
Sofosbuvir Suppresses the Genome Replication of DENV1 in Human Hepatic Huh7 Cells

 , ,
, ,
Abstract
1. Introduction
2. Results
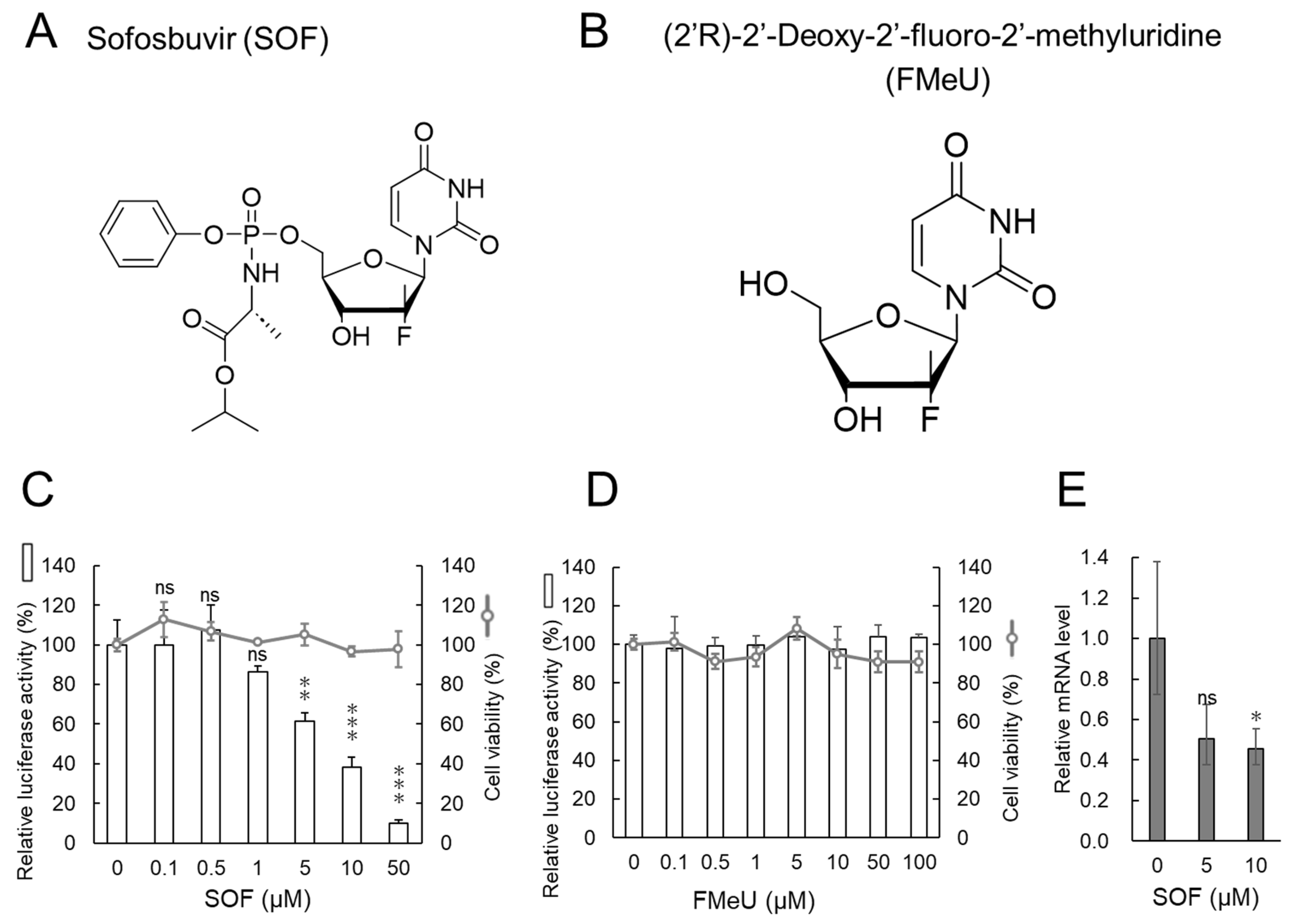
2.1. Replication of the DENV1 Replicon in Human Hepatic Huh7 Cells Is Suppressed by SOF but Not by FMeU
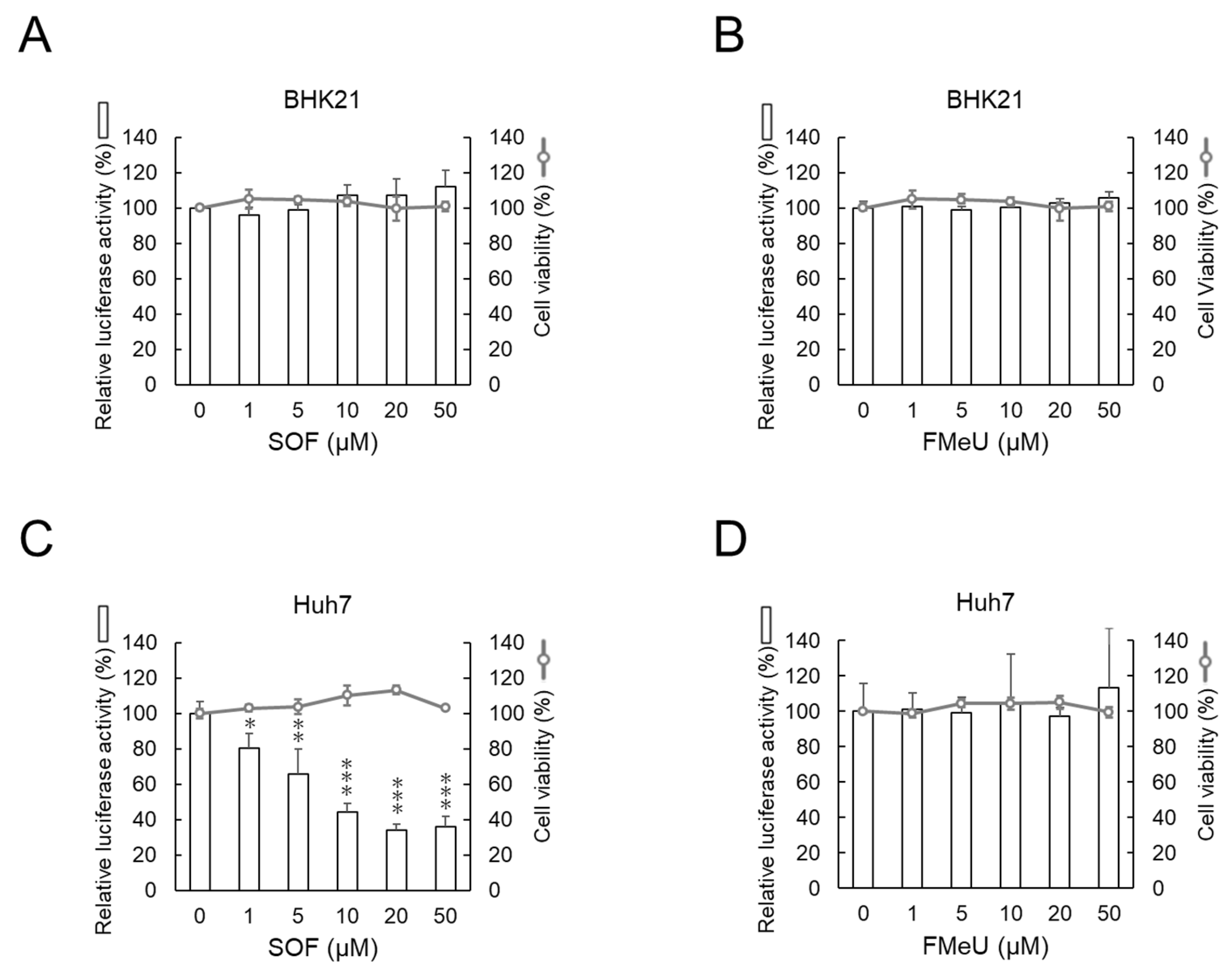
2.2. SOF Suppresses the Replication of the DENV1 Replicon in Human Huh7 Cells but Not in Hamster BHK-21 Cells
