
Periosteum Containing Implicit Stem Cells: A Progressive Source of Inspiration for Bone Tissue Regeneration


Abstract
1. Introduction
2. Periosteum: Its Development and Structure
3. Periosteum Is a Promising Source of Skeletal Stem/Progenitor Cells
3.1. Identification and Useful Markers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp. | Sites | Ref. | ||
|---|---|---|---|---|
| Minimal criteria for defining MSCs | ||||
| CD105 m, h, r | +/− | tibia, femur, calvarial suture | [6,7,18,20,21,24,25,26,27,28,29,30,31] | |
| CD73 h | + | / | [18,27] | |
| CD90 (Thy1) m, h, r | +/− | tibia, femur, calvarial suture | [6,18,20,22,25,27,28,29,32] | |
| CD45 m, h, r | - | femur, phalange, tibia, calvarial suture | [6,7,18,19,20,21,24,25,26,27,28,30,31,32,33] | |
| CD34 m, h | −/low | femur, tibia | [7,18,24,27,28] | |
| CD14 h | - | tibia | [18,27,28] | |
| CD11b m, h | - | femur, tibia | [7,18,33] | |
| CD79α h | - | / | [18] | |
| CD19 h | - | / | [18,27] | |
| HLA-DR h | - | / | [18,27] | |
| Further SC/MSC Markers | ||||
| Sca1 m | +/− | femur, tibia, calvarial suture | [6,7,21,24,25,31,33] | |
| SSEA4 m | + | femur | [24] | |
| CD29 m, r | + | femur, tibia | [7,24,31,32] | |
| MSCA1 h | + | cranium | [34] | |
| CD51 m, h | +/low | tibia, femur | [6,25] | |
| PDGFRα (CD140α) m, h | +/−/low | tibia, femur | [6,7,20,25,30,31,33] | |
| PDGFRβ (CD140β) m | + | tibia, femur | [20,25,31,33] | |
| hSSC Markers | PDPN h, m, CD164 h, m | + | femur, phalange | [19] |
| CD146 h, m | +/−/low | femur, phalange, tibia, calvarial suture, cranium | [6,19,21,26,28,30,34] | |
| CD73 h, m | + | femur, phalange, tibia | [19,26,28] | |
| Nestin m | + | tibia, femur | [6,7,20] | |
| LepR m, h | +/− | tibia, femur | [6,7,20,30,33] | |
| Prx1 m | + | femur, tibia | [7] | |
| Cxcl12 m | + | femur, tibia | [7,30] | |
| Gremlin1 m | + | femur, tibia | [6,7,30] | |
| CD44 m, r | + | tibia, femur | [21,22,29] | |
| STRO1 m, h | + | tibia | [22,35] | |
| CD166 m, h | + | tibia | [22,26] | |
| CD49f m, h | +/low | tibia, femur | [6,21] | |
| BP1 (6C3) m, CD51 m, h | −/low | tibia, femur, calvarial suture | [6] | |
| CD200 m, h | +/− | tibia, femur, calvarial suture | [6,21,30] | |
| CD24 m | + | tibia | [21] | |
| CD106 h, CD13 h | + | tibia | [26] | |
