
Sasanquasaponin from Camellia oleifera Abel Exerts an Anti-Inflammatory Effect in RAW 264.7 Cells via Inhibition of the NF-κB/MAPK Signaling Pathways
Abstract
:1. Introduction
2. Results
2.1. HPLC Analysis
2.2. Effects of SQS on RAW264.7 Cell Viability
2.3. SQS Reverses LPS-Induced ROS Generation
2.4. Effects of SQS on LPS-Induced Gene Expression of iNOS and COX-2
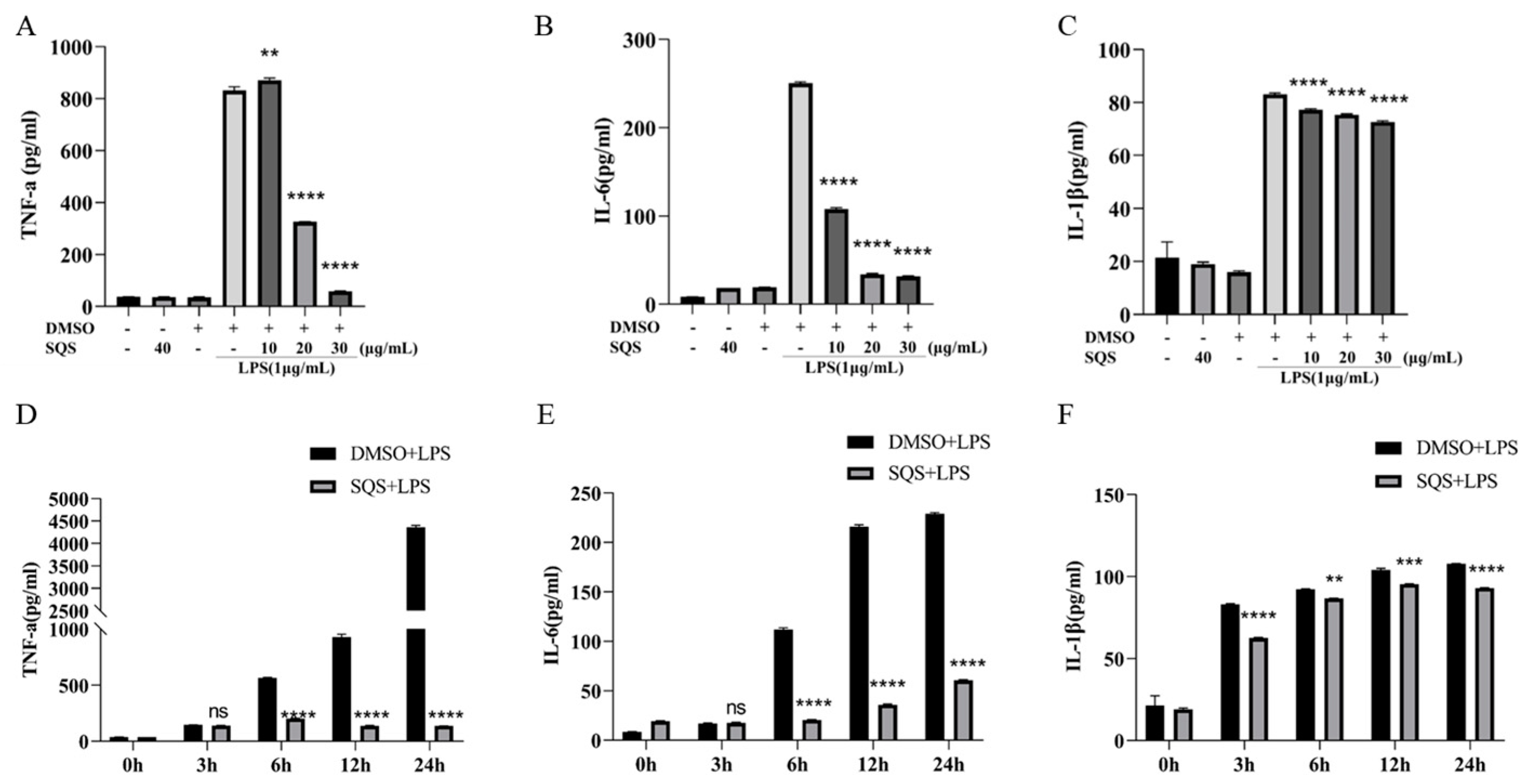
2.5. SQS Suppresses the Pro-Inflammatory Gene Expression in LPS-Treated RAW 264.7 Cells
2.6. SQS Inhibits Pro-Inflammatory Protein Expression in LPS-Treated RAW 264.7 Cells
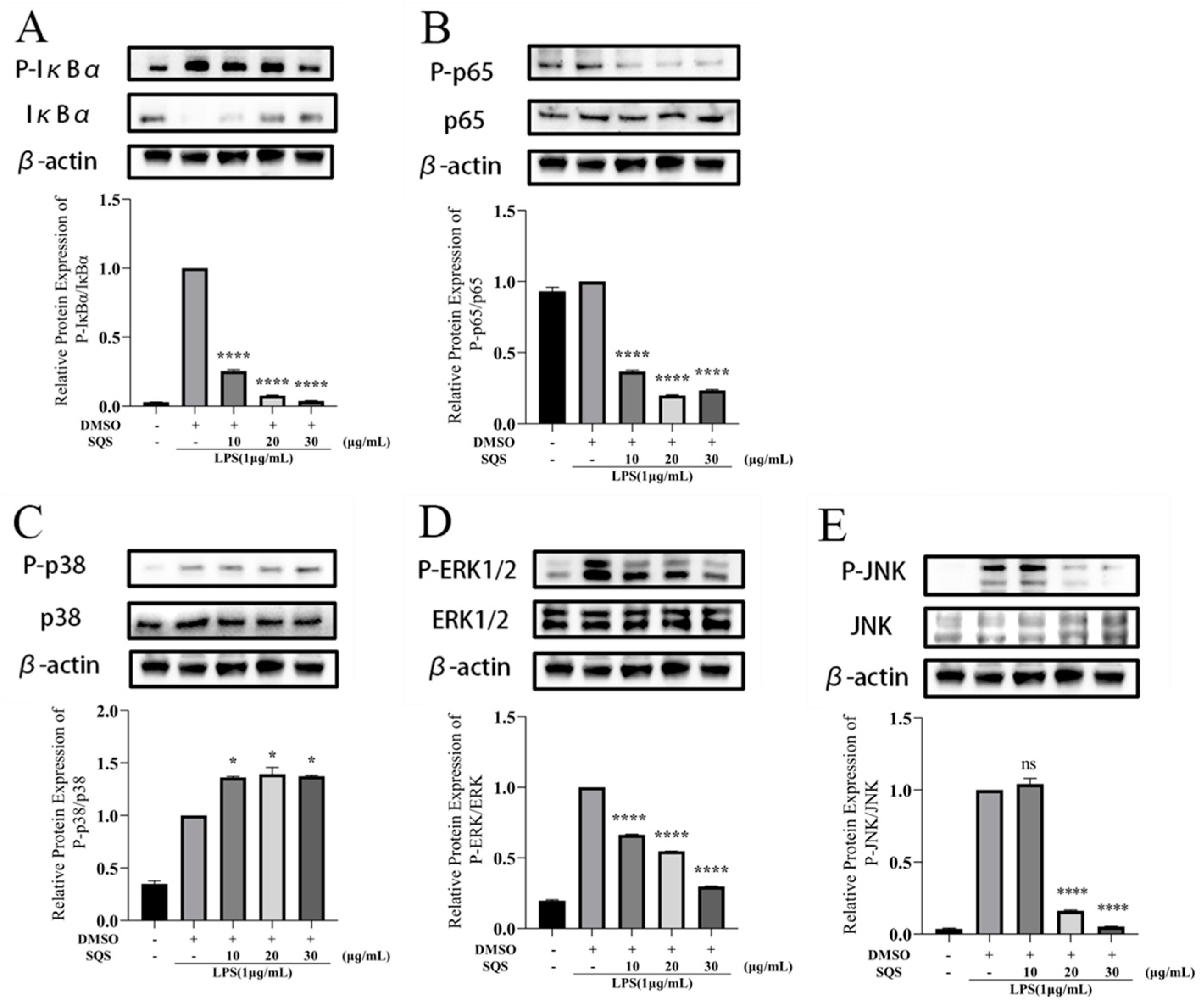
2.7. SQS Exerts Inflammatory Effects by Inhibiting NF-κB and MAPK Signaling
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of SQS
4.3. High-Performance Liquid Chromatography (HPLC) Analysis
4.4. Cell Culture
4.5. Analysis of Cell Viability
4.6. Determination of Intracellular ROS Production
4.7. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Camille, N.; Dealtry, G. Regulation of M1/M2 macrophage polarization by Sutherlandia frutescens via NFkB and MAPK signaling pathways. S. Afr. J. Bot. 2018, 116, 42–51. [Google Scholar] [CrossRef]
- Chen, H.P.; He, M.; Huang, Q.R.; Liu, D.; Huang, M. Sasanquasaponin protects rat cardiomyocytes against oxidative stress induced by anoxia-reoxygenation injury. Eur. J. Pharmacol. 2007, 575, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Yin, D.; Wang, W.; Zeng, G.; Liu, D.; Chen, H.; Huang, Q.; He, M. Cardioprotective effect of sasanquasaponin preconditioning via bradykinin-NO pathway in isolated rat heart. Phytother. Res. PTR 2009, 23, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Xiao, L.; Qiu, L.Y.; Yan, Y.F.; Wang, H.; Duan, G.L.; Liao, Z.P.; Chen, H.P. Sasanquasaponin-induced cardioprotection involves inhibition of mPTP opening via attenuating intracellular chloride accumulation. Fitoterapia 2017, 116, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-C.; Park, J.; Cho, S. Anti-Inflammatory and Anti-Oxidative Effects of luteolin-7-O-glucuronide in LPS-Stimulated Murine Macrophages through TAK1 Inhibition and Nrf2 Activation. Int. J. Mol. Sci. 2020, 21, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Collart, M.A.; Baeuerle, P.; Vassalli, P. Regulation of tumor necrosis factor alpha transcription in