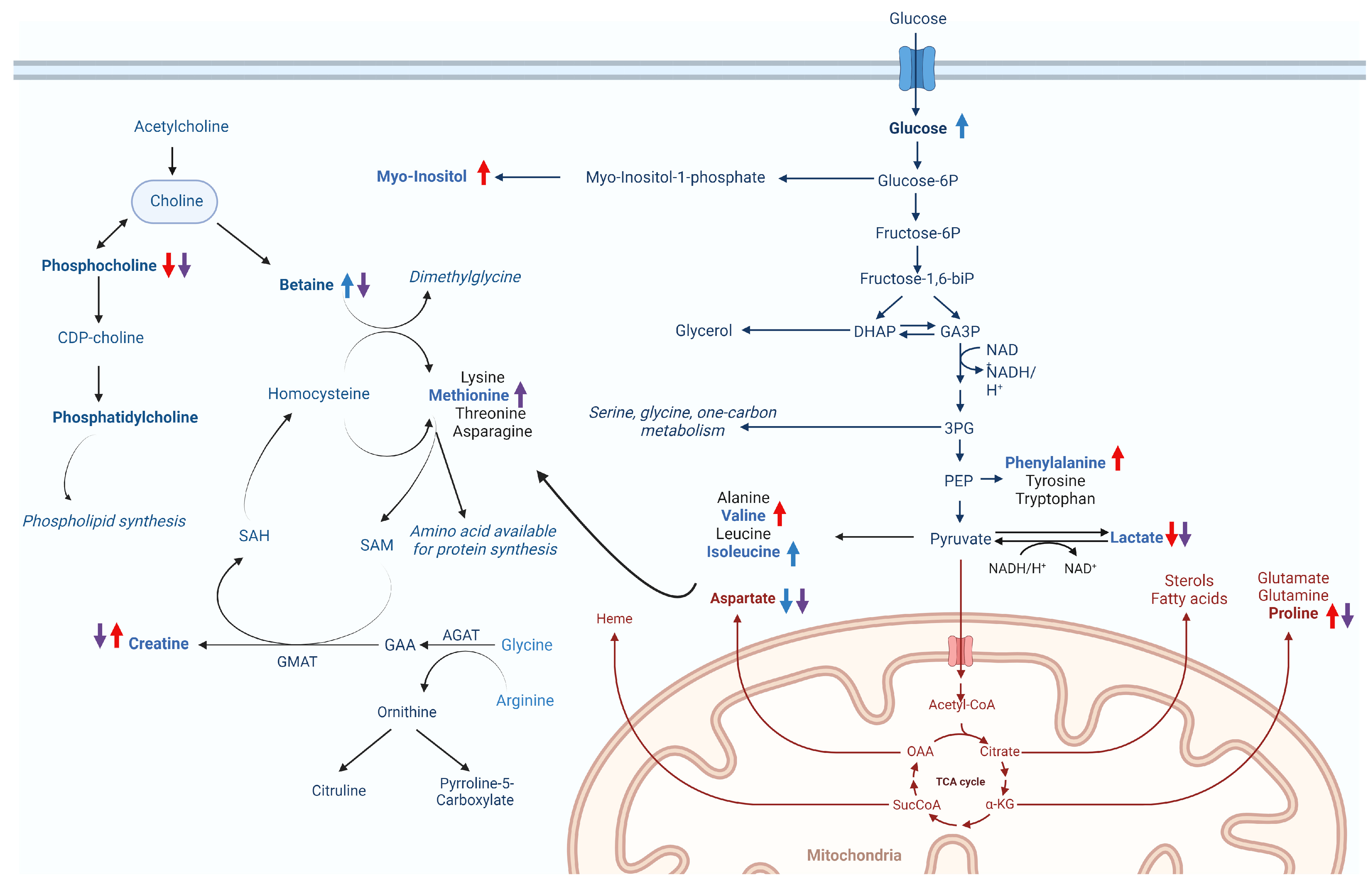
3. Discussion
4. Materials and Methods
4.1. Cloning, Expression and Purification of CHIKV nsP2pro
4.2. Viral Protease Inhibition Assay
4.3. Cell, Virus and Reagents
4.4. Cytotoxicity Assay
4.5. Virus Infection and Titration
4.6. Antiviral In Vitro Activity Assay
4.7. HR-MAS 1H-NMR Sample Preparation
4.8. HR-MAS 1H-NMR and 2D Analysis
4.9. Data Preprocessing, Statistical Analysis and Metabolite Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bellone, R.; Failloux, A.B. The Role of Temperature in Shaping Mosquito-Borne Viruses Transmission. Front. Microbiol. 2020, 11, 584846. [Google Scholar]
- Young, P.R. Arboviruses: A Family on the Move. Adv. Exp. Med. Biol. 2018, 1062, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Feranmi Folahan, F. Neglected Tropical Diseases: Progress and Expectations. Lancet Microbe 2023, 4, e137–e138. [Google Scholar] [CrossRef] [PubMed]
- Mayer, S.V.; Tesh, R.B.; Vasilakis, N. The Emergence of Arthropod-Borne Viral Diseases: A Global Prospective on Dengue, Chikungunya and Zika Fevers. Acta Trop. 2017, 166, 155–163. [Google Scholar] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global Trends in Emerging Infectious Diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Vairo, F.; Haider, N.; Kock, R.; Ntoumi, F.; Ippolito, G.; Zumla, A. Chikungunya: Epidemiology, Pathogenesis, Clinical Features, Management, and Prevention. Infect. Dis. Clin. N. Am. 2019, 33, 1003–1025. [Google Scholar] [CrossRef]
- van Duijl-Richter, M.K.S.; Hoornweg, T.E.; Rodenhuis-Zybert, I.A.; Smit, J.M. Early Events in Chikungunya Virus Infection—From Virus Cell Binding to Membrane Fusion. Viruses 2015, 7, 3647–3674. [Google Scholar] [CrossRef]
- Burt, F.J.; Chen, W.; Miner, J.J.; Lenschow, D.J.; Merits, A.; Schnettler, E.; Kohl, A.; Rudd, P.A.; Taylor, A.; Herrero, L.J.; et al. Chikungunya Virus: An Update on the Biology and Pathogenesis of This Emerging Pathogen. Lancet Infect. Dis. 2017, 17, e107–e117. [Google Scholar] [CrossRef]
- Khan, N.; Bhat, R.; Patel, A.K.; Ray, P. Discovery of Small Molecule Inhibitors of Chikungunya Virus Proteins (NsP2 and E1) Using in Silico Approaches. J. Biomol. Struct. Dyn. 2021, 39, 1373–1385. [Google Scholar] [CrossRef]
- Strauss, J.H.; Strauss, E.G. The Alphaviruses: Gene Expression, Replication, and Evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef]