| CD271 (LNGFR, p75) h | −/+ | tibia, femur | [26,30] | |
| ALP h | - | tibia | [26] | |
| Runx2 m, VCAM1 m | + | tibia | [30] | |
| Endothelial/Hematopoietic Markers | ||||
| CD133 m | - | femur | [24] | |
| CD31 m, h | −/low | femur, phalange, tibia, calvarial suture | [6,7,19,20,21,25,30,31,33] | |
| Ter119 m | - | tibia, femur, calvarial suture | [6,20,21,25,30,31,33] | |
| CD235a h | - | femur, phalange, tibia | [6,19,30] | |
| CD202b (Tie2) h | - | femur, phalange | [19] | |
| MECA32 (Plvap) m | + | tibia | [33] | |
| CD3 m, B220 m, Gr1 m | - | tibia | [33] | |
| CD20 h | - | tibia | [28] | |
| Pericyte Markers | ||||
| NG2 m | + | femur, tibia | [7] | |
| Fibroblast Markers | ||||
| Vimentin m | - | femur, tibia | [7] | |
| D7-FIB h | + | tibia | [26] | |
| Possible Specific Marks | ||||
| CtsK m, h | + | femur, tibia, calvarial suture | [6,21,22] | |
| Sox9 m | + | femur, tibia | [36,37] | |
| αSMA m, h | + | femur, tibia, calvarial suture | [30,33] | |
| Mx1m, h, CCR3/5 m, h | + | tibia, calvarial suture, femur | [30] | |
| Axin2 m | + | tibia | [38] | |
3.2. Biological Characterization
4. The Dominant Role of Periosteum in Fracture Healing
5. Molecular Signaling Pathways Regulating Periosteal Effects
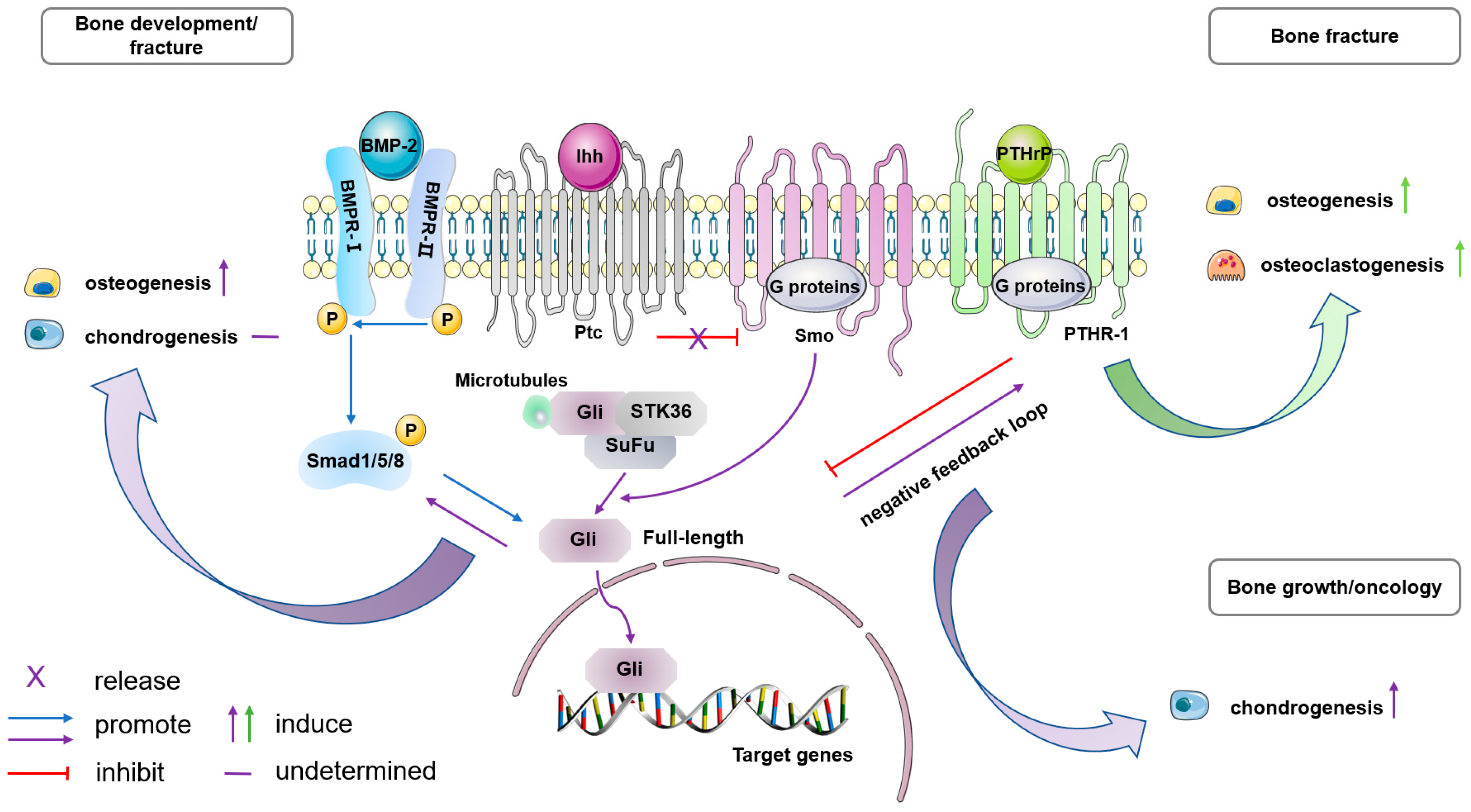
5.1. BMP Signaling
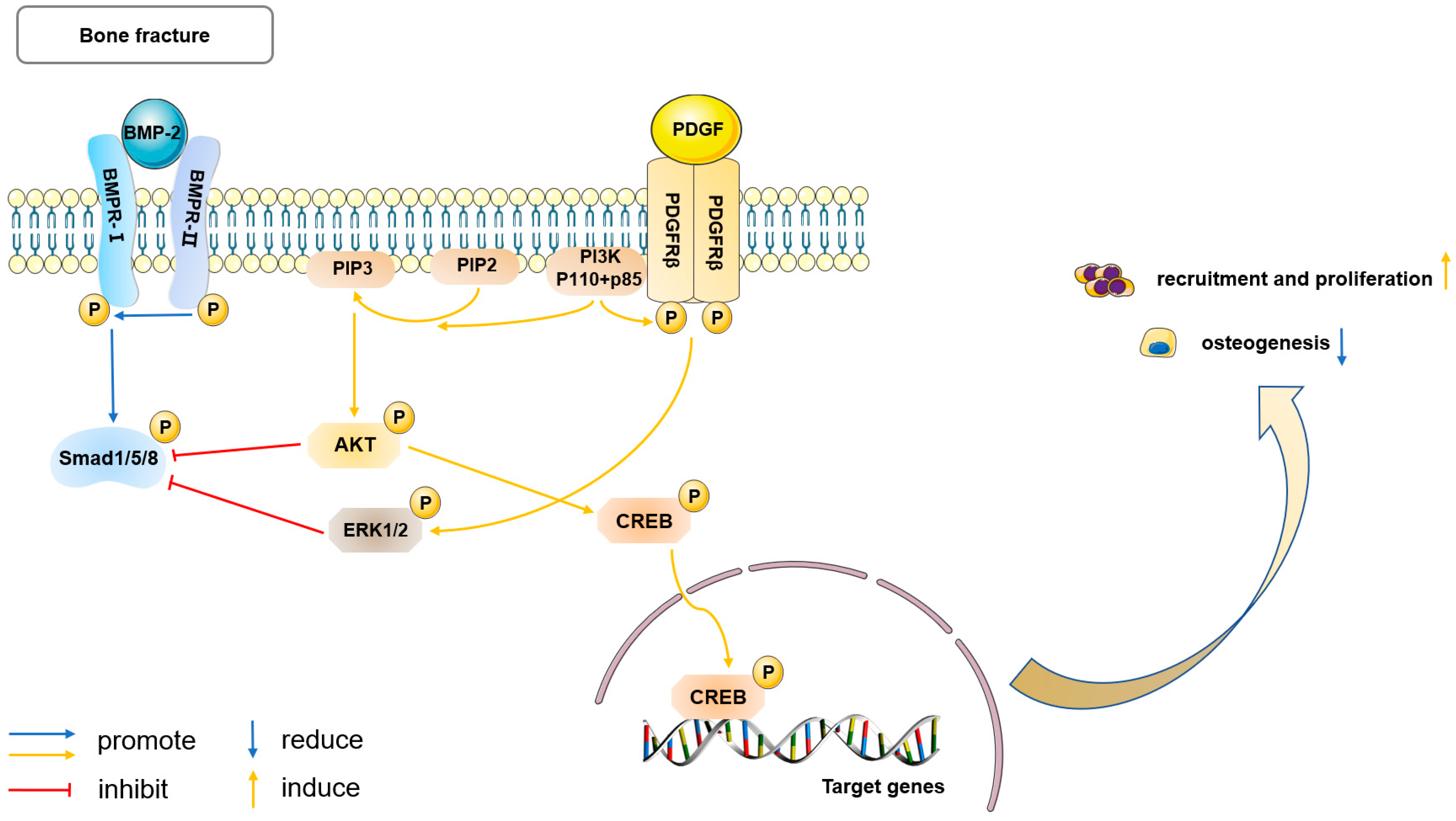
5.2. PDGF Signaling
5.3. Ihh Signaling
5.4. Biological Characterization
5.5. Notch Signaling
5.6. Other Related Molecular Regulation
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duhamel, H.L. Sur le developpement et la crue des os des animaux. Mem. Acad. Roy. DesSci. 1742, 55, 354–357. [Google Scholar]
- Ollier, L. Traite Experimentel et Clinique de la Regeneration des os et de la Production Artificielle du Tissu Osseux; V. Masson: Paris, France, 1867. [Google Scholar]
- Fell, H.B. The Osteogenic Capacity in vitro of Periosteum and Endosteum Isolated from the Limb Skeleton of Fowl Embryos and Young Chicks. J. Anat. 1932, 66 Pt 2, 157–180.11. [Google Scholar]
- Zhang, X.; Xie, C.; Lin, A.S.; Ito, H.; Awad, H.; Lieberman, J.R.; Rubery, P.T.; Schwarz, E.M.; O’Keefe, R.J.; Guldberg, R.E. Periosteal progenitor cell fate in segmental cortical bone graft transplantations: Implications for functional tissue engineering. J. Bone Miner. Res. 2005, 20, 2124–2137. [Google Scholar] [CrossRef]