2.3. SOF Suppresses DENV1 Production in Human Hepatic Huh7 Cells but Not in Monkey Kidney Vero Cells
3. Discussion
4. Material and Methods
4.1. Cells and Compounds
4.2. Cell Viability Assay
4.3. Replicon-Harboring Cell Assay
4.4. Transient Replicon Assay
4.5. Quantitative Reverse Transcription PCR (RT-qPCR)
4.6. DENV1 Infection and CPE-Based Plaque Assay
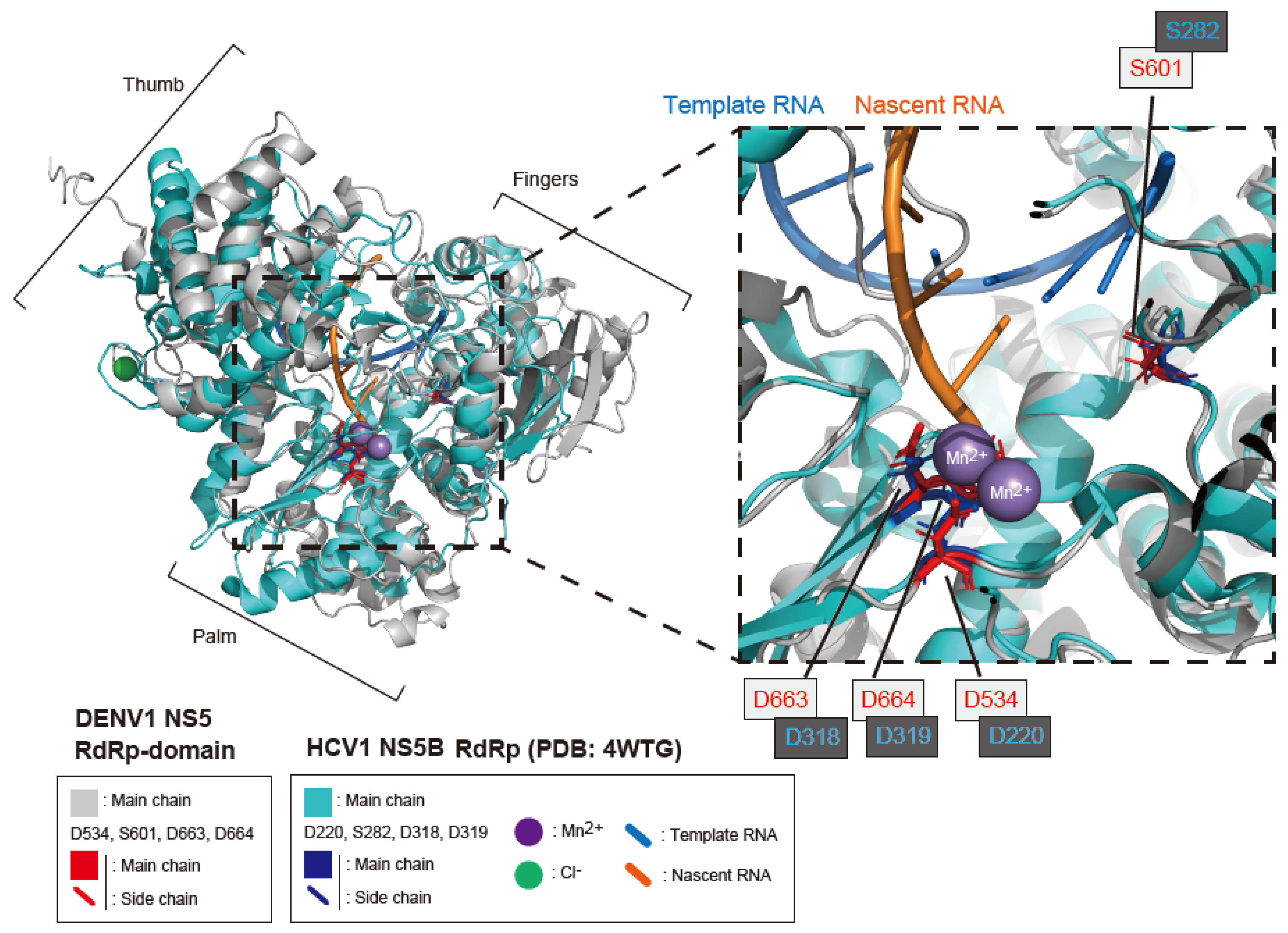
4.7. Molecular Docking Modeling
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gubler, D.J. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Simmons, C.P.; Farrar, J.J.; Nguyen, V.V.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 1423–1432. [Google Scholar] [CrossRef]
- World Health Organization (Regional Office for the Eastern Mediterranean). Dengue and Severe Dengue. Available online: https://apps.who.int/iris/handle/10665/204161 (accessed on 10 December 2023).
