

Metabonomics and Transcriptomics Analyses Reveal the Development Process of the Auditory System in the Embryonic Development Period of the Small Yellow Croaker under Background Noise


Abstract
1. Introduction
2. Results
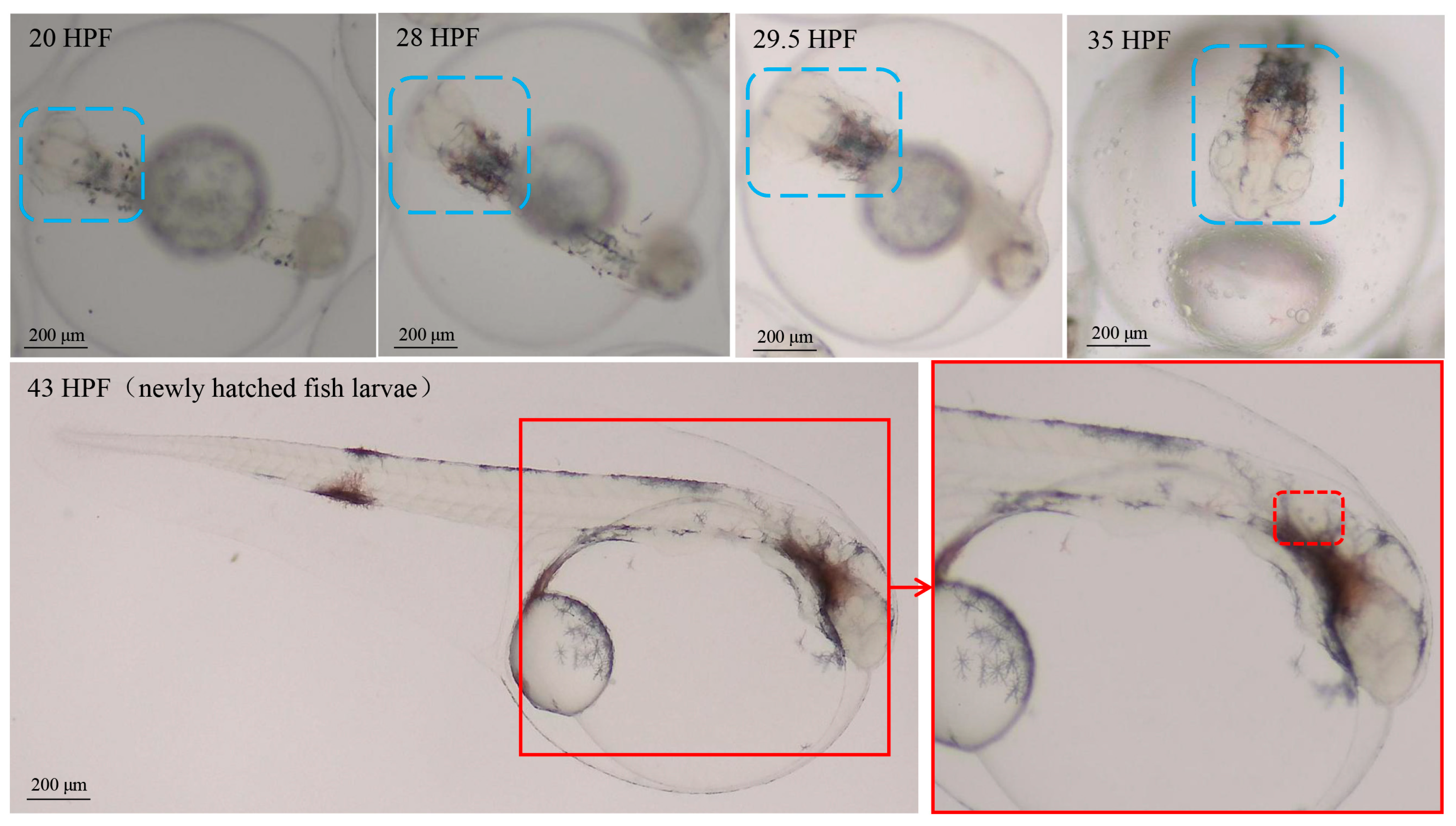
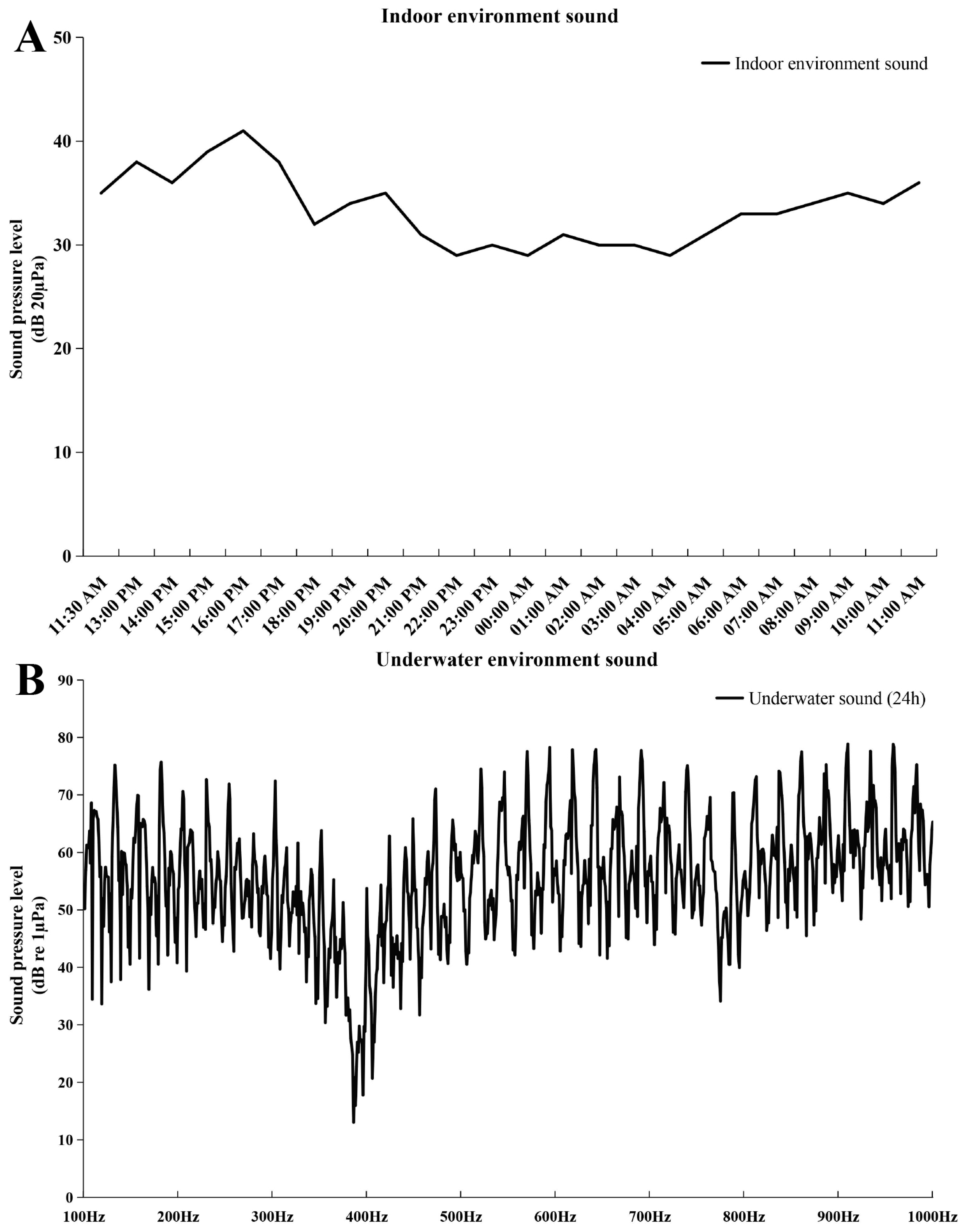
2.1. The Results of Embryonic Staging and Background Noise Detection
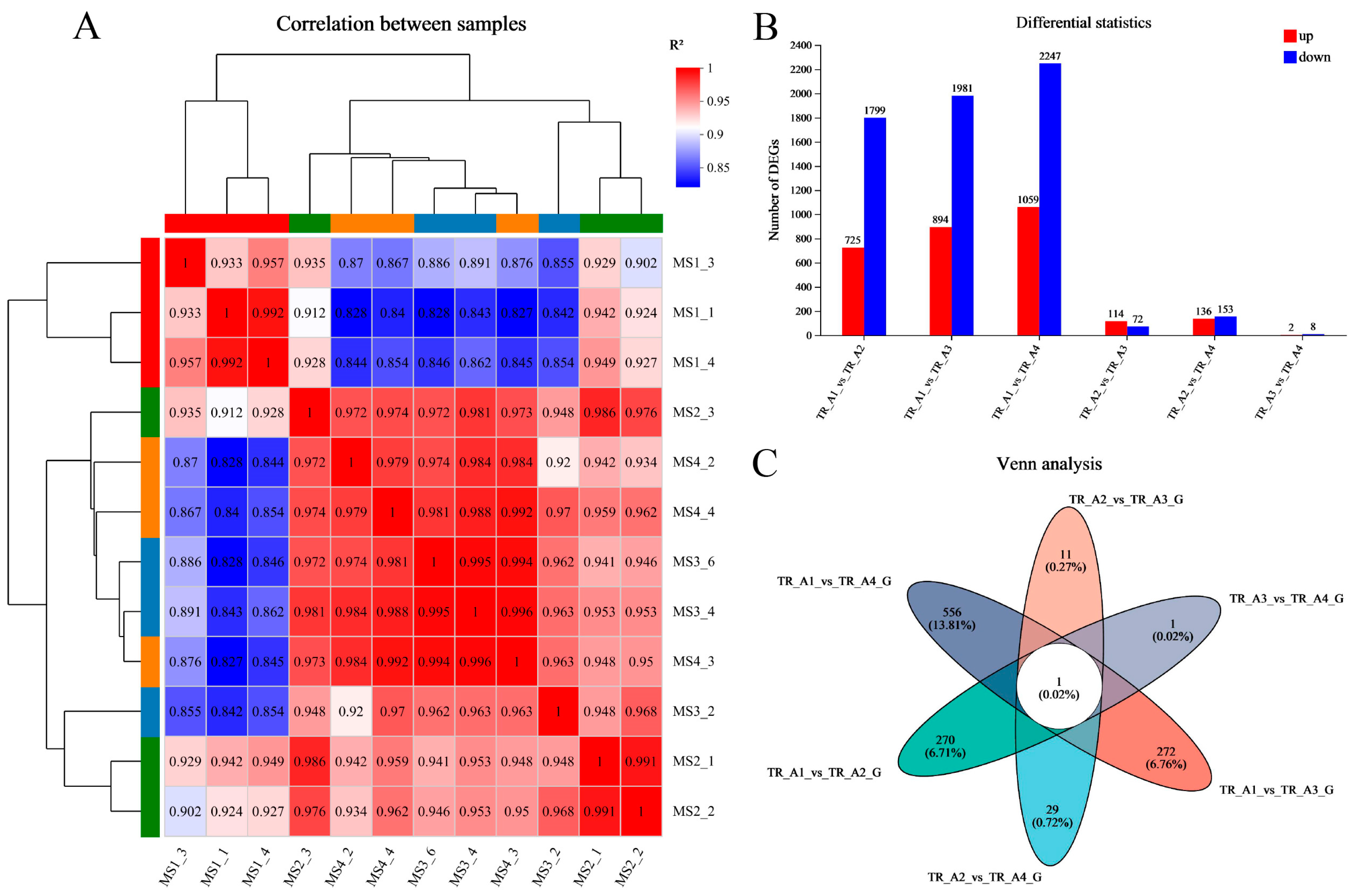
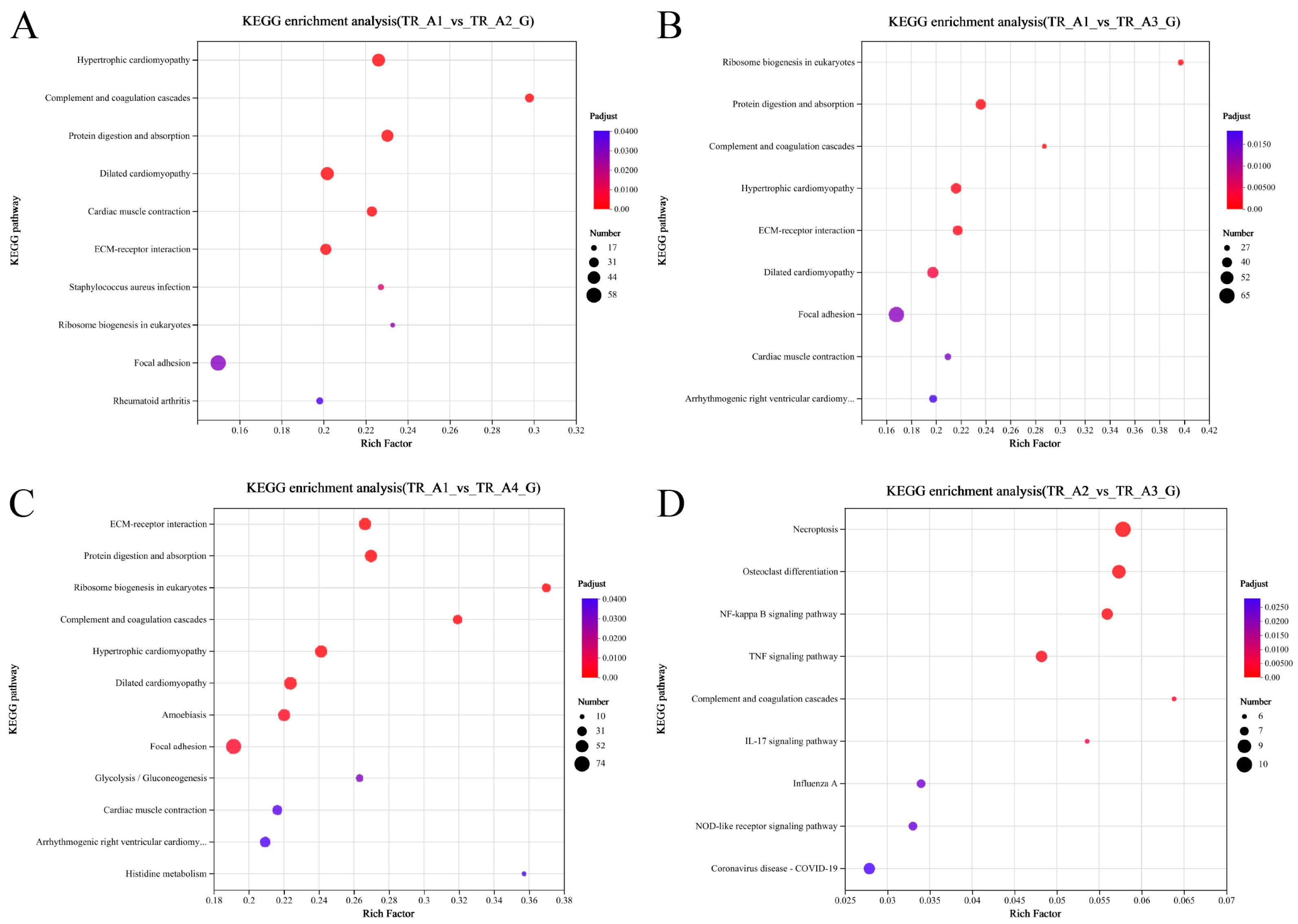
2.2. Analysis of Transcriptomic Data
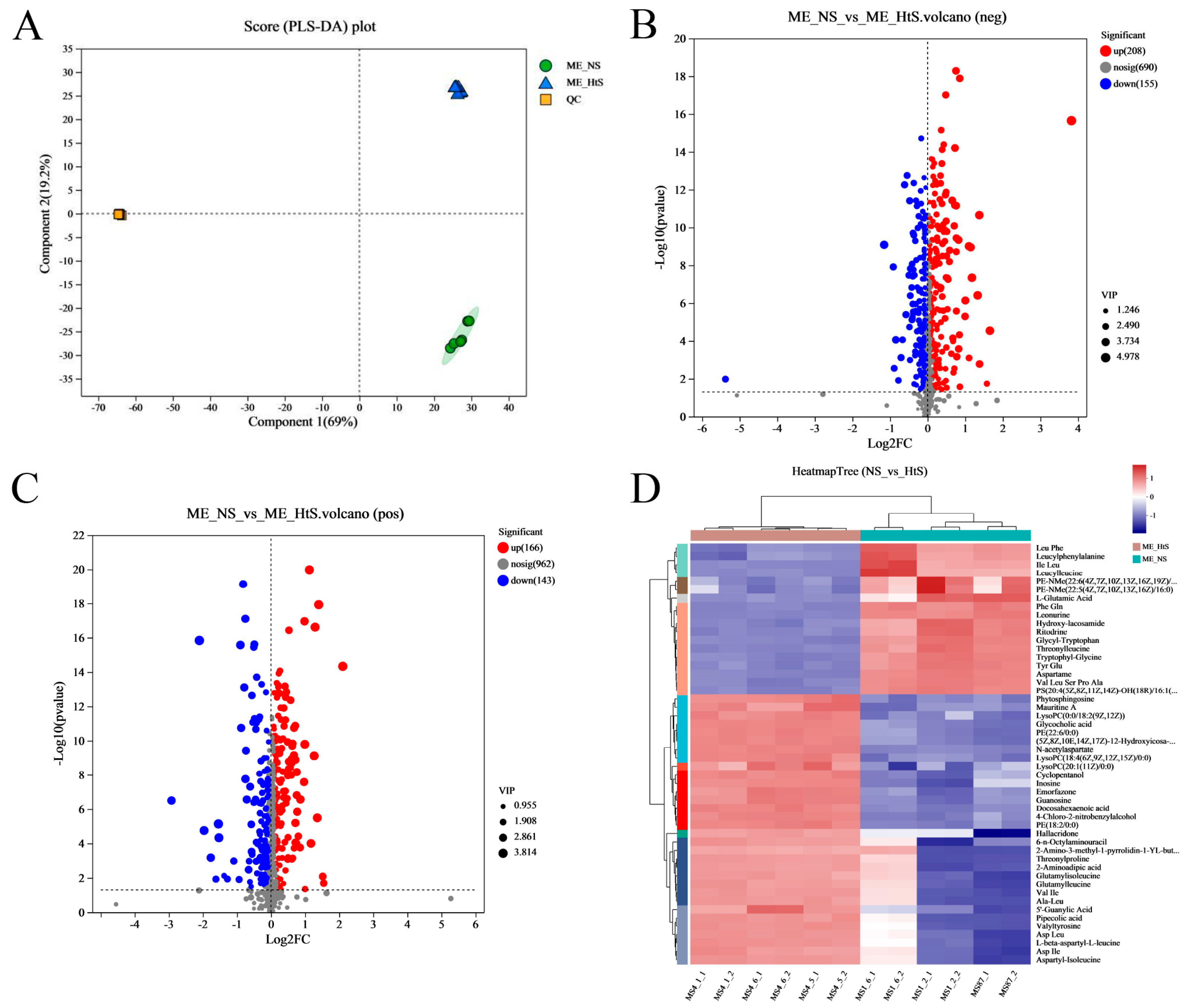
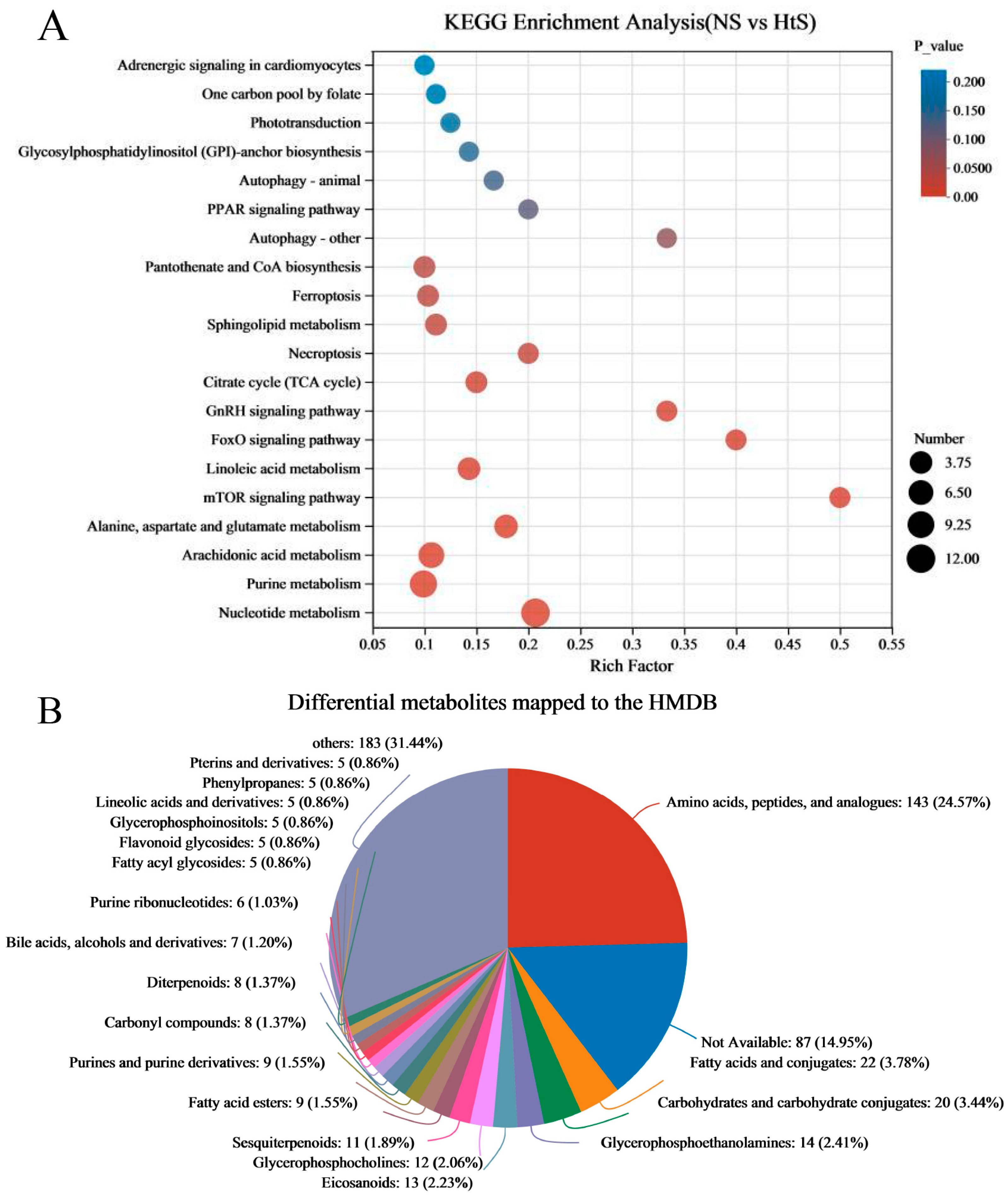
2.3. Analysis of Metabonomic Data
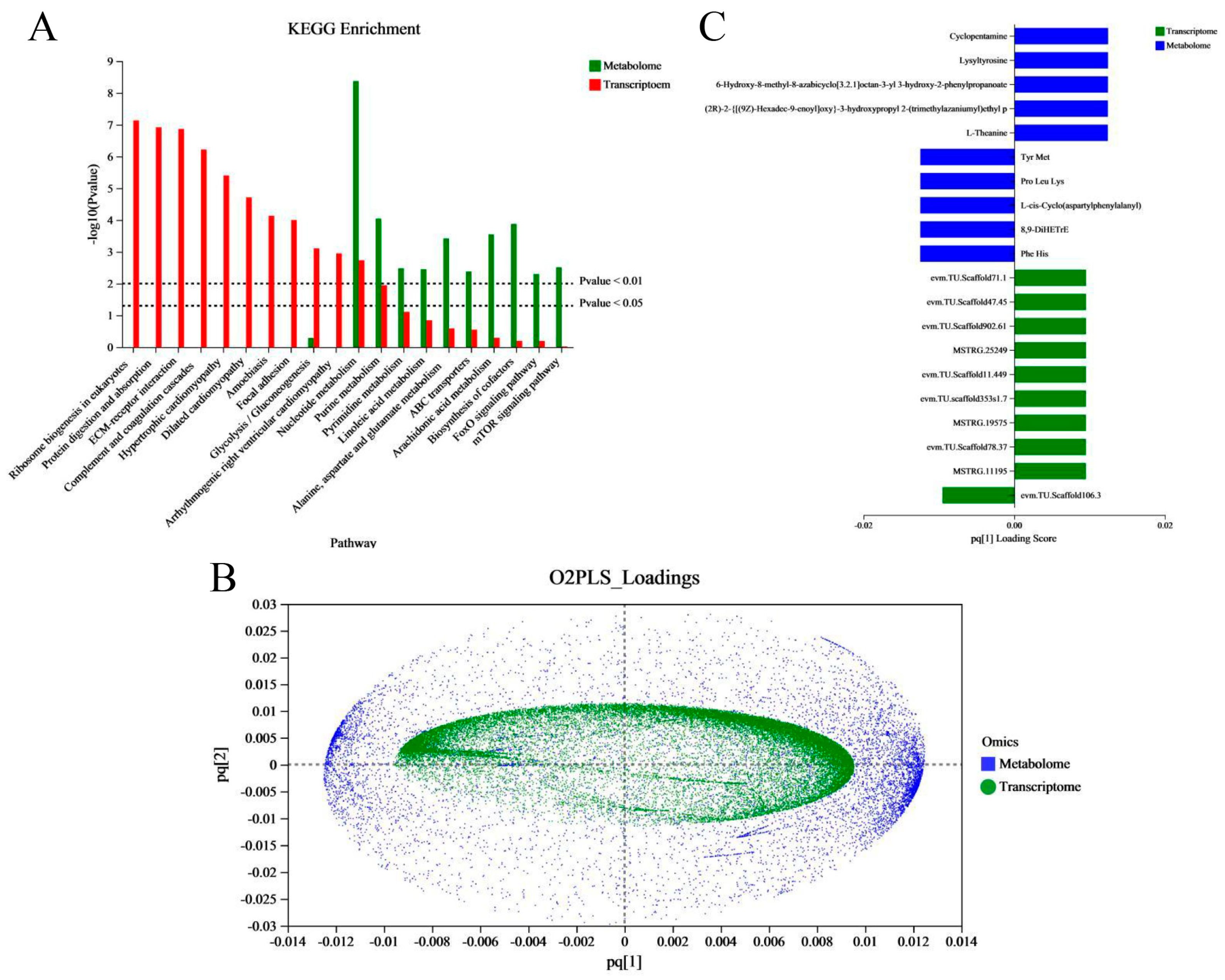
2.4. Correlation Analysis between the Transcriptomic and Metabonomic Data
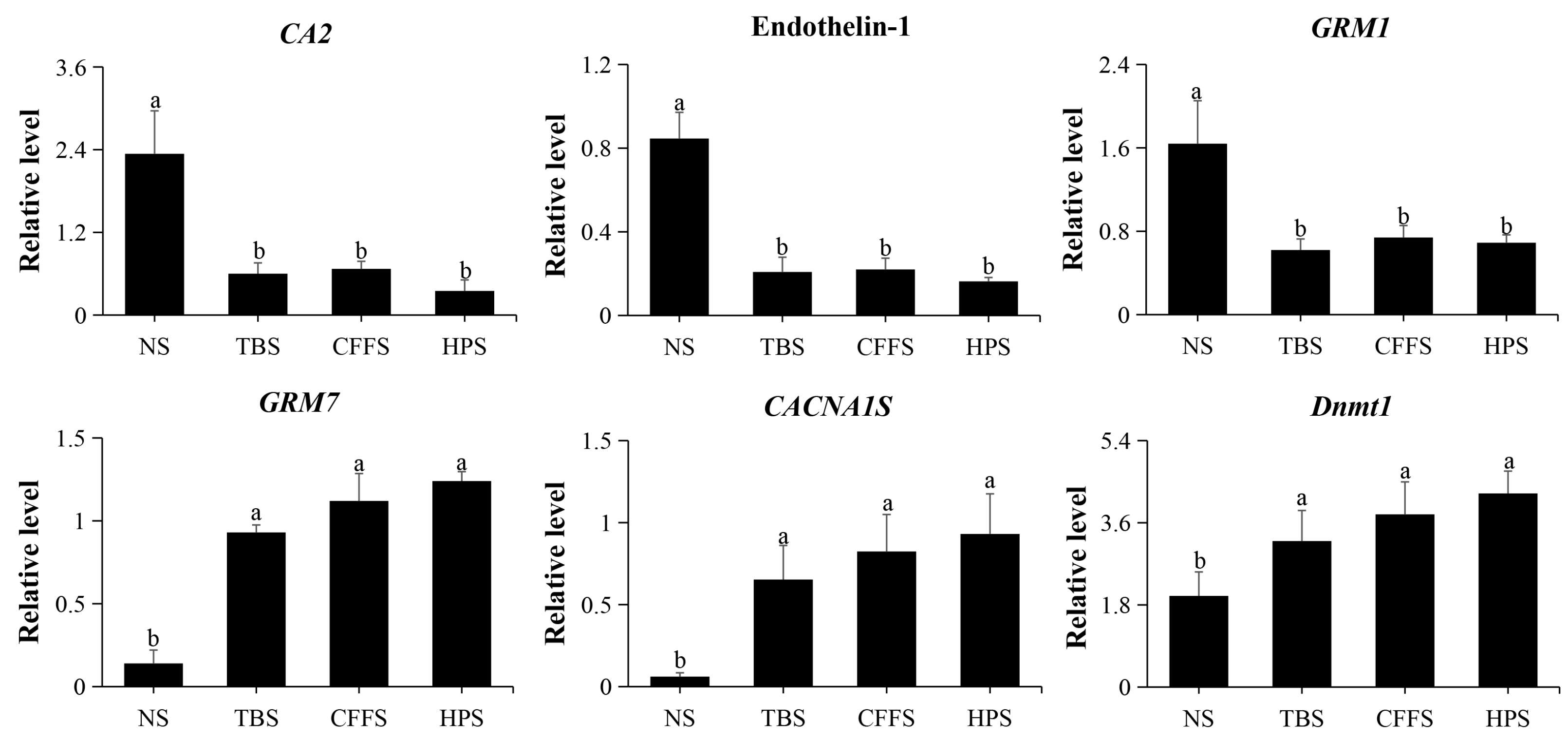
2.5. Confrmation of DEGs via qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. The Environment Sound Detection
4.3. Total RNA Isolation and Illumina Sequencing
4.4. Alignment of Transcriptomic Data
4.5. GO and KEGG Pathway Enrichment Analysis
4.6. Metabolite Extraction and UPLC-MS/MS Analysis
4.7. Metabolites Data Analysis
4.8. Identification of Differential Metabolites, and Pathway Enrichment Analysis
4.9. Validation of RNA-Seq Data via qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amoser, S.; Ladich, F. Are hearing sensitivities of freshwater fish adapted to the ambient noise in their habitats? J. Exp. Biol. 2005, 208, 3533–3542. [Google Scholar] [CrossRef]
- Soares, D.; Niemiller, M.L. Sensory adaptations of fishes to subterranean environments. Bioscience 2013, 63, 274–283. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, X.; Liu, Y.; Fu, Q.; Tian, C.; Wu, C.; Shi, H.; Yuan, Z.; Tan, S.; Liu, S.; et al. Transcriptome analysis reveals enrichment of genes associated with auditory system in swimbladder of channel catfish. Comp. Biochem. Phys. D 2018, 27, 30–39. [Google Scholar] [CrossRef]
- Weilgart, L.S. The impacts of anthropogenic ocean noise on cetaceans and implications for management. Can. J. Zool. 2007, 85, 1091–1116. [Google Scholar] [CrossRef]
- Popper, A.N.; Hastings, M.C. The effects of anthropogenic sources of sound on fishes. J. Fish. Biol. 2009, 75, 455–489. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.; Przeslawski, R.; Duncan, A.J.; Gunning, M.; Bruce, B. A critical review of the potential impacts of marine seismic surveys on fish & invertebrates. Mar. Pollut. Bull. 2017, 114, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Pieretti, N.; Martire, M.L.; Corinaldesi, C.; Musco, L.; Dell’anno, A.; Danovaro, R. Anthropogenic noise and biological sounds in a heavily industrialized coastal area (Gulf of Naples, Mediterranean Sea). Mar. Environ. Res. 2020, 159, 105002. [Google Scholar] [CrossRef] [PubMed]
- Dytham, C.; Simpson, S.D. Elevated mortality of fish larvae on coral reefs drives the evolution of larval movement patterns. Mar. Ecol. Prog. Ser. 2007, 346, 255–264. [Google Scholar] [CrossRef]
- Sierra-Flores, R.; Atack, T.; Migaud, H.; Davie, A. Stress response to anthropogenic noise in Atlantic cod Gadus morhua L. Aquacult. Eng. 2015, 67, 67–76. [Google Scholar] [CrossRef]
- Nedelec, S.L.; Mills, S.C.; Lecchini, D.; Nedelec, B.; Simpson, S.D.; Radford, A.N. Repeated exposure to noise increases tolerance in a coral reef fish. Environ. Pollut. 2016, 216, 428–436. [Google Scholar] [CrossRef]