- Chopra, A.; Saluja, M.; Venugopalan, A. Effectiveness of Chloroquine and Inflammatory Cytokine Response in Patients With Early Persistent Musculoskeletal Pain and Arthritis Following Chikungunya Virus Infection. Arthritis Rheymatol. 2014, 66, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Briolant, S.; Garin, D.; Scaramozzino, N.; Jouan, A.; Crance, J.M. In Vitro Inhibition of Chikungunya and Semliki Forest Viruses Replication by Antiviral Compounds: Synergistic Effect of Interferon-α and Ribavirin Combination. Antivir. Res. 2004, 61, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, B.; Froeyen, M.; Dallmeier, K.; Delang, L.; Guerrero, N.S.; Tas, A.; Que, G.; Jochmans, D.; Herdewijn, P.; Bello, F.; et al. Mutations in the Chikungunya Virus Non-Structural Proteins Cause Resistance to Favipiravir (T-705), a Broad-Spectrum Antiviral. J. Antimicrob. Chemother. 2014, 69, 2770–2784. [Google Scholar] [CrossRef]
- Warter, L.; Lee, C.Y.; Thiagarajan, R.; Lebecque, S.; Lin, R.T.P.; Ng, L.F.P.; Abastado, J.; Wang, C.; Nardin, A.; Warter, L.; et al. Chikungunya Virus Envelope-Specific Human Monoclonal Antibodies with Broad Neutralization Potency. J. Immunol. 2011, 186, 3258–3264. [Google Scholar] [CrossRef]
- Delang, L.; Ng, L.F.P.; Priego, E.; Delang, L.; Ng, L.F.P.; Priego, E. Chikungunya Virus Drug Discovery: Still a Long Way to Go? Expert. Opin. Drug Discov. 2019, 14, 855–866. [Google Scholar] [CrossRef]
- Deeba, F.; Islam, A.; Kazim, S.N.; Naqvi, I.H.; Broor, S.; Ahmed, A.; Parveen, S. Chikungunya Virus: Recent Advances in Epidemiology, Host Pathogen Interaction & Vaccine Strategies. Pathog. Dis. 2016, 74, ftv119. [Google Scholar] [CrossRef]
- Li, X.; Wang, T.; Liu, J.; Liu, Y.; Zhang, J.; Lin, J.; Zhao, Z.; Chen, D. Effect and Mechanism of Wedelolactone as Antioxidant-Coumestan on OH-Treated Mesenchymal Stem Cells. Arab. J. Chem. 2020, 13, 184–192. [Google Scholar] [CrossRef]
- Yuan, F.; Chen, J.; Sun, P.; Guan, S.; Xu, J. Wedelolactone Inhibits LPS-Induced pro-Inflammation via NF-KappaB Pathway. J. Biomed. Sci. 2013, 20, 84. [Google Scholar] [CrossRef]
- Kobori, M.; Yang, Z.; Gong, D.; Heissmeyer, V.; Zhu, H.; Jung, Y.; Gakidis, M.A.M.; Rao, A.; Sekine, T.; Ikegami, F.; et al. Wedelolactone Suppresses LPS-Induced Caspase-11 Expression by Directly Inhibiting the IKK Complex. Cell Death Differ. 2004, 11, 123–130. [Google Scholar] [CrossRef]
- Pan, H.; Lin, Y.; Dou, J.; Fu, Z.; Yao, Y.; Ye, S.; Zhang, S.; Wang, N.; Liu, A.; Li, X.; et al. Wedelolactone Facilitates Ser/Thr Phosphorylation of NLRP3 Dependent on PKA Signalling to Block Inflammasome Activation and Pyroptosis. Cell Prolif. 2020, 53, e12868. [Google Scholar] [CrossRef]
- Zhu, M.-M.; Wang, L.; Yang, D.; Li, C.; Pang, S.-T.; Li, X.-H.; Li, R.; Yang, B.; Lian, Y.P.; Ma, L.; et al. Wedelolactone Alleviates Doxorubicin-Induced Inflammation and Oxidative Stress Damage of Podocytes by IκK/IκB/NFκB Pathway. Biomed. Pharmacother. 2019, 117, 109088. [Google Scholar] [CrossRef] [PubMed]