- Dwek, J.R. The periosteum: What is it, where is it, and what mimics it in its absence? Skelet. Radiol. 2010, 39, 319–323. [Google Scholar] [CrossRef]
- Debnath, S.; Yallowitz, A.R.; McCormick, J.; Lalani, S.; Zhang, T.; Xu, R.; Li, N.; Liu, Y.; Yang, Y.S.; Eiseman, M.; et al. Discovery of a periosteal stem cell mediating intramembranous bone formation. Nature 2018, 562, 133–139. [Google Scholar] [CrossRef]
- Duchamp de Lageneste, O.; Julien, A.; Abou-Khalil, R.; Frangi, G.; Carvalho, C.; Cagnard, N.; Cordier, C.; Conway, S.J.; Colnot, C. Periosteum contains skeletal stem cells with high bone regenerative potential controlled by Periostin. Nat. Commun. 2018, 9, 773. [Google Scholar] [CrossRef]
- Colnot, C. Skeletal cell fate decisions within periosteum and bone marrow during bone regeneration. J. Bone Miner. Res. 2009, 24, 274–282. [Google Scholar] [CrossRef]
- Colnot, C.; Lu, C.; Hu, D.; Helms, J.A. Distinguishing the contributions of the perichondrium, cartilage, and vascular endothelium to skeletal development. Dev. Biol. 2004, 269, 55–69. [Google Scholar] [CrossRef]
- Berendsen, A.D.; Olsen, B.R. Bone development. Bone 2015, 80, 14–18. [Google Scholar] [CrossRef]
- Kronenberg, H.M. Developmental regulation of the growth plate. Nature 2003, 423, 332–336. [Google Scholar] [CrossRef]
- Maes, C.; Kobayashi, T.; Selig, M.K.; Torrekens, S.; Roth, S.I.; Mackem, S.; Carmeliet, G.; Kronenberg, H.M. Osteoblast precursors, but not mature osteoblasts, move into developing and fractured bones along with invading blood vessels. Dev. Cell 2010, 19, 329–344. [Google Scholar] [CrossRef]
- Allen, M.R.; Hock, J.M.; Burr, D.B. Periosteum: Biology, regulation, and response to osteoporosis therapies. Bone 2004, 35, 1003–1012. [Google Scholar] [CrossRef]
- Bisseret, D.; Kaci, R.; Lafage-Proust, M.H.; Alison, M.; Parlier-Cuau, C.; Laredo, J.D.; Bousson, V. Periosteum: Characteristic imaging findings with emphasis on radiologic-pathologic comparisons. Skelet. Radiol. 2015, 44, 321–338. [Google Scholar] [CrossRef]
- Hsiao, H.Y.; Yang, C.Y.; Liu, J.W.; Brey, E.M.; Cheng, M.H. Periosteal Osteogenic Capacity Depends on Tissue Source. Tissue Eng. Part. A 2018, 24, 1733–1741. [Google Scholar] [CrossRef]
