

Human mtDNA-Encoded Long ncRNAs: Knotty Molecules and Complex Functions


Abstract
1. When Non-Coding Is better: An Overview of ncRNAs
2. Long Non-Coding RNAs Encoded by Mitochondrial Genome
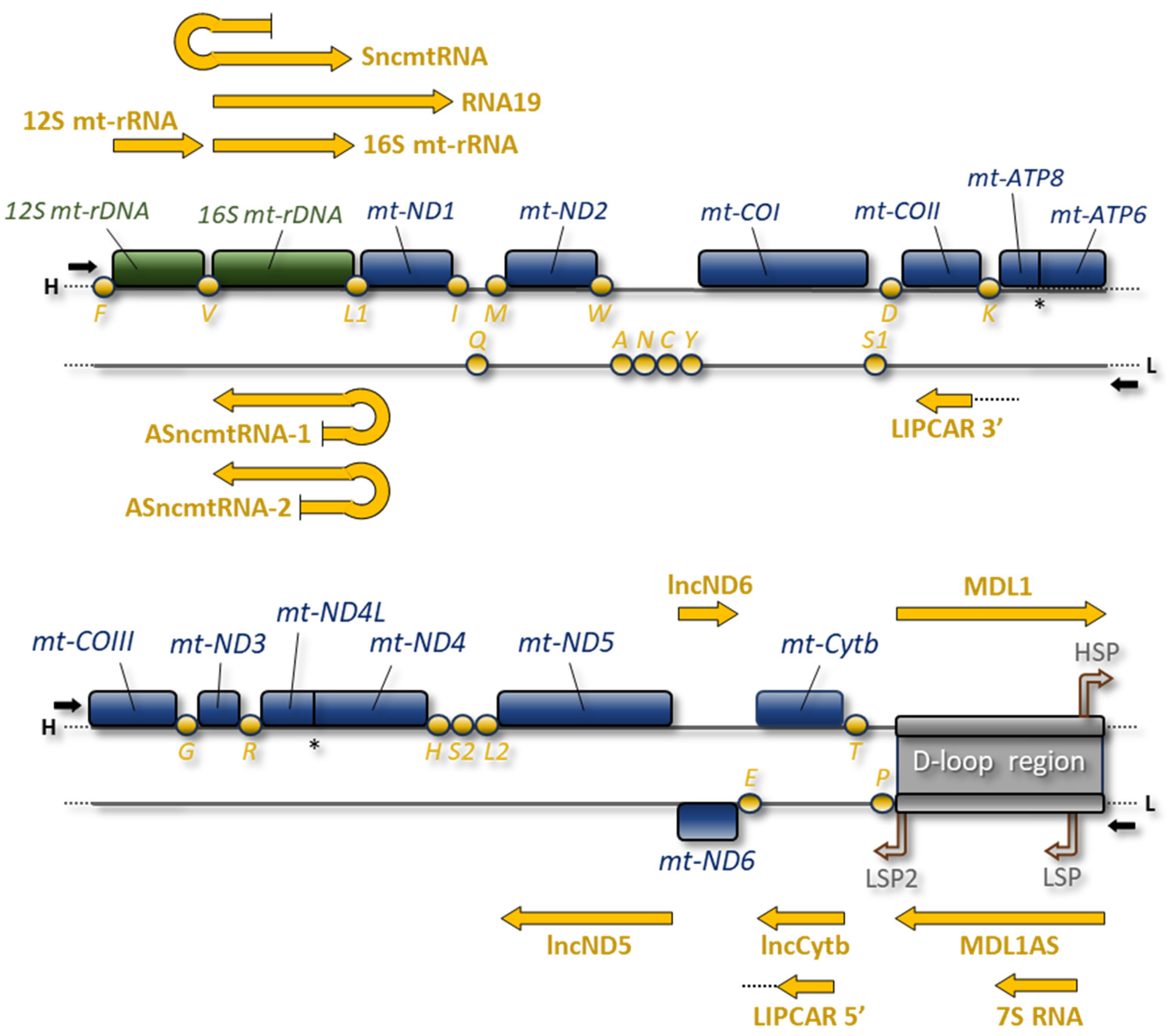
2.1. 16S mt-rRNA and 12S mt-rRNA
2.2. RNA19
2.3. Sense and Antisense ncmtRNAs
2.4. lncND5, lncND6, and lncCytb RNAs
2.5. 7S RNA
2.6. Sense and Antisense Mitochondrial D-Loop 1 RNAs
2.7. LIPCAR
2.8. Mitochondrial Double-Stranded RNAs
2.9. Mitochondrial Circular RNAs
3. Pathological Aspects Related to mtDNA-Encoded lncRNAs
4. Other Mitochondria-Associated lncRNAs
5. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and Their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging Role of Non-coding RNA in Health and Disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Updated Overview. In MicroRNA Profiling: Methods and Protocols; Methods in Molecular Biology; Humana: New York, NY, USA, 2023; Volume 2595, pp. 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.-J.; Liu, J.; Wang, X.; Qu, L.-X. MicroRNA-181 Promotes Prostate Cancer Cell Proliferation by Regulating DAX-1 Expression. Exp. Ther. Med. 2014, 8, 1296–1300. [Google Scholar] [CrossRef] [PubMed]
- Barbato, A.; Iuliano, A.; Volpe, M.; D’Alterio, R.; Brillante, S.; Massa, F.; De Cegli, R.; Carrella, S.; Salati, M.; Russo, A.; et al. Integrated Genomics Identifies MiR-181/TFAM Pathway as a Critical Driver of Drug Resistance in Melanoma. Int. J. Mol. Sci. 2021, 22, 1801. [Google Scholar] [CrossRef]
- Magen, I.; Yacovzada, N.S.; Yanowski, E.; Coenen-Stass, A.; Grosskreutz, J.; Lu, C.-H.; Greensmith, L.; Malaspina, A.; Fratta, P.; Hornstein, E. Circulating MiR-181 Is a Prognostic Biomarker for Amyotrophic Lateral Sclerosis. Nat. Neurosci. 2021, 24, 1534–1541. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qiu, M.; Shen, M.; Dong, S.; Ye, G.; Shi, X.; Sun, M. The Emerging Regulatory Roles of Long Non-Coding RNAs Implicated in Cancer Metabolism. Mol. Ther. 2021, 29, 2209–2218. [Google Scholar] [CrossRef]
- Walther, K.; Schulte, L.N. The Role of LncRNAs in Innate Immunity and Inflammation. RNA Biol. 2021, 18, 587–603. [Google Scholar] [CrossRef]
- Wei, S.; Fan, Q.; Yang, L.; Zhang, X.; Ma, Y.; Zong, Z.; Hua, X.; Su, D.; Sun, H.; Li, H.; et al. Promotion of Glycolysis by HOTAIR through GLUT1 Upregulation via MTOR Signaling. Oncol. Rep. 2017, 38, 1902–1908. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, P.; Wang, L.; Piao, H.; Ma, L. Long Non-Coding RNA HOTAIR in Carcinogenesis and Metastasis. Acta Biochim. Biophys. Sin. 2014, 46, 1–5. [Google Scholar] [CrossRef]
- Oo, J.A.; Brandes, R.P.; Leisegang, M.S. Long Non-Coding RNAs: Novel Regulators of Cellular Physiology and Function. Pflug. Arch. 2022, 474, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Shang, R.; Lee, S.; Senavirathne, G.; Lai, E.C. MicroRNAs in Action: Biogenesis, Function and Regulation. Nat. Rev. Genet. 2023, 24, 816–833. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.-W.; Wang, Y.; Chen, L.-L. Cellular Functions of Long Noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Hu, J.; Yin, W. Systematic Identification of Non-Coding RNAs. In Non-Coding RNAs in Complex Diseases: A Bioinformatics Perspective; Li, X., Xu, J., Xiao, Y., Ning, S., Zhang, Y., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2018; pp. 9–18. ISBN 9789811307195. [Google Scholar]
- Chen, Q.; Meng, X.; Liao, Q.; Chen, M. Versatile Interactions and Bioinformatics Analysis of Noncoding RNAs. Brief. Bioinform. 2019, 20, 1781–1794. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.F.; Lee, E.S. Non-Coding RNA: What Is Functional and What Is Junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, J.; Uszczynska-Ratajczak, B.; Carbonell, S.; Pérez-Lluch, S.; Abad, A.; Davis, C.; Gingeras, T.R.; Frankish, A.; Harrow, J.; Guigo, R.; et al. High-Throughput Annotation of Full-Length Long Noncoding RNAs with Capture Long-Read Sequencing. Nat. Genet. 2017, 49, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of LncRNAs. In Long Non Coding RNA Biology; Advances in Experimental Medicine and Biology; Springer: Singapore, 2017; Volume 1008, pp. 1–46. [Google Scholar] [CrossRef]
- Carbonell Sala, S.; Uszczyńska-Ratajczak, B.; Lagarde, J.; Johnson, R.; Guigó, R. Annotation of Full-Length Long Noncoding RNAs with Capture Long-Read Sequencing (CLS). Methods Mol. Biol. 2021, 2254, 133–159. [Google Scholar] [CrossRef]
- Bruni, F. Mitochondria: From Physiology to Pathology. Life 2021, 11, 991. [Google Scholar] [CrossRef]