- Qingqiong, L.; Ding, J.; Zhu, P.; Chen, F.; Xu, L. Hepatoprotective Effect of Wedelolactone against Concanavalin A-Induced Liver Injury in Mice. Am. J. Chin. Med. 2018, 46, 819–833. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, J.; Cao, Y.; Xu, C.; Li, R.; Pan, Y.; Chen, X. Wedelolactone Exhibits Anti- Fi Brotic Effects on Human Hepatic Stellate. Eur. J. Pharmacol. 2013, 714, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Peng, L.; Yang, L.; Xu, X.; Li, W.; Luo, X. Wedelolactone Regulates Lipid Metabolism and Improves Hepatic Steatosis Partly by AMPK Activation and Up-Regulation of Expression of PPAR α/LPL and LDLR. PLoS ONE 2015, 10, e0132720. [Google Scholar] [CrossRef]
- Hsieh, C.; Kuo, P.; Hou, M.; Hung, J.; Chang, F. Wedelolactone Inhibits Breast Cancer-Induced Osteoclastogenesis by Decreasing Akt/MTOR Signaling. Int. J. Oncol. 2015, 46, 555–562. [Google Scholar] [CrossRef]
- Nehybova, T.; Smarda, J.; Daniel, L.; Brezovsky, J.; Benes, P. Wedelolactone Induces Growth of Breast Cancer Cells by Stimulation of Estrogen Receptor Signalling. J. Steroid Biochem. Mol. Biol. 2015, 152, 76–83. [Google Scholar] [CrossRef]
- Sarveswaran, S.; Ghosh, R.; Parikh, R.; Ghosh, J. Wedelolactone, an Anti-Inflammatory Botanical, Interrupts c-Myc Oncogenic Signaling and Synergizes with Enzalutamide to Induce Apoptosis in Prostate Cancer Cells. Mol. Cancer Ther. 2016, 15, 2791–2801. [Google Scholar]
- Benes, P.; Knopfova, L.; Trcka, F.; Nemajerova, A.; Pinheiro, D.; Soucek, K.; Fojta, M.; Smarda, J. Inhibition of Topoisomerase II a: Novel Function of Wedelolactone. Cancer Lett. 2011, 303, 29–38. [Google Scholar] [CrossRef]
- Chen, H.; Gao, S.; Li, J.; Liu, D.; Sheng, C. Wedelolactone Disrupts the Interaction of EZH2-EED Complex and Inhibits PRC2-Dependent Cancer. Oncotarget 2015, 6, 13049–13059. [Google Scholar] [CrossRef]
- Sarveswaran, S.; Gautam, S.C.; Ghosh, J. Wedelolactone, a Medicinal Plant-Derived Coumestan, Induces Caspase-Dependent Apoptosis in Prostate Cancer Cells via Downregulation of PKC ε without Inhibiting Akt. Int. J. Oncol. 2012, 41, 2191–2199. [Google Scholar] [CrossRef]
- Tewtrakul, S.; Subhadhirasakul, S.; Cheenpracha, S.; Karalai, C. HIV-1 Protease and HIV-1 Integrase Inhibitory Substances from Eclipta Prostrata. Phytother. Res. 2007, 21, 1092–1095. [Google Scholar] [CrossRef] [PubMed]
- Manvar, D.; Mishra, M.; Kumar, S.; Pandey, V.N. Identification and Evaluation of Anti Hepatitis C Virus Phytochemicals from Eclipta Alba. J. Ethnopharmacol. 2012, 144, 545–554. [Google Scholar] [CrossRef]
- Svrlanska, A.; Ruhland, A.; Marschall, M.; Reuter, N. Wedelolactone Inhibits Human Cytomegalovirus Replication by Targeting Distinct Steps of the Viral Replication Cycle. Antivir. Res. 2020, 174, 13–35. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Vermathen, M.; Diserens, G.; Vermathen, P.; Furrer, J. Metabolic Profiling of Cells in Response to Drug Treatment Using 1 H High-Resolution Magic Angle Spinning (HR-MAS) NMR Spectroscopy. Bioorganomet. Chem. Mech. 2017, 71, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Letertre, M.P.M.; Giraudeau, P.; Tullio, P. De Nuclear Magnetic Resonance Spectroscopy in Clinical Metabolomics and Personalized Medicine: Current Challenges and Perspectives. Front. Mol. Biosci. 2021, 8, 698337. [Google Scholar] [CrossRef]