- World Health Organization. Fact Sheet, Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 10 December 2023).
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Kuhn, R.J.; Zhang, W.; Rossmann, M.G.; Pletnev, S.V.; Corver, J.; Lenches, E.; Jones, C.T.; Mukhopadhyay, S.; Chipman, P.R.; Strauss, E.G.; et al. Structure of dengue virus: Implications for flavivirus organization, maturation, and fusion. Cell 2002, 108, 717–725. [Google Scholar] [CrossRef]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef]
- Rawlinson, S.M.; Pryor, M.J.; Wright, P.J.; Jans, D.A. Dengue virus RNA polymerase NS5: A potential therapeutic target? Curr. Drug Targets. 2006, 7, 1623–1638. [Google Scholar] [CrossRef]
- Pathania, S.; Rawal, R.K.; Singh, P.K. RdRp (RNA-dependent RNA polymerase): A key target providing anti-virals for the management of various viral diseases. J. Mol. Struct. 2022, 1250, 131756. [Google Scholar] [CrossRef]
- Kumar, R.; Mishra, S.; Shreya; Maurya, S.K. Recent advances in the discovery of potent RNA-dependent RNA-polymerase (RdRp) inhibitors targeting viruses. RSC Med. Chem. 2020, 12, 306–320. [Google Scholar] [CrossRef]
- Malet, H.; Massé, N.; Selisko, B.; Romette, J.L.; Alvarez, K.; Guillemot, J.C.; Tolou, H.; Yap, T.L.; Vasudevan, S.; Lescar, J.; et al. The flavivirus polymerase as a target for drug discovery. Antivir. Res. 2008, 80, 23–35. [Google Scholar] [CrossRef]
- Roy, V.; Agrofoglio, L.A. Nucleosides and emerging viruses: A new story. Drug Discov. Today 2022, 7, 1945–1953. [Google Scholar] [CrossRef]
- Lim, S.P.; Noble, C.G.; Shi, P.Y. The dengue virus NS5 protein as a target for drug discovery. Antivir. Res. 2015, 119, 57–67. [Google Scholar] [CrossRef]
- Lim, S.P.; Wang, Q.Y.; Noble, C.G.; Chen, Y.L.; Dong, H.; Zou, B.; Yokokawa, F.; Nilar, S.; Smith, P.; Beer, D.; et al. Ten years of dengue drug discovery: Progress and prospects. Antivir. Res. 2013, 100, 500–519. [Google Scholar] [CrossRef]
- Latour, D.R.; Jekle, A.; Javanbakht, H.; Henningsen, R.; Gee, P.; Lee, I.; Tran, P.; Ren, S.; Kutach, A.K.; Harris, S.F.; et al. Biochemical characterization of the inhibition of the dengue virus RNA polymerase by beta-d-2′-ethynyl-7-deaza-adenosine triphosphate. Antivir. Res. 2010, 87, 213–222. [Google Scholar] [CrossRef]
- Yin, Z.; Chen, Y.L.; Schul, W.; Wang, Q.Y.; Gu, F.; Duraiswamy, J.; Kondreddi, R.R.; Niyomrattanakit, P.; Lakshminarayana, S.B.; Goh, A.; et al. An adenosine nucleoside inhibitor of dengue virus. Proc. Natl. Acad. Sci. USA 2009, 106, 20435–20439. [Google Scholar] [CrossRef]