- Giacomello, M.; Pyakurel, A.; Glytsou, C.; Scorrano, L. The Cell Biology of Mitochondrial Membrane Dynamics. Nat. Rev. Mol. Cell Biol. 2020, 21, 204–224. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, L.; Wang, R.R.; Hu, J.-F.; Cui, J. The Effects of Mitochondria-Associated Long Noncoding RNAs in Cancer Mitochondria: New Players in an Old Arena. Crit. Rev. Oncol. Hematol. 2018, 131, 76–82. [Google Scholar] [CrossRef]
- Noh, J.H.; Kim, K.M.; Abdelmohsen, K.; Yoon, J.-H.; Panda, A.C.; Munk, R.; Kim, J.; Curtis, J.; Moad, C.A.; Wohler, C.M.; et al. HuR and GRSF1 Modulate the Nuclear Export and Mitochondrial Localization of the LncRNA RMRP. Genes. Dev. 2016, 30, 1224–1239. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yoshitomi, T.; Hu, J.-F.; Cui, J. Long Noncoding RNAs Coordinate Functions between Mitochondria and the Nucleus. Epigenetics Chromatin 2017, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Wallet, C.; Iqbal, R.K.; Gualberto, J.M.; Lotfi, F. Organellar Non-Coding RNAs: Emerging Regulation Mechanisms. Biochimie 2015, 117, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, S.; Wang, P.; Wang, G. Non-Coding RNA Regulated Cross-Talk between Mitochondria and Other Cellular Compartments. Front. Cell Dev. Biol. 2021, 9, 688523. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.G.; Mutti, C.D.; Shi, Y.; Xie, X.; Zhu, X.; Silva-Pinheiro, P.; Menger, K.E.; Díaz-Maldonado, H.; Wei, W.; Nicholls, T.J.; et al. The Human Mitochondrial Genome Contains a Second Light Strand Promoter. Mol. Cell 2022, 82, 3646–3660.e9. [Google Scholar] [CrossRef] [PubMed]
- Barshad, G.; Marom, S.; Cohen, T.; Mishmar, D. Mitochondrial DNA Transcription and Its Regulation: An Evolutionary Perspective. Trends Genet. 2018, 34, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Bruni, F.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M. Human Mitochondrial Nucleases. FEBS J. 2017, 284, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, J.; Frank, P.; Löffler, E.; Bennett, K.L.; Gerner, C.; Rossmanith, W. RNase P without RNA: Identification and Functional Reconstitution of the Human Mitochondrial TRNA Processing Enzyme. Cell 2008, 135, 462–474. [Google Scholar] [CrossRef]
- Rossmanith, W. Localization of Human RNase Z Isoforms: Dual Nuclear/Mitochondrial Targeting of the ELAC2 Gene Product by Alternative Translation Initiation. PLoS ONE 2011, 6, e19152. [Google Scholar] [CrossRef]
- Siira, S.J.; Rossetti, G.; Richman, T.R.; Perks, K.; Ermer, J.A.; Kuznetsova, I.; Hughes, L.; Shearwood, A.-M.J.; Viola, H.M.; Hool, L.C.; et al. Concerted Regulation of Mitochondrial and Nuclear Non-Coding RNAs by a Dual-Targeted RNase Z. EMBO Rep. 2018, 19, e46198. [Google Scholar] [CrossRef] [PubMed]
- Antonicka, H.; Shoubridge, E.A. Mitochondrial RNA Granules Are Centers for Posttranscriptional RNA Processing and Ribosome Biogenesis. Cell Rep. 2015, 10, 920–932. [Google Scholar] [CrossRef]
- Jedynak-Slyvka, M.; Jabczynska, A.; Szczesny, R.J. Human Mitochondrial RNA Processing and Modifications: Overview. IJMS 2021, 22, 7999. [Google Scholar] [CrossRef] [PubMed]
- Kummer, E.; Ban, N. Mechanisms and Regulation of Protein Synthesis in Mitochondria. Nat. Rev. Mol. Cell Biol. 2021, 22, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Mai, N.; Chrzanowska-Lightowlers, Z.M.A.; Lightowlers, R.N. The Process of Mammalian Mitochondrial Protein Synthesis. Cell Tissue Res. 2017, 367, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Silva, P.; Martinez-Azorin, F.; Micol, V.; Attardi, G. The Human Mitochondrial Transcription Termination Factor (MTERF) Is a Multizipper Protein but Binds to DNA as a Monomer, with Evidence Pointing to Intramolecular Leucine Zipper Interactions. EMBO J. 1997, 16, 1066–1079. [Google Scholar] [CrossRef] [PubMed]
- Kruse, B.; Narasimhan, N.; Attardi, G. Termination of Transcription in Human Mitochondria: Identification and Purification of a DNA Binding Protein Factor That Promotes Termination. Cell 1989, 58, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.; Gaines, G.L.; Attardi, G. The Pattern of Transcription of the Human Mitochondrial RRNA Genes Reveals Two Overlapping Transcription Units. Cell 1983, 34, 151–159. [Google Scholar] [CrossRef]
- Tan, B.G.; Gustafsson, C.M.; Falkenberg, M. Mechanisms and Regulation of Human Mitochondrial Transcription. Nat. Rev. Mol. Cell Biol. 2024, 25, 119–132. [Google Scholar] [CrossRef]
- Lopez Sanchez, M.I.G.; Mercer, T.R.; Davies, S.M.K.; Shearwood, A.-M.J.; Nygård, K.K.A.; Richman, T.R.; Mattick, J.S.; Rackham, O.; Filipovska, A. RNA Processing in Human Mitochondria. Cell Cycle 2011, 10, 2904–2916. [Google Scholar] [CrossRef]
- Boughanem, H.; Böttcher, Y.; Tomé-Carneiro, J.; López de las Hazas, M.-C.; Dávalos, A.; Cayir, A.; Macias-González, M. The Emergent Role of Mitochondrial RNA Modifications in Metabolic Alterations. WIREs RNA 2023, 14, e1753. [Google Scholar] [CrossRef]