- Fan, W.; Crawford, R.; Xiao, Y. Structural and cellular differences between metaphyseal and diaphyseal periosteum in different aged rats. Bone 2008, 42, 81–89. [Google Scholar] [CrossRef]
- Allen, M.R.; Burr, D.B. Human femoral neck has less cellular periosteum, and more mineralized periosteum, than femoral diaphyseal bone. Bone 2005, 36, 311–316. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Chan, C.K.F.; Gulati, G.S.; Sinha, R.; Tompkins, J.V.; Lopez, M.; Carter, A.C.; Ransom, R.C.; Reinisch, A.; Wearda, T.; Murphy, M.; et al. Identification of the Human Skeletal Stem Cell. Cell 2018, 175, 43–56.e21. [Google Scholar] [CrossRef]
- Gao, B.; Deng, R.; Chai, Y.; Chen, H.; Hu, B.; Wang, X.; Zhu, S.; Cao, Y.; Ni, S.; Wan, M.; et al. Macrophage-lineage TRAP+ cells recruit periosteum-derived cells for periosteal osteogenesis and regeneration. J. Clin. Investig. 2019, 129, 2578–2594. [Google Scholar] [CrossRef]
- Han, Y.; Feng, H.; Sun, J.; Liang, X.; Wang, Z.; Xing, W.; Dai, Q.; Yang, Y.; Han, A.; Wei, Z.; et al. Lkb1 deletion in periosteal mesenchymal progenitors induces osteogenic tumors through mTORC1 activation. J. Clin. Investig. 2019, 129, 1895–1909. [Google Scholar] [CrossRef]
- Yang, W.; Neel, B.G. From an orphan disease to a generalized molecular mechanism: PTPN11 loss-of-function mutations in the pathogenesis of metachondromatosis. Rare Dis. 2013, 1, e26657. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Chen, A.; Lei, K. Analysis of Cellular Crosstalk and Molecular Signal between Periosteum-Derived Precursor Cells and Peripheral Cells During Bone Healing Process Using a Paper-Based Osteogenesis-On-A-Chip Platform. ACS Appl. Mater. Interfaces 2023, 15, 49051–49059. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, C.; Zeng, F.; Xue, M.; Zhang, X. Activation of the Hh pathway in periosteum-derived mesenchymal stem cells induces bone formation in vivo: Implication for postnatal bone repair. Am. J. Pathol. 2010, 177, 3100–3111. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Matthews, B.G.; Yu, J.; Novak, S.; Grcevic, D.; Sanjay, A.; Kalajzic, I. PDGF Modulates BMP2-Induced Osteogenesis in Periosteal Progenitor Cells. JBMR Plus 2019, 3, e10127. [Google Scholar] [CrossRef] [PubMed]