- Shafiei-Sabet, S.; Van-Dooren, D.; Slabbekoorn, H. Son et lumiere: Sound and light effects on spatial distribution and swimming behavior in captive zebrafish. Environ. Pollut. 2016, 212, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.D.; Radford, A.N.; Nedelec, S.L.; Ferrari, M.C.O.; Chivers, D.P.; McCormick, M.I.; Meekan, M.G. Anthropogenic noise increases fish mortality by predation. Nat. Commun. 2016, 7, 10544. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-T.; Akamatsu, T.; Nowacek, D.P.; Yuan, J.; Zhou, L.; Lei, P.-Y.; Li, J.; Duan, P.-X.; Wang, K.-X.; Wang, D. Soundscape of an Indo-Pacific humpback dolphin (Sousa chinensis) hotspot before windfarm construction in the Pearl River Estuary, China: Do dolphin engage in noise avoidance and passive eavesdropping behavior? Mar. Pollut. Bull. 2019, 140, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Kusku, H.; Yigit, Ü.; Yilmaz, S.; Yigit, M.; Ergün, S. Acoustic effects of underwater drilling and piling noise on growth and physiological response of Nile tilapia (Oreochromis niloticus). Aquacult. Res. 2020, 51, 3166–3174. [Google Scholar] [CrossRef]
- Mills, S.C.; Beldade, R.; Henry, L.; Laverty, D.; Nedelec, S.L.; Simpson, S.D.; Radford, A.N. Hormonal and behavioural effects of motorboat noise on wild coral reef fish. Environ. Pollut. 2020, 262, 114250. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.; Bebak, J.; Mazik, P. The effects of aquaculture production noise on the growth, condition factor, feed conversion, and survival of rainbow trout, Oncorhynchus mykiss. Aquaculture 2009, 288, 337–343. [Google Scholar] [CrossRef]
- Nedelec, S.L.; Simpson, S.D.; Morley, E.L.; Nedelec, B.; Radford, A.N. Impacts of regular and random noise on the behaviour, growth and development of larval Atlantic cod (Gadus morhua). Proc. Biol. Sci. 2015, 282, 20151943. [Google Scholar] [CrossRef] [PubMed]
- Hang, S.; Zhao, J.; Ji, B.; Li, H.; Zhang, Y.; Peng, Z.; Zhou, F.; Ding, X.; Ye, Z. Impact of underwater noise on the growth, physiology and behavior of Micropterus salmoides in industrial recirculating aquaculture systems. Environ. Pollut. 2021, 291, 118152. [Google Scholar] [CrossRef]
- Friedrich, L.; Tanja, S.M. Diversity in Fish Auditory Systems: One of the Riddles of Sensory Biology. Front. Ecol. Evol. 2016, 4, 28. [Google Scholar] [CrossRef]
- Lowenstein, O. The equilibrium function of the vertebrate labyrinth. Biol. Rev. 1936, 11, 113–145. [Google Scholar] [CrossRef]
- Thomas, O.R.B.; Swearer, S.E.; Kapp, E.A.; Peng, P.; Tonkin-Hill, G.Q.; Papenfuss, A.; Roberts, A.; Bernard, P.; Roberts, B.R. The inner ear proteome of fish. FEBS J. 2019, 286, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Thiessen, K.D.; Higuchi, L.; Kramer, K.L. Zebrafish Otolith Biomineralization Requires Polyketide Synthase. Mech. Dev. 2019, 157, 1–9. [Google Scholar] [CrossRef]
- Mogdans, J.; Krther, S.; Engelmann, J. Electroreception: Strategies for Separation of Signals from Noise. In The Senses of Fish; Springer: Berlin/Heidelberg, Germany, 2004; pp. 330–361. [Google Scholar] [CrossRef]
- Lu, Z.; DeSmidt, A.A. Early Development of Hearing in Zebrafish. J. Assoc. Res. Otolaryngol. 2013, 14, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.A.; Cheng, A.G.; Cunningham, L.L.; MacDonald, G.; Raible, D.W.; Rubel, E.W. Neomycin-Induced Hair Cell Death and Rapid Regeneration in the Lateral Line of Zebrafish (Danio rerio). J. Assoc. Res. Otolaryngol. 2003, 4, 219–234. [Google Scholar] [CrossRef]
- Aval, F.S.; Sargazi, A. Genes associated with regeneration of utricle hair cells in mammalians: A review. IJBPAS 2016, 5, 3397–3408. [Google Scholar] [CrossRef]
- Dey, A.; Flajšhans, M.; Pšenička, M.; Gazo, I. DNA repair genes play a variety of roles in the development of fish embryos. Front. Cell Dev. Biol. 2023, 11, 1119229. [Google Scholar] [CrossRef]