- Rebelo-Guiomar, P.; Powell, C.A.; Van Haute, L.; Minczuk, M. The Mammalian Mitochondrial Epitranscriptome. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2019, 1862, 429–446. [Google Scholar] [CrossRef] [PubMed]
- Khawaja, A.; Cipullo, M.; Krüger, A.; Rorbach, J. Insights into Mitoribosomal Biogenesis from Recent Structural Studies. Trends Biochem. Sci. 2023, 48, 629–641. [Google Scholar] [CrossRef]
- Cipullo, M.; Gesé, G.V.; Khawaja, A.; Hällberg, B.M.; Rorbach, J. Structural Basis for Late Maturation Steps of the Human Mitoribosomal Large Subunit. Nat. Commun. 2021, 12, 3673. [Google Scholar] [CrossRef] [PubMed]
- Harper, N.J.; Burnside, C.; Klinge, S. Principles of Mitoribosomal Small Subunit Assembly in Eukaryotes. Nature 2023, 614, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Hilander, T.; Jackson, C.B.; Robciuc, M.; Bashir, T.; Zhao, H. The Roles of Assembly Factors in Mammalian Mitoribosome Biogenesis. Mitochondrion 2021, 60, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Itoh, Y.; Del’Olio, S.; Hassan, A.; Naschberger, A.; Flygaard, R.K.; Nobe, Y.; Izumikawa, K.; Aibara, S.; Andréll, J.; et al. Structure of Mitoribosome Reveals Mechanism of MRNA Binding, TRNA Interactions with L1 Stalk, Roles of Cofactors and RRNA Modifications. bioRxiv 2023. [Google Scholar] [CrossRef]
- Brown, A.; Amunts, A.; Bai, X.; Sugimoto, Y.; Edwards, P.C.; Murshudov, G.; Scheres, S.H.W.; Ramakrishnan, V. Structure of the Large Ribosomal Subunit from Human Mitochondria. Science 2014, 346, 718–722. [Google Scholar] [CrossRef]
- Amunts, A.; Brown, A.; Toots, J.; Scheres, S.H.W.; Ramakrishnan, V. The Structure of the Human Mitochondrial Ribosome. Science 2015, 348, 95–98. [Google Scholar] [CrossRef]
- Lancaster, L.; Lambert, N.J.; Maklan, E.J.; Horan, L.H.; Noller, H.F. The Sarcin-Ricin Loop of 23S RRNA Is Essential for Assembly of the Functional Core of the 50S Ribosomal Subunit. RNA 2008, 14, 1999–2012. [Google Scholar] [CrossRef]
- Moazed, D.; Robertson, J.M.; Noller, H.F. Interaction of Elongation Factors EF-G and EF-Tu with a Conserved Loop in 23S RNA. Nature 1988, 334, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Schmeing, T.M.; Ramakrishnan, V. What Recent Ribosome Structures Have Revealed about the Mechanism of Translation. Nature 2009, 461, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Winther, K.S.; Brodersen, D.E.; Brown, A.K.; Gerdes, K. VapC20 of Mycobacterium Tuberculosis Cleaves the Sarcin-Ricin Loop of 23S RRNA. Nat. Commun. 2013, 4, 2796. [Google Scholar] [CrossRef] [PubMed]
- Bruni, F.; Proctor-Kent, Y.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M. Messenger RNA Delivery to Mitoribosomes–Hints from a Bacterial Toxin. FEBS J. 2021, 288, 437–451. [Google Scholar] [CrossRef] [PubMed]
- King, M.P.; Koga, Y.; Davidson, M.; Schon, E.A. Defects in Mitochondrial Protein Synthesis and Respiratory Chain Activity Segregate with the TRNA(Leu(UUR)) Mutation Associated with Mitochondrial Myopathy, Encephalopathy, Lactic Acidosis, and Strokelike Episodes. Mol. Cell. Biol. 1992, 12, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Schon, E.A.; Koga, Y.; Davidson, M.; Moraes, C.T.; King, M.P. The Mitochondrial TRNA(Leu)(UUR)) Mutation in MELAS: A Model for Pathogenesis. Biochim. Biophys. Acta 1992, 1101, 206–209. [Google Scholar] [PubMed]
- Koga, Y.; Davidson, M.; Schon, E.A.; King, M.P. Fine Mapping of Mitochondrial RNAs Derived from the MtDNA Region Containing a Point Mutation Associated with MELAS. Nucleic Acids Res. 1993, 21, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Temperley, R.J.; Wydro, M.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M. Human Mitochondrial MRNAs--like Members of All Families, Similar but Different. Biochim. Biophys. Acta 2010, 1797, 1081–1085. [Google Scholar] [CrossRef]
- Maniura-Weber, K.; Helm, M.; Engemann, K.; Eckertz, S.; Möllers, M.; Schauen, M.; Hayrapetyan, A.; von Kleist-Retzow, J.-C.; Lightowlers, R.N.; Bindoff, L.A.; et al. Molecular Dysfunction Associated with the Human Mitochondrial 3302A>G Mutation in the MTTL1 (Mt-TRNALeu(UUR)) Gene. Nucleic Acids Res. 2006, 34, 6404–6415. [Google Scholar] [CrossRef]
- Bindoff, L.A.; Howell, N.; Poulton, J.; McCullough, D.A.; Morten, K.J.; Lightowlers, R.N.; Turnbull, D.M.; Weber, K. Abnormal RNA Processing Associated with a Novel TRNA Mutation in Mitochondrial DNA. A Potential Disease Mechanism. J. Biol. Chem. 1993, 268, 19559–19564. [Google Scholar] [CrossRef]
- Chomyn, A.; Enriquez, J.A.; Micol, V.; Fernandez-Silva, P.; Attardi, G. The Mitochondrial Myopathy, Encephalopathy, Lactic Acidosis, and Stroke-like Episode Syndrome-Associated Human Mitochondrial TRNA Leu(UUR) Mutation Causes Aminoacylation Deficiency and Concomitant Reduced Association of MRNA with Ribosomes. J. Biol. Chem. 2000, 275, 19198–19209. [Google Scholar] [CrossRef]