- Weybright, P.; Millis, K.; Campbell, N.; Cory, D.G.; Singer, S. Gradient, High-Resolution, Magic Angle Spinning H Nuclear Magnetic Resonance Spectroscopy of Intact Cells. Magn. Reson. Med. 1998, 39, 337–345. [Google Scholar] [CrossRef]
- Eberle, R.J.; Olivier, D.S.; Pacca, C.C.; Avilla, C.M.S.; Nogueira, M.L.; Amaral, M.S.; Willbold, D.; Arni, R.K. In Vitro Study of Hesperetin and Hesperidin as Inhibitors of Zika and Chikungunya Virus Proteases. PLoS ONE 2021, 16, e0246319. [Google Scholar] [CrossRef]
- Ivanova, L.; Rausalu, K.; Žusinaite, E.; Tammiku-Taul, J.; Merits, A.; Karelson, M. 1,3-Thiazolbenzamide Derivatives as Chikungunya Virus NsP2 Protease Inhibitors. ACS Omega 2021, 6, 5786–5794. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, 469–472. [Google Scholar] [CrossRef]
- Zhang, R.; Qin, X.; Kong, F.; Chen, P.; Pan, G. Improving Cellular Uptake of Therapeutic Entities through Interaction with Components of Cell Membrane. Drug Deliv. 2019, 26, 328–342. [Google Scholar] [PubMed]
- El-bacha, T.; Menezes, M.T.; Azevedo, M.C.; Sola-penna, M.; Poian, A.T. Da Mayaro Virus Infection Alters Glucose Metabolism in Cultured Cells through Activation of the Enzyme 6-Phosphofructo 1-Kinase. Mol. Cell. Biochem. 2004, 266, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef]
- Mulukutla, B.C.; Khan, S.; Lange, A.; Hu, W. Glucose Metabolism in Mammalian Cell Culture: New Insights for Tweaking Vintage Pathways. Trends Biotechnol. 2010, 28, 476–484. [Google Scholar] [CrossRef]
- Polcicova, K.; Badurova, L.; Tomaskova, J. Metabolic Reprogramming as a Feast for Virus Replication. Acta Virol. 2020, 64, 201–215. [Google Scholar]
- Deberardinis, R.J.; Cheng, T. Q’s next: The Diverse Functions of Glutamine in Metabolism, Cell Biology and Cancer. Oncogene 2010, 29, 313–324. [Google Scholar] [CrossRef]
- Sa, F.J.; Ange, C.; Rodr, M. The Role of Tricarboxylic Acid Cycle Metabolites in Viral Infections. Front. Cell Infect. Microbiol. 2021, 11, 725043. [Google Scholar] [CrossRef]
- Newsholme, P.; Procopio, J.; Maria, M.; Lima, R.; Pithon-curi, T.C. Glutamine and Glutamate—Their Central Role in Cell Metabolism and Function. Cell Biochem. Funct. 2003, 21, 1–9. [Google Scholar] [PubMed]
- Findlay, J.S.; Ulaeto, D.; Findlay, J.S. Semliki Forest Virus and Sindbis Virus, but Not Vaccinia Virus, Require Glycolysis for Optimal Replication. J. General. Virol. 2015, 96, 2693–2696. [Google Scholar] [CrossRef]