- Wang, X.; Lu, G.; Zhao, L.; Yang, Q.; Gao, T. Assessment of fishery resources using environmental DNA: Small yellow croaker (Larimichthys polyactis) in East China Sea. PLoS ONE 2020, 15, e0244495. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, J.; Xu, W.; Zhan, W.; Zou, H.; Lin, J. Transcriptomic and Behavioral Studies of Small Yellow Croaker (Larimichthys polyactis) in Response to Noise Exposure. Animals 2022, 12, 2061. [Google Scholar] [CrossRef]
- Zhan, W.; Lou, B.; Chen, R.-Y.; Mao, G.-M.; Liu, F.; Xu, D.-D.; Wang, L.-G.; Ma, T.; Xu, Q.-X. Observation on embryonic development and morphological characteristics of Larimichthys polyactis. Oceanol. Limnol. Sin. 2016, 5, 7. [Google Scholar] [CrossRef]
- Yin, L.M.; Huang, H.L.; Cheng, X.Z. The study on the auditory characteristics of large yellow croaker under cage aquaculture. In Proceedings of the Academic Annual Meeting of Chinese Fisheries Society, Chengdu, China, 2 November 2016. [Google Scholar]
- Xie, Q.; Zhan, W.; Shi, J.; Liu, F.; Niu, B.; He, X.; Liu, M.; Wang, J.; Liang, Q.; Xie, Y.; et al. Whole-genome assembly and annotation for the little yellow croaker (Larimichthys polyactis) provide insights into the evolution of hermaphroditism and gonochorism. Mol. Ecol. Resour. 2022, 23, 632–658. [Google Scholar] [CrossRef]
- Hughes, I.; Blasiole, B.; Huss, D.; Warchol, M.E.; Rath, N.P.; Hurle, B.; Ignatova, E.; Dickman, J.D.; Thalmann, R.; Levenson, R.; et al. Otopetrin 1 is required for otolith formation in the zebrafish Danio rerio. Dev. Biol. 2004, 276, 391–402. [Google Scholar] [CrossRef]
- Weigele, J.; Franz-Odendaal, T.A.; Hilbig, R. Not all inner ears are the same: Otolith matrix proteins in the inner ear of sub-adult cichlid fish, oreochromis mossambicus, reveal insights into the biomineralization process. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2016, 299, 234–245. [Google Scholar] [CrossRef]
- Seist, R.; Landegger, L.D.; Robertson, N.G.; Vasilijic, S.; Morton, C.C.; Stankovic, K.M. Cochlin Deficiency Protects Against Noise-Induced Hearing Loss. Front. Mol. Neurosci. 2021, 14, 670013. [Google Scholar] [CrossRef]
- Sumanas, S.; Larson, J.D.; Bever, M.M. Zebrafish chaperone protein GP96 is required for otolith formation during ear development. Dev. Biol. 2003, 261, 443–455. [Google Scholar] [CrossRef]
- Davies, D.; Holley, M.C. Differential expression of alpha 3 and alpha 6 integrins in the developing mouse inner ear. J. Comp. Neurol. 2002, 445, 122. [Google Scholar] [CrossRef]
- Evans, A.L.; Ulrich, M. Stereocilia defects in the sensory hair cells of the inner ear in mice deficient in integrin α8β1. Nat. Genet. 2000, 24, 424–428. [Google Scholar] [CrossRef]
- Kiss, P.J.; Knisz, J.; Zhang, Y.; Baltrusaitis, J.; Sigmund, C.D.; Thalmann, R.; Smith, R.J.; Verpy, E.; Bánfi, B. Inactivation of NADPH oxidase organizer 1 Results in Severe Imbalance. Curr. Biol. 2006, 16, 208–213. [Google Scholar] [CrossRef]
- Schröder, K.; Weissmann, N.; Brandes, R.P. Organizers and activators: Cytosolic Nox proteins impacting on vascular function. Free. Radic. Biol. Med. 2017, 109, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Chen, X.; Lv, F.; Ren, H.; Fan, Z.; Qin, H.; Yu, L.; Shi, X.; Xu, Y. Serum response factor (SRF) promotes ROS generation and hepatic stellate cell activation by epigenetically stimulating NCF1/2 transcription. Redox Biol. 2019, 26, 101302. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, Y.W.; Xu, Y.; Thiessen, K.D.; Kramer, K.L. Mechanisms of otoconia and otolith development. Dev. Dyn. 2015, 244, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Lopez, I.A.; Ishiyama, G.; Acuna, D.; Ishiyama, A. Otopetrin-2 Immunolocalization in the Human Macula Utricle. Ann. Otol. Rhinol. Laryngol. 2019, 128, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Tohse, H.; Murayama, E.; Ohira, T.; Takagi, Y.; Nagasawa, H. Localization and diurnal variations of carbonic anhydrase mRNA expression in the inner ear of the rainbow trout Oncorhynchus mykiss. Comp. Biochem. Phys. B 2006, 145, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Paine, M.L. Bicarbonate Transport During Enamel Maturation. Calcif. Tissue Int. 2017, 101, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.; Tohse, H.; Murayama, E.; Ohira, T.; Nagasawa, H. Diel changes in endolymph aragonite saturation rate and mRNA expression of otolith matrix proteins in the trout otolith organ. Mar. Ecol. Prog. 2005, 294, 249–256. [Google Scholar] [CrossRef]
- Liu, Q.; Feng, B.; Meng, G.; Liu, Y. The study on plasma ET and NO of patients with sudden hearing loss. J. Clin. Otorhinolaryngol. 2003, 17, 668. [Google Scholar]
- White, R.J.; Collins, J.E.; Sealy, I.M.; Wali, N.; Dooley, C.M.; Digby, Z.; Stemple, D.L.; Murphy, D.N.; Billis, K.; Hourlier, T.; et al. A high-resolution mRNA expression time course of embryonic development in zebrafish. eLife 2017, 6, e30860. [Google Scholar] [CrossRef]
- Miller, C.T.; Schilling, T.F.; Lee, K.; Parker, J.; Kimmel, C.B. Sucker encodes a zebrafish Endothelin-1 required for ventral pharyngeal arch development. Development 2000, 127, 3815. [Google Scholar] [CrossRef]
- Barta, C.L.; Liu, H.; Chen, L.; Giffen, K.P.; Li, Y.; Kramer, K.L.; Beisel, K.W.; He, D.Z. RNA-seq transcriptomic analysis of adult zebrafish inner ear hair cells. Data 2018, 5, 180005. [Google Scholar] [CrossRef]
- Rotschafer, S.E.; Cramer, K.S. Developmental Emergence of Phenotypes in the Auditory Brainstem Nuclei of Fmr1 Knockout Mice. eNeuro 2017, 4, ENEURO.0264-17. [Google Scholar] [CrossRef]
- Bardoni, B.; Willemsen, R.; Weiler, I.J.; Schenck, A.; Severijnen, L.-A.; Hindelang, C.; Lalli, E.; Mandel, J.-L. NUFIP1 (nuclear FMRP interacting protein 1) is a nucleocytoplasmic shuttling protein associated with active synaptoneurosomes. Exp. Cell Res. 2003, 289, 95–107. [Google Scholar] [CrossRef]
- Chizhikov, V.V.; Iskusnykh, I.Y.; Fattakhov, N.; Fritzsch, B. Lmx1a and Lmx1b are Redundantly Required for the Development of Multiple Components of the Mammalian Auditory System. Neuroscience 2021, 452, 247–264. [Google Scholar] [CrossRef]
- Patel, D.; Shimomura, A.; Majumdar, S.; Holley, M.C.; Hashino, E. The histone demethylase LSD1 regulates inner ear progenitor differentiation through interactions with Pax2 and the NuRD repressor complex. PLoS ONE 2018, 13, e0191689. [Google Scholar] [CrossRef] [PubMed]
- Takago, H.; Oshima-Takago, T. Pre- and postsynaptic ionotropic glutamate receptors in the auditory system of mammals. Hear. Res. 2018, 362, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Curry, R.J.; Kang, P.; Yong, L. Neurotransmitter- and release mode-specific modulation of inhibitory transmission by group I metabotropic glutamate receptors in central auditory neurons of the mouse. J. Neurosci. 2018, 38, 8187–8199. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Wang, X.; Wang, Y.; Li, D.; Huang, H.; Lu, Y. Mechanisms Underlying Enhancement of Spontaneous Glutamate Release by Group I mGluRs at a Central Auditory Synapse. J. Neurosci. 2020, 40, 7027–7042. [Google Scholar] [CrossRef] [PubMed]