- Sofia, M.J.; Bao, D.; Chang, W.; Du, J.; Nagarathnam, D.; Rachakonda, S.; Reddy, P.G.; Ross, B.S.; Wang, P.; Zhang, H.R.; et al. Discovery of a b-d-2′-deoxy-2′-a-fluoro-2′-b-C-methyluridine nucleotide prodrug (PSI-7977) for the treatment of hepatitis C virus. J. Med. Chem. 2010, 53, 7202–7218. [Google Scholar] [CrossRef]
- Lawitz, E.; Lalezari, J.P.; Hassanein, T.; Kowdley, K.V.; Poordad, F.F.; Sheikh, A.M.; Afdhal, N.H.; Bernstein, D.E.; Dejesus, E.; Freilich, B.; et al. Sofosbuvir in combination with peginterferon alfa-2a and ribavirin for non-cirrhotic, treatment-naive patients with genotypes 1, 2, and 3 hepatitis C infection: A randomised, double-blind, phase 2 trial. Lancet Infect. Dis. 2013, 5, 401–408. [Google Scholar] [CrossRef]
- Jacobson, I.M.; Gordon, S.C.; Kowdley, K.V.; Yoshida, E.M.; Rodriguez-Torres, M.; Sulkowski, M.S.; Shiffman, M.L.; Lawitz, E.; Everson, G.; Bennett, M.; et al. Sofosbuvir for hepatitis C genotype 2 or 3 in patients without treatment options. N. Engl. J. Med. 2013, 368, 1867–1877. [Google Scholar] [CrossRef]
- Fung, A.; Jin, Z.; Dyatkina, N.; Wang, G.; Beigelman, L.; Deval, J. Efficiency of incorporation and chain termination determines the inhibition potency of 2′-modified nucleotide analogs against hepatitis C virus polymerase. Antimicrob. Agents Chemother. 2014, 58, 3636–3645. [Google Scholar] [CrossRef]
- Appleby, T.C.; Perry, J.K.; Murakami, E.; Barauskas, O.; Feng, J.; Cho, A.; Fox, D., 3rd; Wetmore, D.R.; McGrath, M.E.; Ray, A.S.; et al. Viral replication. Structural basis for RNA replication by the hepatitis C virus polymerase. Science 2015, 347, 771–775. [Google Scholar] [CrossRef]
- Ma, H.; Jiang, W.R.; Robledo, N.; Leveque, V.; Ali, S.; Lara-Jaime, T.; Masjedizadeh, M.; Smith, D.B.; Cammack, N.; Klumpp, K.; et al. Characterization of the metabolic activation of hepatitis C virus nucleoside inhibitor beta-D-2′-Deoxy-2′-fluoro-2′-C-methylcytidine (PSI-6130) and identification of a novel active 5′-triphosphate species. J. Biol. Chem. 2007, 282, 29812–29820. [Google Scholar] [CrossRef]
- FDA Drug Trials Snapshots: EPCLUSA (Prescribing Information, Reference ID: 3951919). Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2016/208341s000lbl.pdf (accessed on 10 December 2023).
- Xu, H.T.; Colby-Germinario, S.P.; Hassounah, S.A.; Fogarty, C.; Osman, N.; Palanisamy, N.; Han, Y.; Oliveira, M.; Quan, Y.; Wainberg, M.A. Evaluation of Sofosbuvir (β-D-2′-deoxy-2′-α-fluoro-2′-β-C-methyluridine) as an inhibitor of Dengue virus replication. Sci. Rep. 2017, 7, 6345. [Google Scholar] [CrossRef]
- National Institute of Infectious Diseases. Dengue fever and dengue hemorrhagic fever in 2011–2014. Infect. Agents Surveill. Rep. (IASR) 2015, 36, 33–35. Available online: https://www.niid.go.jp/niid/images/idsc/iasr/36/421j.pdf (accessed on 20 January 2024).