macrophages: Involvement of four kappa B-like motifs and of constitutive and inducible forms of NF-kappa B. Mol. Cell Biol. 1990, 10, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, R.D.; Ahmed, M.L.; Khalid, M.S.; Swamy, P. Anti-inflammatory activity of Madhuca longifolia seed saponin mixture. Pharm. Biol. 2009, 47, 592–597. [Google Scholar] [CrossRef]
- Huang, Q.; He, M.; Chen, H.; Shao, L.; Liu, D.; Luo, Y.; Dai, Y. Protective Effects of Sasanquasaponin on Injury of Endothelial Cells Induced by Anoxia and Reoxygenation in vitro. Basic Clin. Pharmacol. Toxicol. 2007, 101, 301–308. [Google Scholar] [CrossRef]
- Huang, Q.; Shao, L.; He, M.; Chen, H.; Liu, D.; Luo, Y.; Dai, Y. Inhibitory effects of sasanquasaponin on over-expression of ICAM-1 and on enhancement of capillary permeability induced by burns in rats. Burns 2005, 31, 637–642. [Google Scholar] [CrossRef]
- Hwang, J.H.; Lim, S.B. Antioxidant and Anti-inflammatory Activities of Broccoli Florets in LPS-stimulated RAW 264.7 Cells. Prev. Nutr. Food Sci. 2014, 19, 89–97. [Google Scholar] [CrossRef]
- Kang, J.H.; Sung, M.K.; Kawada, T.; Yoo, H.; Kim, Y.K.; Kim, J.S.; Yu, R. Soybean saponins suppress the release of proinflammatory mediators by LPS-stimulated peritoneal macrophages. Cancer Lett. 2005, 230, 219–227. [Google Scholar] [CrossRef]
- Khan, A.N.; Singh, R.; Bhattacharya, A.; Kumar, S.; Ghosh, A.; Nag, D.; Ravichandiran, V.; Ghosh, D. Glucogallin Attenuates RAW 264.7 Cells from Arsenic Trioxide Induced Toxicity via the NF-κB/NLRP3 Pathway. Molecules 2022, 27, 5263–5276. [Google Scholar] [CrossRef]
- Krementsov, D.N.; Thornton, T.M.; Teuscher, C.; Rincon, M. The Emerging Role of p38 Mitogen-Activated Protein Kinase in Multiple Sclerosis and Its Models. Mol. Cell. Biol. 2023, 33, 3728–3734. [Google Scholar] [CrossRef]
- Liu, Y.L.; Gao, Z.Z.; Guo, Q.T.; Wang, T.; Conger, L.; Chen, Y.; Sheng, Q.; Chen, J.; Nie, Z.M.; Zhang, Y.Z.; et al. Anti-Diabetic Effects of CTB-APSL Fusion Protein in Type 2 Diabetic Mice. Mar. Drugs 2014, 12, 1512–1529. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Ying, H.H.; Tong, F.D.; Zhang, C.; Quan, Y.P.; Zhang, Y.Z. Protective Effect of the Silkworm Protein 30Kc6 on Human Vascular Endothelial Cells Damaged by Oxidized Low Density Lipoprotein (Ox-LDL). PLoS ONE 2013, 8, e68746. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.N.; Xu, W.Z.; Ying, Q.; Ni, J.J.; Jia, X.Y.; Zhou, Y.R.; Ye, T.; Li, J.C.; Chen, K. Oncolytic Vaccinia Virus Carrying Aphrocallistes vastus Lectin (oncoVV-AVL) Enhances Inflammatory Response in Hepatocellular Carcinoma Cells. Mar. Drugs 2022, 20, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Le, H.T.T.; Park, J.; Ha, J.; Kusumaningrum, S.; Paik, J.H.; Cho, S. Synedrella nodiflora (Linn.) Gaertn. inhibits inflammatory responses through the regulation of Syk in RAW 264.7 macrophages. Exp. Ther. Med. 2020, 20, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, Y.Y.; Lee, J.; Jang, Y.J.; Jang, H.W. Chemical Composition, Antioxidant, and Anti-Inflammatory Activity of Essential Oil from Omija (Schisandra chinensis (Turcz.) Baill.) Produced by Supercritical Fluid Extraction Using CO2. Foods 2021, 10, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Hu, Y.; Yan, Y.; Zhou, W.; Chen, G.; Zeng, X.; Cao, Y. Characterization and Evaluation of Antioxidant and Anti-Inflammatory Activities of Flavonoids from the Fruits of Lycium barbarum. Foods 2022, 11, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Liu, P.; Shi, H.; Fan, W.; Feng, X.; Chen, J.; Jing, S.; Wang, L.; Zheng, Y.; Zhang, D.; et al. Identification of anti-inflammatory components in Dioscorea nipponica Makino based on HPLC-MS/MS, quantitative analysis of multiple components by single marker and chemometric methods. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2022, 1213, 123531. [Google Scholar] [CrossRef]
- Li, W.; Mao, Y.; Hua, B.; Gu, X.; Lu, C.; Xu, B.; Pan, W. Sasanquasaponin inhibited epithelial to mesenchymal transition in prostate cancer by regulating the PI3K/Akt/mTOR and Smad pathways. Pharm. Biol. 2022, 60, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xiao, X.; Ji, L.; Xie, L.; Wu, S.; Liu, Z. Camellia cake extracts reduce burn injury through suppressing inflammatory responses and enhancing collagen synthesis. Food Nutr. Res. 2020, 64, 3782–3797. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Park, E.-M.; Kim, D.-H.; Jung, K.; Jung, J.-S.; Lee, E.-J.; Hyun, J.-W.; Kang, J.L.; Kim, H.-S. Anti-inflammatory mechanism of ginseng saponins in activated microglia. J. Neuroimmunol. 