- Rivolta, C.; Sweklo, E.A.; Berson, E.L.; Dryja, T.P. Missense Mutation in the USH2A Gene: Association with Recessive Retinitis Pigmentosa without Hearing Loss. Am. J. Hum. Genet. 2000, 66, 1975–1978. [Google Scholar] [CrossRef] [PubMed]
- Dona, M.; Slijkerman, R.; Lerner, K.; Broekman, S.; Wegner, J.; Howat, T.; Peters, T.; Hetterschijt, L.; Boon, N.; de Vrieze, E.; et al. Usherin defects lead to early-onset retinal dysfunction in zebrafish. Exp. Eye Res. 2018, 173, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Coate, T.M.; A Spita, N.; Zhang, K.D.; Isgrig, K.T.; Kelley, M.W. Neuropilin-2/Semaphorin-3F-mediated repulsion promotes inner hair cell innervation by spiral ganglion neurons. eLife 2015, 4, e07830. [Google Scholar] [CrossRef]
- Scott, K.E.; Schormans, A.L.; Pacoli, K.Y.; De Oliveira, C.; Allman, B.L.; Schmid, S. Altered Auditory Processing, Filtering, and Reactivity in the Cntnap2 Knock-Out Rat Model for Neurodevelopmental Disorders. J. Neurosci. 2018, 38, 8588–8604. [Google Scholar] [CrossRef]
- Krasewicz, J.; Yu, W.M. Eph and ephrin signaling in the development of the central auditory system. Dev. Dyn. 2022, 252, 10–26. [Google Scholar] [CrossRef]
- Elbracht, M.; Senderek, J.; Eggermann, T.; Thürmer, C.; Park, J.; Westhofen, M.; Zerres, K. Autosomal recessive postlingual hearing loss (DFNB8): Compound heterozygosity for two novel TMPRSS3 mutations in German siblings. J. Med. Genet. 2007, 44, e81. [Google Scholar] [CrossRef]
- Mellott, J.G.; Beebe, N.L.; Schofield, B.R. GABAergic and non-GABAergic projections to the superior colliculus from the auditory brainstem. Brain Struct. Funct. 2018, 223, 1923–1936. [Google Scholar] [CrossRef]
- Lalwani, P.; Gagnon, H.; Cassady, K.; Simmonite, M.; Peltier, S.; Seidler, R.D.; Taylor, S.F.; Weissman, D.H.; Polk, T.A. Neural distinctiveness declines with age in auditory cortex and is associated with auditory GABA levels. NeuroImage 2019, 201, 116033. [Google Scholar] [CrossRef]
- Newman, D.L.; Fisher, L.M.; Ohmen, J.; Parody, R.; Fong, C.-T.; Frisina, S.T.; Mapes, F.; Eddins, D.A.; Frisina, D.R.; Frisina, R.D.; et al. GRM7 variants associated with age-related hearing loss based on auditory perception. Hear. Res. 2012, 294, 125–132. [Google Scholar] [CrossRef]
- Wu, X.; Indzhykulian, A.A.; Niksch, P.D.; Webber, R.M.; Garcia-Gonzalez, M.; Watnick, T.; Zhou, J.; Vollrath, M.A.; Corey, D.P. Hair-Cell Mechanotransduction Persists in TRP Channel Knockout Mice. PLoS ONE 2016, 11, e0155577. [Google Scholar] [CrossRef]
- Shpargel, K.B.; Makishima, T.; Griffith, A.J. Col11a1 and Col11a2 mRNA expression in the developing mouse cochlea: Implications for the correlation of hearing loss phenotype with mutant type XI collagen genotype. Acta Oto-Laryngologica 2004, 124, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Machnicka, B.; Grochowalska, R.; Bogusławska, D.M.; Sikorski, A.F.; Lecomte, M.C. Spectrin-based skeleton as an actor in cell signaling. Cell. Mol. Life Sci. 2012, 69, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-J.; Stevenson, A.K.; Yau, P.M.; Kollmar, R. Sparc protein is required for normal growth of zebrafish otoliths. J. Assoc. Res. Otolaryngol. 2008, 9, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Huang, L.; Wei, Z.-B.; Silvius, D.; Tang, B.; Xu, P.-X. The role of Six1 in mammalian auditory system development. Development 2003, 130, 3989–4000. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Zheng, S.; Zheng, Z.; Liu, C.; Zhang, J.; Yan, R.; Wu, C.; Zuo, N.; Wu, L.; Xu, H.; et al. Dnmt1 is required for the development of auditory organs via cell cycle arrest and Fgf signalling. Cell Prolif. 