- Koga, A.; Koga, Y.; Akita, Y.; Fukiyama, R.; Ueki, I.; Yatsuga, S.; Matsuishi, T. Increased Mitochondrial Processing Intermediates Associated with Three TRNALeu(UUR) Gene Mutations. Neuromuscul. Disord. 2003, 13, 259–262. [Google Scholar] [CrossRef]
- Tani, H.; Ishikawa, K.; Tamashiro, H.; Ogasawara, E.; Yasukawa, T.; Matsuda, S.; Shimizu, A.; Kang, D.; Hayashi, J.-I.; Wei, F.-Y.; et al. Aberrant RNA Processing Contributes to the Pathogenesis of Mitochondrial Diseases in Trans-Mitochondrial Mouse Model Carrying Mitochondrial TRNALeu(UUR) with a Pathogenic A2748G Mutation. Nucleic Acids Res. 2022, 50, 9382–9396. [Google Scholar] [CrossRef] [PubMed]
- Rackham, O.; Davies, S.M.K.; Shearwood, A.-M.J.; Hamilton, K.L.; Whelan, J.; Filipovska, A. Pentatricopeptide Repeat Domain Protein 1 Lowers the Levels of Mitochondrial Leucine TRNAs in Cells. Nucleic Acids Res. 2009, 37, 5859–5867. [Google Scholar] [CrossRef] [PubMed]
- Perks, K.L.; Rossetti, G.; Kuznetsova, I.; Hughes, L.A.; Ermer, J.A.; Ferreira, N.; Busch, J.D.; Rudler, D.L.; Spahr, H.; Schöndorf, T.; et al. PTCD1 Is Required for 16S RRNA Maturation Complex Stability and Mitochondrial Ribosome Assembly. Cell Rep. 2018, 23, 127–142. [Google Scholar] [CrossRef]
- Capriglia, F.; Rizzo, F.; Petrosillo, G.; Morea, V.; d’Amati, G.; Cantatore, P.; Roberti, M.; Loguercio Polosa, P.; Bruni, F. Exploring the Ability of LARS2 Carboxy-Terminal Domain in Rescuing the MELAS Phenotype. Life 2021, 11, 674. [Google Scholar] [CrossRef] [PubMed]
- Loguercio Polosa, P.; Capriglia, F.; Bruni, F. Molecular Investigation of Mitochondrial RNA19 Role in the Pathogenesis of MELAS Disease. Life 2023, 13, 1863. [Google Scholar] [CrossRef] [PubMed]
- Villegas, J.; Burzio, V.; Villota, C.; Landerer, E.; Martinez, R.; Santander, M.; Martinez, R.; Pinto, R.; Vera, M.I.; Boccardo, E.; et al. Expression of a Novel Non-Coding Mitochondrial RNA in Human Proliferating Cells. Nucleic Acids Res. 2007, 35, 7336–7347. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Rasmusson, A.G.; Van Aken, O. Plant Mitochondria-Past, Present and Future. Plant J. 2021, 108, 912–959. [Google Scholar] [CrossRef]
- Herai, R.H.; Negraes, P.D.; Muotri, A.R. Evidence of Nuclei-Encoded Spliceosome Mediating Splicing of Mitochondrial RNA. Hum. Mol. Genet. 2017, 26, 2472–2479. [Google Scholar] [CrossRef]
- Rodrigues, K.S.; Petroski, L.P.; Utumi, P.H.; Ferrasa, A.; Herai, R.H. IARA: A Complete and Curated Atlas of the Biogenesis of Spliceosome Machinery during RNA Splicing. Life Sci. Alliance 2023, 6, e202201593. [Google Scholar] [CrossRef] [PubMed]
- Burzio, V.A.; Villota, C.; Villegas, J.; Landerer, E.; Boccardo, E.; Villa, L.L.; Martínez, R.; Lopez, C.; Gaete, F.; Toro, V.; et al. Expression of a Family of Noncoding Mitochondrial RNAs Distinguishes Normal from Cancer Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 9430–9434. [Google Scholar] [CrossRef] [PubMed]
- Landerer, E.; Villegas, J.; Burzio, V.A.; Oliveira, L.; Villota, C.; Lopez, C.; Restovic, F.; Martinez, R.; Castillo, O.; Burzio, L.O. Nuclear Localization of the Mitochondrial NcRNAs in Normal and Cancer Cells. Cell Oncol. 2011, 34, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Rackham, O.; Shearwood, A.-M.J.; Mercer, T.R.; Davies, S.M.K.; Mattick, J.S.; Filipovska, A. Long Noncoding RNAs Are Generated from the Mitochondrial Genome and Regulated by Nuclear-Encoded Proteins. RNA 2011, 17, 2085–2093. [Google Scholar] [CrossRef] [PubMed]
- Ojala, D.; Attardi, G. Identification of Discrete Polyadenylate-Containing RNA Components Transcribed from HeLa Cell Mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1974, 71, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.D.; Clayton, D.A. Precise Identification of Individual Promoters for Transcription of Each Strand of Human Mitochondrial DNA. Cell 1984, 36, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Ojala, D.; Crews, S.; Montoya, J.; Gelfand, R.; Attardi, G. A Small Polyadenylated RNA (7S RNA), Containing a Putative Ribosome Attachment Site, Maps near the Origin of Human Mitochondrial DNA Replication. J. Mol. Biol. 1981, 150, 303–314. [Google Scholar] [CrossRef]
- Jørgensen, T.E.; Bakke, I.; Ursvik, A.; Andreassen, M.; Moum, T.; Johansen, S.D. An Evolutionary Preserved Intergenic Spacer in Gadiform Mitogenomes Generates a Long Noncoding RNA. BMC Evol. Biol. 2014, 14, 182. [Google Scholar] [CrossRef]