2009, 209, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Lee, U.; Seo, S.; Wibowo, A.E.; Pongtuluran, O.B.; Lee, K.; Han, S.B.; Cho, S. Anti-inflammatory and antioxidant activities of methanol extract of Piper betle Linn. (Piper betle L.) leaves and stems by inhibiting NF-kappaB/MAPK/Nrf2 signaling pathways in RAW 264.7 macrophages. Biomed. Pharmacother. 2022, 155, 113734. [Google Scholar] [CrossRef] [PubMed]
- Shakhov, A.N.; Collart, M.A.; Vassalli, P.I.E.R.R.E.; Nedospasov, S.A.; Jongeneel, C.V. Kappa B-type enhancers are involved in lipopolysaccharide-mediated transcriptional activation of the tumor necrosis factor alpha gene in primary macrophages. J. Exp. Med. 1990, 171, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Su, X.D.; Jang, H.J.; Wang, C.Y.; Lee, S.W.; Rho, M.C.; Kim, Y.H.; Yang, S.Y. Anti-inflammatory Potential of Saponins from Aster tataricus via NF-κB/MAPK Activation. J. Nat. Prod. 2019, 82, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Chang, Y.; Wang, R.; Kang, Z.; Wang, Q.; Tong, Z.; Zhou, A.; Cui, C.; Liu, M. Optimization of ultrasound extraction of Tribulus terrestris L. leaves saponins and their HPLC-DAD-ESI-MS(n) profiling, anti-inflammatory activity and mechanism in vitro and in vivo. J. Ethnopharmacol. 2021, 278, 114225. [Google Scholar] [CrossRef]
- Truong, V.L.; Manochai, B.; Pham, T.T.; Jeong, W.S. Antioxidant and Anti-Inflammatory Activities of Zingiber montanum Oil in HepG2 Cells and Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. J. Med. Food 2021, 24, 595–605. [Google Scholar] [CrossRef]
- Xiao, X.; He, L.; Chen, Y.; Wu, L.; Wang, L.; Liu, Z. Anti-inflammatory and antioxidative effects of Camellia oleifera Abel components. Future Med. Chem. 2017, 9, 2069–2079. [Google Scholar] [CrossRef]
- Yang, C.-Y.; Hung, Y.-L.; Tang, K.-W.; Wang, S.-C.; Tseng, C.-H.; Tzeng, C.-C.; Liu, P.-L.; Li, C.-Y.; Chen, Y.-L. Discovery of 2-Substituted 3-Arylquinoline Derivatives as Potential Anti-Inflammatory Agents through Inhibition of LPS-Induced Inflammatory Responses in Macrophages. Molecules 2019, 24, 1162–1181. [Google Scholar] [CrossRef]
- Chen, J.J.; Peng, H.; Chen, C.J.; Wang, Y.; Sang, T.T.; Cai, Z.Q.; Zhao, Q.; Chen, S.J.; Lin, X.J.; Eling, T.E.; et al. NAG-1/GDF15 inhibits diabetic nephropathy via inhibiting AGE/RAGE-mediated inflammation signaling pathways in C57BL/6 mice and HK-2 cells. Life Sci. 2022, 311, 121142–121154. [Google Scholar] [CrossRef]