- Eagle, H. Amino Acid Metabolism Cell Cultures in Mammalian. Science 1955, 130, 432–437. [Google Scholar] [CrossRef]
- Bastin, G.; Chotteau, V.; Wouwer, A. Vande Metabolic Flux Analysis of VERO Cells under Various Culture Conditions. Processes 2021, 9, 2097. [Google Scholar] [CrossRef]
- Thaker, S.K.; Ch, J.; Christofk, H.R. Viral Hijacking of Cellular Metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 7th ed.; W.H. Freeman: New York, NY, USA, 2017. [Google Scholar]
- Holub, B.J. Metabolism and Function of Myo-Inositol and Inositol Phospholipds. Annu. Rev. Nutr. 1986, 6, 563–597. [Google Scholar] [CrossRef] [PubMed]
- Tu-sekine, B.; Kim, S.F. The Inositol Phosphate System—A Coordinator of Metabolic Adaptability. Int. J. Mol. Sci. 2022, 23, 6747. [Google Scholar] [PubMed]
- Zhang, Z.; He, G.; Filipowicz, N.A.; Randall, G.; Belov, G.A.; Kopek, B.G.; Wang, X. Host Lipids in Positive-Strand RNA Virus Genome Replication. Front. Microbiol. 2019, 10, 286. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, V.; Heaton, N.S.; Randall, G. Lipids at the Interface of Virus-Host Interactions. Curr. Opin. Microbiol. 2013, 15, 512–518. [Google Scholar] [CrossRef]
- Vance, J.E. Phospholipid Synthesis and Transport in Mammalian Cells. Traffic 2015, 16, 1–18. [Google Scholar]
- Li, Z.; Vance, D.E. Phosphatidylcholine and Choline Homeostasis. J. Lipid Res. 2008, 49, 1187–1194. [Google Scholar] [CrossRef]
- Perera, R.; Riley, C.; Isaac, G.; Hopf-Jannasch, A.S.; Moore, R.J.; Weitz, K.W.; Pasa-Tolic, L.; Metz, T.O.; Adamec, J.; Kuhn, R.J. Dengue Virus Infection Perturbs Lipid Homeostasis in Infected Mosquito Cells. PLoS Pathog. 2012, 8, e1002584. [Google Scholar] [CrossRef]
- Vance, D.E.; Trip, E.M.; Paddon, H.B. Poliovirus Increases Phosphatidylcholine Biosynthesis in HeLa Cells by Stimulation of the Rate-Limiting Reaction Catalyzed by CTP: Phosphocholine cytidylyltransferase. J. Biol. Chem. 1980, 255, 1064–1069. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Z.; Chukkapalli, V.; Nchoutmboube, J.A.; Li, J.; Randall, G. Positive-Strand RNA Viruses Stimulate Host Phosphatidylcholine Synthesis at Viral Replication Sites. Proc. Natl. Acad. Sci. USA 2016, 113, E1064–E1073. [Google Scholar] [CrossRef]
- Holeček, M. Roles of Malate and Aspartate in Gluconeogenesis in Various Physiological and Pathological States. Metabolism 2023, 145, 155614. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, J.H.; Noh, S.; Hur, H.J.; Sung, M.J.; Hwang, J.T.; Park, J.H.; Yang, H.J.; Kim, M.S.; Kwon, D.Y.; et al. Metabolomic Analysis of Livers and Serum from High-Fat Diet Induced Obese Mice. J. Proteome Res. 2011, 10, 722–731. [Google Scholar] [CrossRef]