- Jemt, E.; Persson, Ö.; Shi, Y.; Mehmedovic, M.; Uhler, J.P.; Dávila López, M.; Freyer, C.; Gustafsson, C.M.; Samuelsson, T.; Falkenberg, M. Regulation of DNA Replication at the End of the Mitochondrial D-Loop Involves the Helicase TWINKLE and a Conserved Sequence Element. Nucleic Acids Res. 2015, 43, 9262–9275. [Google Scholar] [CrossRef]
- Falkenberg, M.; Larsson, N.-G.; Gustafsson, C.M. DNA Replication and Transcription in Mammalian Mitochondria. Annu. Rev. Biochem. 2007, 76, 679–699. [Google Scholar] [CrossRef]
- Misic, J.; Milenkovic, D.; Al-Behadili, A.; Xie, X.; Jiang, M.; Jiang, S.; Filograna, R.; Koolmeister, C.; Siira, S.J.; Jenninger, L.; et al. Mammalian RNase H1 Directs RNA Primer Formation for MtDNA Replication Initiation and Is Also Necessary for MtDNA Replication Completion. Nucleic Acids Res. 2022, 50, 8749–8766. [Google Scholar] [CrossRef]
- Reyes, A.; Rusecka, J.; Tońska, K.; Zeviani, M. RNase H1 Regulates Mitochondrial Transcription and Translation via the Degradation of 7S RNA. Front. Genet. 2019, 10, 1393. [Google Scholar] [CrossRef]
- Cantatore, P.; Polosa, P.L.; Mustich, A.; Petruzzella, V.; Gadaleta, M.N. Faithful and Highly Efficient RNA Synthesis in Isolated Mitochondria from Rat Liver. Curr. Genet. 1988, 14, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Xie, X.; Das, H.; Tan, B.G.; Shi, Y.; Al-Behadili, A.; Peter, B.; Motori, E.; Valenzuela, S.; Posse, V.; et al. Non-Coding 7S RNA Inhibits Transcription via Mitochondrial RNA Polymerase Dimerization. Cell 2022, 185, 2309–2323. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Memon, A.A.; Hedelius, A.; Grundberg, A.; Elf, J.L.; Svensson, P.J.; Sundquist, J.; Sundquist, K. Association of Circulating Long Noncoding 7S RNA with Deep Vein Thrombosis. Semin. Thromb. Hemost. 2023, 49, 702–708. [Google Scholar] [CrossRef]
- Gao, S.; Tian, X.; Chang, H.; Sun, Y.; Wu, Z.; Cheng, Z.; Dong, P.; Zhao, Q.; Ruan, J.; Bu, W. Two Novel LncRNAs Discovered in Human Mitochondrial DNA Using PacBio Full-Length Transcriptome Data. Mitochondrion 2018, 38, 41–47. [Google Scholar] [CrossRef]
- Taft, R.J.; Glazov, E.A.; Cloonan, N.; Simons, C.; Stephen, S.; Faulkner, G.J.; Lassmann, T.; Forrest, A.R.R.; Grimmond, S.M.; Schroder, K.; et al. Tiny RNAs Associated with Transcription Start Sites in Animals. Nat. Genet. 2009, 41, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Bai, R.; Yang, W.; Miao, H.; Li, Y.; Dai, H.; Li, L.; Zhao, Y.; Song, X. The Mitochondrial-Derived LncRNA MDL1 Mediates a Mitochondria-to-Nucleus Retrograde Regulation by Inhibiting the Nuclear Translocation of P53. MedComm-Oncology 2022, 1, e15. [Google Scholar] [CrossRef]
- Trinh, D.L.N.; Elwi, A.N.; Kim, S.-W. Direct Interaction between P53 and Tid1 Proteins Affects P53 Mitochondrial Localization and Apoptosis. Oncotarget 2010, 1, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.Y.; Trinh, D.L.N.; Zajchowski, L.D.; Lee, B.; Elwi, A.N.; Kim, S.-W. Tid1 Is a New Regulator of P53 Mitochondrial Translocation and Apoptosis in Cancer. Oncogene 2010, 29, 1155–1166. [Google Scholar] [CrossRef]
- Farrelly, F.; Butow, R.A. Rearranged Mitochondrial Genes in the Yeast Nuclear Genome. Nature 1983, 301, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Wakasugi, S.; Tsuzuki, T.; Nomiyama, H.; Shimada, K.; Miyata, T. Mitochondrial DNA-like Sequences in the Human Nuclear Genome: Characterization and Implications in the Evolution of Mitochondrial DNA. J. Mol. Biol. 1985, 186, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Gellissen, G.; Bradfield, J.Y.; White, B.N.; Wyatt, G.R. Mitochondrial DNA Sequences in the Nuclear Genome of a Locust. Nature 1983, 301, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Hadler, H.I.; Dimitrijevic, B.; Mahalingam, R. Mitochondrial DNA and Nuclear DNA from Normal Rat Liver Have a Common Sequence. Proc. Natl. Acad. Sci. USA 1983, 80, 6495–6499. [Google Scholar] [CrossRef] [PubMed]
- van den Boogaart, P.; Samallo, J.; Agsteribbe, E. Similar Genes for a Mitochondrial ATPase Subunit in the Nuclear and Mitochondrial Genomes of Neurospora Crassa. Nature 1982, 298, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Moreira, J.D.; Smith, K.K.; Fetterman, J.L. The Mighty NUMT: Mitochondrial DNA Flexing Its Code in the Nuclear Genome. Biomolecules 2023, 13, 753. [Google Scholar] [CrossRef] [PubMed]
- Kumarswamy, R.; Bauters, C.; Volkmann, I.; Maury, F.; Fetisch, J.; Holzmann, A.; Lemesle, G.; de Groote, P.; Pinet, F.; Thum, T. Circulating Long Noncoding RNA, LIPCAR, Predicts Survival in Patients with Heart Failure. Circ. Res. 2014, 114, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gao, W.; Long, Q.-Q.; Zhang, J.; Li, Y.-F.; Liu, D.-C.; Yan, J.-J.; Yang, Z.-J.; Wang, L.-S. Increased Plasma Levels of LncRNA H19 and LIPCAR Are Associated with Increased Risk of Coronary Artery Disease in a Chinese Population. Sci. Rep. 2017, 7, 7491. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.-F.; Yang, X.-C.; Xu, L.; Li, W.-M.; Xia, K.; Zhang, D.-P.; Wu, R.-N.; Gan, T. Circulating Long Noncoding RNA LIPCAR Acts as a Novel Biomarker in Patients with ST-Segment Elevation Myocardial Infarction. Med. Sci. Monit. 2018, 24, 5064–5070. [Google Scholar] [CrossRef]