- Ye, Y.; Xing, H.; Chen, X. Anti-inflammatory and analgesic activities of the hydrolyzed sasanquasaponins from the defatted seeds of Camellia oleifera. Arch. Pharmacal Res. 2013, 36, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yan, H.; Wu, Y.; Wang, Y.; Xia, P. Quality Evaluation of the Oil of Camellia spp. Foods 2022, 11, 2221–2237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guo, Q.; Liang, Z.; Wang, M.; Wang, B.; Sun-Waterhouse, D.; Waterhouse, G.I.N.; Wang, J.; Ma, C.; Kang, W. Anti-inflammatory and antioxidant effects of Chaetoglobosin V(b) in LPS-induced RAW264.7 cells: Achieved via the MAPK and NF-kappaB signaling pathways. Food Chem. Toxicol. 2021, 147, 111915. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) |
|---|---|
| GAPDH-F | GTGGCAAAGTGGAGATTGTTG |
| GAPDH-R | CTCCTGGAAGATGGTGATGG |
| IL-6-F | CTGCAAGAGACTTCCATCCAG |
| IL-6-R | AGTGGTATAGACAGGTCTGTTGG |
| IL-1β-F | GAAATGCCACCTTTTGACAGTG |
| IL-1β-R | TGGATGCTCTCATCAGGACAG |
| TNF-α-F | CTGAACTTCGGGGTGATCGG |
| TNF-α-R | GGCTTGTCACTCGAATTTTGAGA |
| iNOS-F | GGA GCG AGT TGT GGA TTG TC |
| iNOS-R | GTG AGG GCT TGG CTG AGT GAG |
| COX-2-F | GAA GTC TTT GGT CTG GTG CCT G |
| COX-2-R | GTC TGC TGG TTT GGA ATA GTT GC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Zhao, N.; Kollie, L.; Yang, D.; Zhang, X.; Zhang, H.; Liang, Z. Sasanquasaponin from Camellia oleifera Abel Exerts an Anti-Inflammatory Effect in RAW 264.7 Cells via Inhibition of the NF-κB/MAPK Signaling Pathways. Int. J. Mol. Sci. 2024, 25, 2149. https://doi.org/10.3390/ijms25042149
Zhao Y, Zhao N, Kollie L, Yang D, Zhang X, Zhang H, Liang Z. Sasanquasaponin from Camellia oleifera Abel Exerts an Anti-Inflammatory Effect in RAW 264.7 Cells via Inhibition of the NF-κB/MAPK Signaling Pathways. International Journal of Molecular Sciences. 2024; 25(4):2149. https://doi.org/10.3390/ijms25042149
Chicago/Turabian StyleZhao, Yaxin, Nanshan Zhao, Larwubah Kollie, Dongfeng Yang, Xiaodan Zhang, Haihua Zhang, and Zongsuo Liang. 2024. "Sasanquasaponin from Camellia oleifera Abel Exerts an Anti-Inflammatory Effect in RAW 264.7 Cells via Inhibition of the NF-κB/MAPK Signaling Pathways" International Journal of Molecular Sciences 25, no. 4: 2149. https://doi.org/10.3390/ijms25042149