2022, 55, e13225. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, V.; Gupta, S. Role of osteocytes in mediating bone mineralization during hyperhomocysteinemia. J. Endocrinol. 2017, 233, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Nitzahn, M.; Lipshutz, G.S. CPS1: Looking at an ancient enzyme in a modern light. Mol. Genet. Metab. 2020, 131, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Speed, J.S.; George, E.M.; Arany, M.; Cockrell, K.; Granger, J.P. Role of 20-Hydroxyeicosatetraenoic Acid in Mediating Hypertension in Response to Chronic Renal Medullary Endothelin Type B Receptor Blockade. PLoS ONE 2011, 6, e26063. [Google Scholar] [CrossRef] [PubMed]
- Medhora, M.; Chen, Y.; Harland, D.; Bodiga, S.; Zielonka, J.; Gebremedhin, D.; Gao, Y.; Anjaiah, S.; Jacobs, E.R.; Zhang, C.; et al. 20-HETE increases superoxide production and activates NAPDH oxidase in pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, 902–911. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Res. 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Tang, H.; Klopfenstein, D.; Pedersen, B.; Flick, P.; Sato, K.; Ramirez, F.; Yunes, J.; Mungall, C. GOATOOLS: Tools for gene ontology. Zenodo 2015. Available online: https://zenodo.org/records/819475 (accessed on 20 June 2023).
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Yu, G.; Shi, C.; Liu, L.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.; Gao, H.; et al. Majorbio Cloud: A one-stop, comprehensive bioinformatic platform for multi-omics analyses. iMeta 2022, 1, e12. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Group | Sample Name | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Q30 (%) | GC Content (%) | Mapped Reads | Mapped Ratio (%) |
|---|---|---|---|---|---|---|---|---|---|
| TR_A1 | MS1_1 | 46,560,464 | 7,030,630,064 | 45,613,948 | 6,659,981,695 | 93.35 | 47.97 | 35,026,369 | 76.79 |
| MS1_3 | 46,079,848 | 6,958,057,048 | 45,478,904 | 6,718,090,478 | 93.96 | 48.55 | 38,418,011 | 84.47 | |
| MS1_4 | 45,763,860 | 6,910,342,860 | 44,868,434 | 6,486,429,872 | 94.16 | 47.83 | 35,457,605 | 79.03 | |
| TR_A2 | MS2_1 | 43,973,876 | 6,640,055,276 | 43,310,308 | 6,441,264,366 | 93.97 | 49.11 | 37,017,865 | 85.47 |
| MS2_2 | 45,550,584 | 6,878,138,184 | 44,926,238 | 6,662,511,904 | 93.91 | 48.63 | 37,569,943 | 83.63 | |
| MS2_3 | 43,448,516 | 6,560,725,916 | 42,889,492 | 6,381,334,581 | 93.84 | 49.51 | 36,411,339 | 84.9 | |
| TR_A3 | MS3_2 | 44,253,422 | 6,682,266,722 | 43,604,360 | 6,443,996,202 | 94.07 | 47.73 | 36,374,553 | 83.42 |
| MS3_4 | 42,974,092 | 6,489,087,892 | 42,461,328 | 6,280,080,349 | 94.12 | 49.46 | 36,352,605 | 85.61 | |
| MS3_6 | 49,807,428 | 7,520,921,628 | 49,213,898 | 7,316,291,487 | 93.65 | 49.4 | 42,158,826 | 85.66 | |
| TR_A4 | MS4_2 | 48,560,896 | 7,332,695,296 | 48,013,414 | 7,121,588,700 | 94.09 | 49.91 | 41,594,404 | 86.63 |
| MS4_3 | 51,011,010 | 7,702,662,510 | 50,454,496 | 7,500,276,460 | 94.14 | 49.08 | 43,603,271 | 86.42 | |
| MS4_4 | 50,702,630 | 7,656,097,130 | 50,055,884 | 7,424,923,409 | 93.93 | 48.69 | 43,144,247 | 86.19 |
| Gene | A1_vs_A2 | A1_vs_A3 | A1_vs_A4 | A2_vs_A3 | A2_vs_A4 | A3_vs_A4 | Sequence_ID |
|---|---|---|---|---|---|---|---|
| Carbonic anhydrase 2 | yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold208.24 |
| Integrin α-6 | yes|up | yes|up | yes|up | no|down | no|down | no|up | evm.TU.Scaffold254.286 |
| Integrin α-5 | yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold804.200 |