- Santer, L.; López, B.; Ravassa, S.; Baer, C.; Riedel, I.; Chatterjee, S.; Moreno, M.U.; González, A.; Querejeta, R.; Pinet, F.; et al. Circulating Long Noncoding RNA LIPCAR Predicts Heart Failure Outcomes in Patients Without Chronic Kidney Disease. Hypertension 2019, 73, 820–828. [Google Scholar] [CrossRef]
- Cao, M.; Luo, H.; Li, D.; Wang, S.; Xuan, L.; Sun, L. Research Advances on Circulating Long Noncoding RNAs as Biomarkers of Cardiovascular Diseases. Int. J. Cardiol. 2022, 353, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Meessen, J.M.T.A.; Bär, C.; di Dona, F.M.; Staszewsky, L.I.; Di Giulio, P.; Di Tano, G.; Costa, A.; Leonardy, J.; Novelli, D.; Nicolis, E.B.; et al. LIPCAR Is Increased in Chronic Symptomatic HF Patients. A Sub-Study of the GISSI-HF Trial. Clin. Chem. 2021, 67, 1721–1731. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Song, T.; Zhao, Y.; Zhao, J.; Wang, X.; Fu, X. Long Non-Coding RNA LICPAR Regulates Atrial Fibrosis via TGF-β/Smad Pathway in Atrial Fibrillation. Tissue Cell 2020, 67, 101440. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhang, Y.; Wang, M.; Wang, L.; Zhang, W.; Ge, Z.-R. Circulating LIPCAR Is a Potential Biomarker of Heart Failure in Patients Post-Acute Myocardial Infarction. Exp. Biol. Med. 2021, 246, 2589–2594. [Google Scholar] [CrossRef] [PubMed]
- Barbalata, T.; Niculescu, L.S.; Stancu, C.S.; Pinet, F.; Sima, A.V. Elevated Levels of Circulating LncRNAs LIPCAR and MALAT1 Predict an Unfavorable Outcome in Acute Coronary Syndrome Patients. Int. J. Mol. Sci. 2023, 24, 12076. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Lin, W.; Feng, Y.; Huang, C.; Yan, Y.; Guo, W.; Zhang, H.; Lei, Z.; Lu, Q.; Liu, P.; et al. Plasma LncRNA LIPCAR Expression Levels Associated with Neurological Impairment and Stroke Subtypes in Patients with Acute Cerebral Infarction: A Prospective Observational Study with a Control Group. Neurol. Ther. 2023, 12, 1385–1398. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zhang, Q.; Mao, J.; Zhang, J.; Li, L. The Roles of LncRNA in Myocardial Infarction: Molecular Mechanisms, Diagnosis Biomarkers, and Therapeutic Perspectives. Front. Cell Dev. Biol. 2021, 9, 680713. [Google Scholar] [CrossRef]
- Wang, X.; Li, D.; Chen, H.; Wei, X.; Xu, X. Expression of Long Noncoding RNA LIPCAR Promotes Cell Proliferation, Cell Migration, and Change in Phenotype of Vascular Smooth Muscle Cells. Med. Sci. Monit. 2019, 25, 7645–7651. [Google Scholar] [CrossRef]
- Bongolo, C.C.; Thokerunga, E.; Fidele, N.B.; Souraka, T.D.M.; Kisembo, P.; Rugera, S.P.; Worley, P.F.; Tu, J.-C. Upregulation of the Long Non-Coding RNA, LIPCAR Promotes Proliferation, Migration, and Metastasis of Hepatocellular Carcinoma. Cancer Biomark. 2022, 35, 245–256. [Google Scholar] [CrossRef]
- Young, P.G.; Attardi, G. Characterization of Double-Stranded RNA from HeLa Cell Mitochondria. Biochem. Biophys. Res. Commun. 1975, 65, 1201–1207. [Google Scholar] [CrossRef]
- Kim, S.; Yoon, J.; Lee, K.; Kim, Y. Analysis of Mitochondrial Double-Stranded RNAs in Human Cells. STAR Protoc. 2023, 4, 102007. [Google Scholar] [CrossRef]
- Dhir, A.; Dhir, S.; Borowski, L.S.; Jimenez, L.; Teitell, M.; Rötig, A.; Crow, Y.J.; Rice, G.I.; Duffy, D.; Tamby, C.; et al. Mitochondrial Double-Stranded RNA Triggers Antiviral Signalling in Humans. Nature 2018, 560, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, J.; Kim, S.; Kim, M.; Kang, M.-G.; Kwak, C.; Kang, M.; Kim, B.; Rhee, H.-W.; Kim, V.N. PKR Senses Nuclear and Mitochondrial Signals by Interacting with Endogenous Double-Stranded RNAs. Mol. Cell 2018, 71, 1051–1063.e6. [Google Scholar] [CrossRef] [PubMed]
- Grochowska, J.; Czerwinska, J.; Borowski, L.S.; Szczesny, R.J. Mitochondrial RNA, a New Trigger of the Innate Immune System. WIREs RNA 2022, 13, e1690. [Google Scholar] [CrossRef]
- Borowski, L.S.; Dziembowski, A.; Hejnowicz, M.S.; Stepien, P.P.; Szczesny, R.J. Human Mitochondrial RNA Decay Mediated by PNPase–HSuv3 Complex Takes Place in Distinct Foci. Nucleic Acids Res. 2013, 41, 1223–1240. [Google Scholar] [CrossRef] [PubMed]
- Bruni, F.; Gramegna, P.; Oliveira, J.M.A.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.A. REXO2 Is an Oligoribonuclease Active in Human Mitochondria. PLoS ONE 2013, 8, e64670. [Google Scholar] [CrossRef]