- Craig, S.A.S. Betaine in Human Nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, H.; Li, H.; Lai, F.; Li, X.; Tang, Y.; Min, T.; Wu, H. Antioxidant Mechanism of Betaine without Free Radical Scavenging Ability. J. Agric. Food Chem. 2016, 64, 7921–7930. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.; Finhelstein, D.; Kyle, E. Methionine Metabolism in Mammals: Kinetic Study of Betaine-Homocysteine Methyltransferase. Arch. Biochem. Biophys. 1972, 153, 320–324. [Google Scholar]
- Lauinger, L.; Kaiser, P. Sensing and Signaling of Methionine Metabolism. Metabolites 2021, 11, 83. [Google Scholar] [CrossRef]
- Joubert, P.E.; Werneke, S.; De La Calle, C.; Guivel-Benhassine, F.; Giodini, A.; Peduto, L.; Levine, B.; Schwartz, O.; Lenschow, D.; Albert, M.L. Chikungunya-Induced Cell Death Is Limited by ER and Oxidative Stress-Induced Autophagy. Autophagy 2012, 8, 1261–1263. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, M.; Li, H.; Cai, L.; He, H.; Wu, Q.; Yang, L. L-Methionine Activates Nrf2-ARE Pathway to Induce Endogenous Antioxidant Activity for Depressing ROS-Derived Oxidative Stress in Growing Rats. J. Sci. Food Agric. 2019, 99, 4849–4862. [Google Scholar] [CrossRef]
- Walvekar, A.S.; Srinivasan, R.; Gupta, R.; Laxman, S. Methionine Coordinates a Hierarchically Organized Anabolic Program Enabling Proliferation. Mol. Biol. Cell 2018, 29, 3183–3200. [Google Scholar] [CrossRef]
- Sharma, S.; Trivedi, S.; Pandey, T.; Ranjan, S.; Trivedi, M.; Pandey, R. Wedelolactone Mitigates Parkinsonism Via Alleviating Oxidative Stress and Mitochondrial Dysfunction Through NRF2/SKN-1. Mol. Neurobiol. 2021, 58, 65–77. [Google Scholar] [PubMed]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and Creatinine Metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, D.A.; Kreider, R.B.; Stout, J.R.; Forero, D.A.; Kerksick, C.M.; Roberts, M.D.; Rawson, E.S. Metabolic Basis of Creatine in Health and Disease: A Bioinformatics-Assisted Review. Nutrients 2021, 13, 1238. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Aizaki, H.; Matsuda, M.; Shinkai-Ouchi, F.; Inoue, Y.; Murakami, K.; Shoji, I.; Kawakami, H.; Matsuura, Y.; Lai, M.M.C.; et al. Involvement of Creatine Kinase B in Hepatitis C Virus Genome Replication through Interaction with the Viral NS4A Protein. J. Virol. 2009, 83, 5137–5147. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. Creatine: Endogenous Metabolite, Dietary, and Therapeutic Supplement. Annu. Rev. Nutr. 2007, 27, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Eberle, R.J.; Olivier, D.S.; Amaral, M.S.; Pacca, C.C.; Nogueira, M.L.; Arni, R.K.; Willbold, D.; Coronado, M.A. Riboflavin, a Potent Neuroprotective Vitamin: Focus on Flavivirus and Alphavirus Proteases. Microorganisms 2022, 10, 1331. [Google Scholar] [CrossRef]