| Integrin α-3 CD49 antigen-like family member C | yes|up | no|up | yes|up | no|down | no|down | no|up | evm.TU.Scaffold1029.76 |
| Otopetrin-2 | yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold84.30 |
| yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold114.78 | |
| yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold114.79 | |
| Neutrophil cytosol factor 2 | yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold620.51 |
| NADPH oxidase 1 | yes|up | yes|up | yes|up | no|up | no|up | no|down | evm.TU.Scaffold476.222 |
| Transmembrane protease serine | no|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold238.12 |
| Metabotropic glutamate receptor-1 | yes|up | yes|up | yes|up | no|down | no|up | no|up | evm.TU.Scaffold169.511 |
| Neuropilin-2 | yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold254.538 |
| Integrin β-1 | no|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold620.12 |
| Endothelin-1 | yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.scaffold325s1.3 |
| Cntnap2 | yes|up | yes|up | yes|up | no|down | no|down | no|up | evm.TU.Scaffold123.15 |
| no|up | yes|up | yes|up | no|up | no|up | no|down | evm.TU.scaffold125s1.6 | |
| NUFIP1 | yes|up | yes|up | yes|up | no|up | no|up | no|down | evm.TU.Scaffold133.51 |
| Ephrin type-A receptor 7 | yes|up | yes|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold169.12 |
| USH2A | no|up | yes|up | yes|up | no|up | no|up | no|down | evm.TU.Scaffold69.380 |
| NADPH oxidase 4 | no|up | no|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold69.544 |
| Pax2 | no|up | no|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold63.125 |
| Lmx1a | no|up | no|up | yes|up | no|up | no|up | no|up | evm.TU.Scaffold114.16 |
| Carbonic anhydrase 4 | yes|down | yes|down | yes|down | no|up | no|down | no|down | evm.TU.Scaffold1817.320 |
| Collagen α-1(XI) chain | yes|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold789.161 |
| yes|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold620.61 | |
| Voltage-dependent L-type calcium channel subunit α-1S | yes|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold411.4 |
| Excitatory amino acid transporter 3 | no|down | no|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold630.83 |
| Fatty acid synthase | yes|down | yes|down | yes|down | no|down | no|down | no|up | evm.TU.scaffold700s1.8 |
| Otolith Matrix Protein-1 | no|down | yes|down | yes|down | no|down | no|down | no|up | evm.TU.Scaffold1603.13 |
| α-Tectorin | yes|down | yes|down | yes|down | no|up | no|down | no|down | evm.TU.Scaffold195.21 |
| yes|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold611.1 | |
| NADPH oxidase organizer 1 | yes|down | no|down | yes|down | yes|up | no|up | no|down | evm.TU.Scaffold1029.136 |
| Carbonic anhydrase 6 | yes|down | yes|down | yes|down | no|up | no|up | no|down | evm.TU.Scaffold293.98 |
| Gamma-aminobutyric acid receptor subunit β-3 | yes|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold158.122 |
| Spectrin β chain, non-erythrocytic 1 | yes|down | yes|down | yes|down | no|up | no|up | no|down | evm.TU.Scaffold195.103 |
| Metabotropic glutamate receptor 7 | yes|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold2937.38 |
| Transient receptor potential cation channel subfamily M member 2 | no|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold254.405 |
| Ephrin type-B receptor 1-B | yes|down | no|down | yes|down | no|up | no|down | no|down | evm.TU.Scaffold165.282 |
| no|down | no|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold1030.19 | |