- Szewczyk, M.; Malik, D.; Borowski, L.S.; Czarnomska, S.D.; Kotrys, A.V.; Klosowska-Kosicka, K.; Nowotny, M.; Szczesny, R.J. Human REXO2 Controls Short Mitochondrial RNAs Generated by MtRNA Processing and Decay Machinery to Prevent Accumulation of Double-Stranded RNA. Nucleic Acids Res. 2020, 48, 5572–5590. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhong, Y.; Wang, X.; Shen, J.; An, W. Advances in Circular RNA and Its Applications. Int. J. Med. Sci. 2022, 19, 975–985. [Google Scholar] [CrossRef]
- Liu, C.-X.; Chen, L.-L. Circular RNAs: Characterization, Cellular Roles, and Applications. Cell 2022, 185, 2016–2034. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Li, J.; Hu, S.; Deng, Y.; Yin, H.; Bao, X.; Zhang, Q.C.; Wang, G.; Wang, B.; et al. Identification of MecciRNAs and Their Roles in the Mitochondrial Entry of Proteins. Sci. China Life Sci. 2020, 63, 1429–1449. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, J.; Deng, H.; Ma, R.; Liao, J.-Y.; Liang, H.; Hu, J.; Li, J.; Guo, Z.; Cai, J.; et al. Targeting Mitochondria-Located CircRNA SCAR Alleviates NASH via Reducing MROS Output. Cell 2020, 183, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Sun, H.; Wang, C.; Liu, W.; Liu, M.; Zhu, Y.; Xu, W.; Jin, H.; Li, J. Mitochondrial Genome-Derived CircRNA Mc-COX2 Functions as an Oncogene in Chronic Lymphocytic Leukemia. Mol. Ther. Nucleic Acids 2020, 20, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.J.; Amode, M.R.; Aneja, A.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Becker, A.; Bennett, R.; Berry, A.; Bhai, J.; et al. Ensembl 2023. Nucleic Acids Res. 2023, 51, D933–D941. [Google Scholar] [CrossRef] [PubMed]
- Elson, J.L.; Smith, P.M.; Vila-Sanjurjo, A. Heterologous Inferential Analysis (HIA) as a Method to Understand the Role of Mitochondrial RRNA Mutations in Pathogenesis. Methods Mol. Biol. 2015, 1264, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Elson, J.L.; Greaves, L.C.; Wortmann, S.B.; Rodenburg, R.J.T.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.A.; Taylor, R.W.; Vila-Sanjurjo, A. The Role of the Mitochondrial Ribosome in Human Disease: Searching for Mutations in 12S Mitochondrial RRNA with High Disruptive Potential. Hum. Mol. Genet. 2014, 23, 949–967. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Elson, J.L.; Smith, P.M.; Greaves, L.C.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.A.; Taylor, R.W.; Vila-Sanjurjo, A. The Presence of Highly Disruptive 16S RRNA Mutations in Clinical Samples Indicates a Wider Role for Mutations of the Mitochondrial Ribosome in Human Disease. Mitochondrion 2015, 25, 17. [Google Scholar] [CrossRef] [PubMed]
- Vila-Sanjurjo, A.; Mallo, N.; Elson, J.L.; Smith, P.M.; Blakely, E.L.; Taylor, R.W. Structural Analysis of Mitochondrial RRNA Gene Variants Identified in Patients with Deafness. Front. Physiol. 2023, 14, 1163496. [Google Scholar] [CrossRef]
- Vidaurre, S.; Fitzpatrick, C.; Burzio, V.A.; Briones, M.; Villota, C.; Villegas, J.; Echenique, J.; Oliveira-Cruz, L.; Araya, M.; Borgna, V.; et al. Down-Regulation of the Antisense Mitochondrial Non-Coding RNAs (NcRNAs) Is a Unique Vulnerability of Cancer Cells and a Potential Target for Cancer Therapy. J. Biol. Chem. 2014, 289, 27182–27198. [Google Scholar] [CrossRef]
- Villota, C.; Campos, A.; Vidaurre, S.; Oliveira-Cruz, L.; Boccardo, E.; Burzio, V.A.; Varas, M.; Villegas, J.; Villa, L.L.; Valenzuela, P.D.T.; et al. Expression of Mitochondrial Non-Coding RNAs (NcRNAs) Is Modulated by High Risk Human Papillomavirus (HPV) Oncogenes *. J. Biol. Chem. 2012, 287, 21303–21315. [Google Scholar] [CrossRef]
- Fitzpatrick, C.; Bendek, M.F.; Briones, M.; Farfán, N.; Silva, V.A.; Nardocci, G.; Montecino, M.; Boland, A.; Deleuze, J.-F.; Villegas, J.; et al. Mitochondrial NcRNA Targeting Induces Cell Cycle Arrest and Tumor Growth Inhibition of MDA-MB-231 Breast Cancer Cells through Reduction of Key Cell Cycle Progression Factors. Cell Death Dis. 2019, 10, 423. [Google Scholar] [CrossRef]
- Borgna, V.; Lobos-González, L.; Guevara, F.; Landerer, E.; Bendek, M.; Ávila, R.; Silva, V.; Villota, C.; Araya, M.; Rivas, A.; et al. Targeting Antisense Mitochondrial Noncoding RNAs Induces Bladder Cancer Cell Death and Inhibition of Tumor Growth through Reduction of Survival and Invasion Factors. J. Cancer 2020, 11, 1780–1791. [Google Scholar] [CrossRef]