- Guo, R.N.; Lin, J.Y.; Li, L.H.; Ke, C.W.; He, J.F.; Zhong, H.J.; Zhou, H.Q.; Peng, Z.Q.; Yang, F.; Liang, W.J. The prevalence and endemic nature of dengue infections in Guangdong, South China: An epidemiological, serological, and etiological study from 2005–2011. PLoS ONE 2014, 9, e85596. [Google Scholar] [CrossRef]
- Takasaki, T. Domestic dengue epidemic in Japan. 2014. Virus 2015, 56, 115–118. [Google Scholar] [CrossRef]
- Shen, S.Q.; Wei, H.X.; Fu, Y.H.; Zhang, H.; Mo, Q.Y.; Wang, X.J.; Deng, S.Q.; Zhao, W.; Liu, Y.; Feng, X.S.; et al. Multiple Sources of Infection and Potential Endemic Characteristics of the Large Outbreak of Dengue in Guangdong in 2014. Sci. Rep. 2015, 5, 16913. [Google Scholar] [CrossRef]
- Kato, F.; Kobayashi, T.; Tajima, S.; Takasaki, T.; Miura, T.; Igarashi, T.; Hishiki, T. Development of a novel Dengue-1 virus replicon system expressing secretory Gaussia luciferase for analysis of viral replication and discovery of antiviral drugs. JPN J. Infect. Dis. 2014, 67, 209–212. [Google Scholar] [CrossRef]
- Kato, F.; Nio, Y.; Yagasaki, K.; Suzuki, R.; Hijikata, M.; Miura, T.; Miyazaki, I.; Tajima, S.; Lim, C.K.; Saijo, M.; et al. Identification of inhibitors of dengue viral replication using replicon cells expressing secretory luciferase. Antivir. Res. 2019, 172, 104643. [Google Scholar] [CrossRef]
- Hishiki, T.; Kato, F.; Tajima, S.; Toume, K.; Umezaki, M.; Takasaki, T.; Miura, T. Hirsutine, an Indole Alkaloid of Uncaria rhynchophylla, Inhibits Late Step in Dengue Virus Lifecycle. Front. Microbiol. 2017, 8, 1674. [Google Scholar] [CrossRef]
- Hishiki, T.; Kato, F.; Nio, Y.; Watanabe, S.; Wen Tan, N.W.; Yamane, D.; Miyazaki, Y.; Lin, C.C.; Suzuki, R.; Tajima, S.; et al. Stearoyl-CoA desaturase-1 is required for flavivirus RNA replication. Antivir. Res. 2019, 165, 42–46. [Google Scholar] [CrossRef]
- Okano, Y.; Saito-Tarashima, N.; Kurosawa, M.; Iwabu, A.; Ota, M.; Watanabe, T.; Kato, F.; Hishiki, T.; Fujimuro, M.; Minakawa, N. Synthesis and biological evaluation of novel imidazole nucleosides as potential anti-dengue virus agents. Bioorg. Med. Chem. 2019, 27, 2181–2186. [Google Scholar] [CrossRef]
- Leyssen, P.; Balzarini, J.; De Clercq, E.; Neyts, J. The predominant mechanism by which ribavirin exerts its antiviral activity in vitro against flaviviruses and paramyxoviruses is mediated by inhibition of IMP dehydrogenase. J. Virol. 2005, 79, 1943–1947. [Google Scholar] [CrossRef]
- Shum, D.; Smith, J.L.; Hirsch, A.J.; Bhinder, B.; Radu, C.; Stein, D.A.; Nelson, J.A.; Früh, K.; Djaballah, H. High-content assay to identify inhibitors of dengue virus infection. Assay Drug Dev. Technol. 2010, 8, 553–570. [Google Scholar] [CrossRef]
- Tseng, C.H.; Lin, C.K.; Chen, Y.L.; Hsu, C.Y.; Wu, H.N.; Tseng, C.K.; Lee, J.C. Synthesis, antiproliferative and anti-dengue virus evaluations of 2-aroyl-3-arylquinoline derivatives. Eur. J. Med. Chem. 2014, 79, 66–76. [Google Scholar] [CrossRef]