- De Bari, C.; Dell’Accio, F.; Vanlauwe, J.; Eyckmans, J.; Khan, I.M.; Archer, C.W.; Jones, E.A.; McGonagle, D.; Mitsiadis, T.A.; Pitzalis, C.; et al. Mesenchymal multipotency of adult human periosteal cells demonstrated by single-cell lineage analysis. Arthritis Rheum. 2006, 54, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, C.; Lucarini, G.; Andreoni, C.; Salvolini, E.; Bianchi, N.; Vozzi, G.; Gigante, A.; Mattioli-Belmonte, M. Human Periosteal Derived Stem Cell Potential: The Impact of age. Stem Cell Rev. Rep. 2015, 11, 487–500. [Google Scholar] [CrossRef]
- Roberts, S.J.; Geris, L.; Kerckhofs, G.; Desmet, E.; Schrooten, J.; Luyten, F.P. The combined bone forming capacity of human periosteal derived cells and calcium phosphates. Biomaterials 2011, 32, 4393–4405. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Ruan, Y.C.; Yu, M.K.; O’Laughlin, M.; Wise, H.; Chen, D.; Tian, L.; Shi, D.; Wang, J.; et al. Implant-derived magnesium induces local neuronal production of CGRP to improve bone-fracture healing in rats. Nat. Med. 2016, 22, 1160–1169. [Google Scholar] [CrossRef]
- Ortinau, L.C.; Wang, H.; Lei, K.; Deveza, L.; Jeong, Y.; Hara, Y.; Grafe, I.; Rosenfeld, S.B.; Lee, D.; Lee, B.; et al. Identification of Functionally Distinct Mx1+αSMA+ Periosteal Skeletal Stem Cells. Cell Stem Cell 2019, 25, 784–796.e5. [Google Scholar] [CrossRef]
- Doherty, L.; Yu, J.; Wang, X.; Hankenson, K.D.; Kalajzic, I.; Sanjay, A. A PDGFRβ-PI3K signaling axis mediates periosteal cell activation during fracture healing. PLoS ONE 2019, 14, e0223846. [Google Scholar] [CrossRef]
- Hayashi, O.; Katsube, Y.; Hirose, M.; Ohgushi, H.; Ito, H. Comparison of osteogenic ability of rat mesenchymal stem cells from bone marrow, periosteum, and adipose tissue. Calcif. Tissue Int. 2008, 82, 238–247. [Google Scholar] [CrossRef]
- Matthews, B.G.; Grcevic, D.; Wang, L.; Hagiwara, Y.; Roguljic, H.; Joshi, P.; Shin, D.G.; Adams, D.J.; Kalajzic, I. Analysis of αSMA-labeled progenitor cell commitment identifies notch signaling as an important pathway in fracture healing. J. Bone Miner. Res. 2014, 29, 1283–1294. [Google Scholar] [CrossRef]