- Lobos-González, L.; Bustos, R.; Campos, A.; Silva, V.; Silva, V.; Jeldes, E.; Salomon, C.; Varas-Godoy, M.; Cáceres-Verschae, A.; Duran, E.; et al. Exosomes Released upon Mitochondrial ASncmtRNA Knockdown Reduce Tumorigenic Properties of Malignant Breast Cancer Cells. Sci. Rep. 2020, 10, 343. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Fenster, R.J.; Pineda, S.S.; Gibbs, W.S.; Mohammadi, S.; Davila-Velderrain, J.; Garcia, F.J.; Therrien, M.; Novis, H.S.; Gao, F.; et al. Cell Type-Specific Transcriptomics Reveals That Mutant Huntingtin Leads to Mitochondrial RNA Release and Neuronal Innate Immune Activation. Neuron 2020, 107, 891–908.e8. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Shim, Y.-R.; Seo, W.; Kim, M.-H.; Choi, W.-M.; Kim, H.-H.; Kim, Y.E.; Yang, K.; Ryu, T.; Jeong, J.-M.; et al. Mitochondrial Double-Stranded RNA in Exosome Promotes Interleukin-17 Production Through Toll-Like Receptor 3 in Alcohol-Associated Liver Injury. Hepatology 2020, 72, 609–625. [Google Scholar] [CrossRef]
- Peel, A.L.; Rao, R.V.; Cottrell, B.A.; Hayden, M.R.; Ellerby, L.M.; Bredesen, D.E. Double-Stranded RNA-Dependent Protein Kinase, PKR, Binds Preferentially to Huntington’s Disease (HD) Transcripts and Is Activated in HD Tissue. Hum. Mol. Genet. 2001, 10, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Lee, M.; Ali, A.A.; Oh, Y.R.; Choi, Y.S.; Kim, S.; Lee, N.; Jang, S.G.; Park, S.; Chung, J.-H.; et al. Mitochondrial Double-Stranded RNAs as a Pivotal Mediator in the Pathogenesis of Sjӧgren’s Syndrome. Mol. Ther. Nucleic Acids 2022, 30, 257–269. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, M.; Wang, W.; Qu, S.; Liu, M.; Rong, W.; Yang, W.; Liang, H.; Zeng, C.; Zhu, X.; et al. Polynucleotide Phosphorylase Protects against Renal Tubular Injury via Blocking Mt-DsRNA-PKR-EIF2α Axis. Nat. Commun. 2023, 14, 1223. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, K.; Choi, Y.S.; Ku, J.; Kim, H.; Kharbash, R.; Yoon, J.; Lee, Y.S.; Kim, J.-H.; Lee, Y.J.; et al. Mitochondrial Double-Stranded RNAs Govern the Stress Response in Chondrocytes to Promote Osteoarthritis Development. Cell Rep. 2022, 40, 111178. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Jiao, C.; Ma, C.; Zhang, Y.; Hao, X.; Zhou, G.; Fu, J.; Qiu, X.; Li, H.; Yang, W.; et al. CircMTND5 Participates in Renal Mitochondrial Injury and Fibrosis by Sponging MIR6812 in Lupus Nephritis. Oxidative Med. Cell. Longev. 2022, 2022, e2769487. [Google Scholar] [CrossRef]
- Vendramin, R.; Verheyden, Y.; Ishikawa, H.; Goedert, L.; Nicolas, E.; Saraf, K.; Armaos, A.; Delli Ponti, R.; Izumikawa, K.; Mestdagh, P.; et al. SAMMSON Fosters Cancer Cell Fitness by Concertedly Enhancing Mitochondrial and Cytosolic Translation. Nat. Struct. Mol. Biol. 2018, 25, 1035–1046. [Google Scholar] [CrossRef]
- Leucci, E.; Vendramin, R.; Spinazzi, M.; Laurette, P.; Fiers, M.; Wouters, J.; Radaelli, E.; Eyckerman, S.; Leonelli, C.; Vanderheyden, K.; et al. Melanoma Addiction to the Long Non-Coding RNA SAMMSON. Nature 2016, 531, 518–522. [Google Scholar] [CrossRef]
- Arun, G.; Aggarwal, D.; Spector, D.L. MALAT1 Long Non-Coding RNA: Functional Implications. Noncoding RNA 2020, 6, 22. [Google Scholar] [CrossRef]
- Zhao, Y.; Cui, J.; Hu, J.; Hoffman, A.R. Effect of MALAT1 in the Crosstalk between Nucleus and Mitochondria on Mitochondrial Reprogramming in Hepatocellular Carcinoma Cells. JCO 2019, 37, e14711. [Google Scholar] [CrossRef]
- Chernega, T.; Choi, J.; Salmena, L.; Andreazza, A.C. Mitochondrion-Targeted RNA Therapies as a Potential Treatment Strategy for Mitochondrial Diseases. Mol. Ther. Nucleic Acids 2022, 30, 359–377. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mt-lncRNA | Length (nt) | Function(s) | Localisation(s) |
|---|---|---|---|
| 16S mt-rRNA | 1559 |
| mitochondria |
| 12S mt-rRNA | 954 |
| mitochondria |
| RNA19 | 2591 |
| mitochondria |
| SncmtRNA | 2374 |
| nucleus, nucleolus and cytoplasm |
| ASncmtRNA-1 | 1866 |
| |
| ASncmtRNA-2 | 2104 | ||
| lncND5 lncND6 lncCyt b | 1806 753 1212 |
| mitochondria |
| 7S RNA | 188 |
| mitochondria |
| hsaMDL1 hsaMDL1AS | 381 384 |
| mitochondria and nucleus |
| LIPCAR | 781 |
| plasma |
| mt-dsRNAs | variable |
| ubiquitous |
| mecciND1 | 144 |
| mitochondria and cytoplasm |
| mecciND5 | 154 |
| |
| SCAR, mc-COX2 | 396 |
| mitochondria and exosomes |
| circMTND5 | 346 |
| mitochondria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruni, F. Human mtDNA-Encoded Long ncRNAs: Knotty Molecules and Complex Functions. Int. J. Mol. Sci. 2024, 25, 1502. https://doi.org/10.3390/ijms25031502
Bruni F. Human mtDNA-Encoded Long ncRNAs: Knotty Molecules and Complex Functions. International Journal of Molecular Sciences. 2024; 25(3):1502. https://doi.org/10.3390/ijms25031502
Chicago/Turabian StyleBruni, Francesco. 2024. "Human mtDNA-Encoded Long ncRNAs: Knotty Molecules and Complex Functions" International Journal of Molecular Sciences 25, no. 3: 1502. https://doi.org/10.3390/ijms25031502
APA StyleBruni, F. (2024). Human mtDNA-Encoded Long ncRNAs: Knotty Molecules and Complex Functions. International Journal of Molecular Sciences, 25(3), 1502. https://doi.org/10.3390/ijms25031502