| Osteonectin | yes|down | yes|down | yes|down | no|down | no|down | no|down | evm.TU.Scaffold78.49 |
| Neuroserpin | yes|down | no|down | yes|down | no|down | no|down | no|down | evm.TU.scaffold188s1.1 |
| yes|down | yes|down | yes|down | no|up | no|down | no|down | evm.TU.Scaffold857.38 | |
| Homeobox protein SIX1 | yes|down | yes|down | yes|down | no|up | no|down | no|down | evm.TU.Scaffold1603.234 |
| DNA (cytosine-5)-methyltransferase 1 | yes|down | yes|down | yes|down | no|up | no|down | no|down | evm.TU.Scaffold282.335 |
| Gene | Sequence_ID | Regulate Metabolite | Common Annotated Pathway | Pathway ID |
|---|---|---|---|---|
| CA2 | evm.TU.Scaffold208.24 | L-Glutamic Acid | Nitrogen metabolism | lco00910 |
| CA4 | evm.TU.Scaffold1817.320 | L-Glutamic Acid | Nitrogen metabolism | lco00910 |
| CA6 | evm.TU.Scaffold293.98 | L-Glutamic Acid | Nitrogen metabolism | lco00910 |
| GABA A receptor | evm.TU.Scaffold158.122 | N-Acetyl-1-aspartylglutamic acid; p-Octopamine; L-glutamic Acid | Neuroactive ligand–receptor interaction | lco04080 |
| GRM7 | evm.TU.Scaffold2937.38 | N-Acetyl-1-aspartylglutamic acid; P-Octopamine; L-Glutamic Acid | Neuroactive ligand–receptor interaction | lco04080 |
| CACNA1S | evm.TU.Scaffold411.4 | PA(8:0/8:0); PA(8:0/10:0); Arachidonic acid | GnRH signaling pathway | lco04912 |
| Arachidonic acid | Vascular smooth muscle contraction | lco04270 | ||
| Isoproterenol | Adrenergic signaling in cardiomyocytes | lco04261 | ||
| Endothelin-1 | evm.TU.scaffold325s1.3 | Arachidonic acid | Vascular smooth muscle contraction | lco04270 |
| N-Acetyl-1-aspartylglutamic acid; P-Octopamine; L-Glutamic Acid | Neuroactive ligand–receptor interaction | lco04080 | ||
| GRM1 | evm.TU.Scaffold169.511 | Adenosine 5′-Monophosphate; L-Glutamic Acid | FoxO signaling pathway | lco04068 |
| N-Acetyl-1-aspartylglutamic acid; P-Octopamine; L-Glutamic Acid | Neuroactive ligand–receptor interaction | lco04080 | ||
| L-Glutamic Acid | Gap junction | lco04540 | ||
| Dnmt1 | evm.TU.Scaffold282.335 | S-Adenosylhomocysteine; Methionine Sulfoxide; L-Cystathionine | Cysteine and methionine metabolism | lco00270 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Liang, X.; Ye, T.; Zhang, Y.; Lou, B. Metabonomics and Transcriptomics Analyses Reveal the Development Process of the Auditory System in the Embryonic Development Period of the Small Yellow Croaker under Background Noise. Int. J. Mol. Sci. 2024, 25, 1954. https://doi.org/10.3390/ijms25041954
Jiang Q, Liang X, Ye T, Zhang Y, Lou B. Metabonomics and Transcriptomics Analyses Reveal the Development Process of the Auditory System in the Embryonic Development Period of the Small Yellow Croaker under Background Noise. International Journal of Molecular Sciences. 2024; 25(4):1954. https://doi.org/10.3390/ijms25041954
Chicago/Turabian StyleJiang, Qinghua, Xiao Liang, Ting Ye, Yu Zhang, and Bao Lou. 2024. "Metabonomics and Transcriptomics Analyses Reveal the Development Process of the Auditory System in the Embryonic Development Period of the Small Yellow Croaker under Background Noise" International Journal of Molecular Sciences 25, no. 4: 1954. https://doi.org/10.3390/ijms25041954
APA StyleJiang, Q., Liang, X., Ye, T., Zhang, Y., & Lou, B. (2024). Metabonomics and Transcriptomics Analyses Reveal the Development Process of the Auditory System in the Embryonic Development Period of the Small Yellow Croaker under Background Noise. International Journal of Molecular Sciences, 25(4), 1954. https://doi.org/10.3390/ijms25041954