- Nunes, M.R.T.; Faria, N.R.; de Vasconcelos, J.M.; Golding, N.; Kraemer, M.U.G.; de Oliveira, L.F.; da Silva Azevedo, R.d.S.; da Silva, D.E.A.; da Silva, E.V.P.; da Silva, S.P.; et al. Emergence and Potential for Spread of Chikungunya Virus in Brazil. BMC Med. 2015, 13, 102. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, 521–526. [Google Scholar] [CrossRef]
- Ulrich, E.L.; Akutsu, H.; Doreleijers, J.F.; Harano, Y.; Ioannidis, Y.E.; Lin, J.; Livny, M.; Mading, S.; Maziuk, D.; Miller, Z.; et al. BioMagResBank. Nucleic Acids Res. 2008, 36, 402–408. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Hinzman, A.A.; Kang, E.L.; Szczesniak, R.D.; Lu, L.J.; Lu, L.J. Computational and Statistical Analysis of Metabolomics Data. Metabolomics 2015, 11, 1492–1513. [Google Scholar] [CrossRef]
- Barker, M.; Rayens, W. Partial Least Squares for Discrimination. J. Chemom. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Berrueta, L.A.; Alonso-salces, R.M. Supervised Pattern Recognition in Food Analysis. J. Chromatogr. A 2007, 1158, 196–214. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classes | Accuracy | R2 | Q2 |
|---|---|---|---|
| CC × CV | 0.91 | 0.99 | 0.78 |
| CC × CW | 1.0 | 0.99 | 0.90 |
| CC × CWV | 1.0 | 0.99 | 0.85 |
| CV × CWV | 0.91 | 0.99 | 0.72 |
| CC × CV × CW × CWV | 0.87 | 0.98 | 0.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peinado, R.d.S.; Martins, L.G.; Pacca, C.C.; Saivish, M.V.; Borsatto, K.C.; Nogueira, M.L.; Tasic, L.; Arni, R.K.; Eberle, R.J.; Coronado, M.A. HR-MAS NMR Metabolomics Profile of Vero Cells under the Influence of Virus Infection and nsP2 Inhibitor: A Chikungunya Case Study. Int. J. Mol. Sci. 2024, 25, 1414. https://doi.org/10.3390/ijms25031414
Peinado RdS, Martins LG, Pacca CC, Saivish MV, Borsatto KC, Nogueira ML, Tasic L, Arni RK, Eberle RJ, Coronado MA. HR-MAS NMR Metabolomics Profile of Vero Cells under the Influence of Virus Infection and nsP2 Inhibitor: A Chikungunya Case Study. International Journal of Molecular Sciences. 2024; 25(3):1414. https://doi.org/10.3390/ijms25031414
Chicago/Turabian StylePeinado, Rafaela dos S., Lucas G. Martins, Carolina C. Pacca, Marielena V. Saivish, Kelly C. Borsatto, Maurício L. Nogueira, Ljubica Tasic, Raghuvir K. Arni, Raphael J. Eberle, and Mônika A. Coronado. 2024. "HR-MAS NMR Metabolomics Profile of Vero Cells under the Influence of Virus Infection and nsP2 Inhibitor: A Chikungunya Case Study" International Journal of Molecular Sciences 25, no. 3: 1414. https://doi.org/10.3390/ijms25031414
APA StylePeinado, R. d. S., Martins, L. G., Pacca, C. C., Saivish, M. V., Borsatto, K. C., Nogueira, M. L., Tasic, L., Arni, R. K., Eberle, R. J., & Coronado, M. A. (2024). HR-MAS NMR Metabolomics Profile of Vero Cells under the Influence of Virus Infection and nsP2 Inhibitor: A Chikungunya Case Study. International Journal of Molecular Sciences, 25(3), 1414. https://doi.org/10.3390/ijms25031414