- Yap, T.L.; Xu, T.; Chen, Y.L.; Malet, H.; Egloff, M.P.; Canard, B.; Vasudevan, S.G.; Lescar, J. Crystal structure of the dengue virus RNA-dependent RNA polymerase catalytic domain at 1.85-angstrom resolution. J. Virol. 2007, 81, 4753–4765. [Google Scholar] [CrossRef]
- Wu, J.; Wang, X.; Liu, Q.; Lu, G.; Gong, P. Structural basis of transition from initiation to elongation in de novo viral RNA-dependent RNA polymerases. Proc. Natl. Acad. Sci. USA 2023, 120, e2211425120. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Watanabe, T.; Iwaisako, Y.; Fujimuro, M. Kaposi’s Sarcoma-Associated Herpesvirus ORF21 Enhances the Phosphorylation of MEK and the Infectivity of Progeny Virus. Int. J. Mol. Sci. 2023, 24, 1238. [Google Scholar] [CrossRef]
- Tajima, S.; Nukui, Y.; Ito, M.; Takasaki, T.; Kurane, I. Nineteen nucleotides in the variable region of 3′ non-translated region are dispensable for the replication of dengue type 1 virus in vitro. Virus Res. 2006, 116, 38–44. [Google Scholar] [CrossRef]
- Butt, S.S.; Badshah, Y.; Shabbir, M.; Rafiq, M. Molecular Docking Using Chimera and Autodock Vina Software for Nonbioinformaticians. JMIR Bioinform. Biotech. 2020, 1, e14232. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 a (µM) | CC50 b (µM) | S.I. c |
|---|---|---|---|
| SOF | 8.31 | >100 | >12.0 |
| FMeU | >100 | >100 | - |
| ribavirin | 5.74 | >100 | >17.4 |
| Cell | Compound | IC50 a (µM) | CC50 b (µM) | S.I. c |
|---|---|---|---|---|
| BHK-21 | SOF | >100 | >100 | - |
| FMeU | >100 | >100 | - | |
| Huh7 | SOF | 8.34 | >100 | >12.0 |
| FMeU | >100 | >100 | - |
| Cell | Compound | IC50 a (µM) | CC50 b (µM) | S.I. c |
|---|---|---|---|---|
| Vero | SOF | >20 | >50 | - |
| Huh7 | SOF | 5.24 | >50 | >9.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurosawa, M.; Kato, F.; Hishiki, T.; Ito, S.; Fujisawa, H.; Yamaguchi, T.; Moriguchi, M.; Hosokawa, K.; Watanabe, T.; Saito-Tarashima, N.; et al. Sofosbuvir Suppresses the Genome Replication of DENV1 in Human Hepatic Huh7 Cells. Int. J. Mol. Sci. 2024, 25, 2022. https://doi.org/10.3390/ijms25042022
Kurosawa M, Kato F, Hishiki T, Ito S, Fujisawa H, Yamaguchi T, Moriguchi M, Hosokawa K, Watanabe T, Saito-Tarashima N, et al. Sofosbuvir Suppresses the Genome Replication of DENV1 in Human Hepatic Huh7 Cells. International Journal of Molecular Sciences. 2024; 25(4):2022. https://doi.org/10.3390/ijms25042022
Chicago/Turabian StyleKurosawa, Madoka, Fumihiro Kato, Takayuki Hishiki, Saori Ito, Hiroki Fujisawa, Tatsuo Yamaguchi, Misato Moriguchi, Kohei Hosokawa, Tadashi Watanabe, Noriko Saito-Tarashima, and et al. 2024. "Sofosbuvir Suppresses the Genome Replication of DENV1 in Human Hepatic Huh7 Cells" International Journal of Molecular Sciences 25, no. 4: 2022. https://doi.org/10.3390/ijms25042022
APA StyleKurosawa, M., Kato, F., Hishiki, T., Ito, S., Fujisawa, H., Yamaguchi, T., Moriguchi, M., Hosokawa, K., Watanabe, T., Saito-Tarashima, N., Minakawa, N., & Fujimuro, M. (2024). Sofosbuvir Suppresses the Genome Replication of DENV1 in Human Hepatic Huh7 Cells. International Journal of Molecular Sciences, 25(4), 2022. https://doi.org/10.3390/ijms25042022