- Umrath, F.; Thomalla, C.; Pöschel, S.; Schenke-Layland, K.; Reinert, S.; Alexander, D. Comparative Study of MSCA-1 and CD146 Isolated Periosteal Cell Subpopulations. Cell Physiol. Biochem. 2018, 51, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Ball, M.D.; Bonzani, I.C.; Bovis, M.J.; Williams, A.; Stevens, M.M. Human periosteum is a source of cells for orthopaedic tissue engineering: A pilot study. Clin. Orthop. Relat. Res. 2011, 469, 3085–3093. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Bougioukli, S.; Ortega, B.; Arevalo, E.; Lieberman, J.R.; McMahon, A.P. Sox9 positive periosteal cells in fracture repair of the adult mammalian long bone. Bone 2017, 103, 12–19. [Google Scholar] [CrossRef]
- Murao, H.; Yamamoto, K.; Matsuda, S.; Akiyama, H. Periosteal cells are a major source of soft callus in bone fracture. J. Bone Miner. Metab. 2013, 31, 390–398. [Google Scholar] [CrossRef]
- Ransom, R.C.; Hunter, D.J.; Hyman, S.; Singh, G.; Ransom, S.C.; Shen, E.Z.; Perez, K.C.; Gillette, M.; Li, J.; Liu, B.; et al. Axin2-expressing cells execute regeneration after skeletal injury. Sci. Rep. 2016, 6, 36524. [Google Scholar] [CrossRef]
- De Bari, C.; Dell’Accio, F.; Luyten, F.P. Human periosteum-derived cells maintain phenotypic stability and chondrogenic potential throughout expansion regardless of donor age. Arthritis Rheum. 2001, 44, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Eyckmans, J.; Roberts, S.J.; Schrooten, J.; Luyten, F.P. A clinically relevant model of osteoinduction: A process requiring calcium phosphate and BMP/Wnt signalling. J. Cell Mol. Med. 2010, 14, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- Bolander, J.; Chai, Y.C.; Geris, L.; Schrooten, J.; Lambrechts, D.; Roberts, S.J.; Luyten, F.P. Early BMP, Wnt and Ca2+/PKC pathway activation predicts the bone forming capacity of periosteal cells in combination with calcium phosphates. Biomaterials 2016, 86, 106–118. [Google Scholar] [CrossRef]
- Okamoto, K.; Nakashima, T.; Shinohara, M.; Negishi-Koga, T.; Komatsu, N.; Terashima, A.; Sawa, S.; Nitta, T.; Takayanagi, H. Osteoimmunology: The Conceptual Framework Unifying the Immune and Skeletal Systems. Physiol. Rev. 2017, 97, 1295–1349. [Google Scholar] [CrossRef]
- Roberts, S.J.; van Gastel, N.; Carmeliet, G.; Luyten, F.P. Uncovering the periosteum for skeletal regeneration: The stem cell that lies beneath. Bone 2015, 70, 10–18. [Google Scholar] [CrossRef]
- Iuchi, T.; Kusuhara, H.; Ueda, Y.; Morotomi, T.; Isogai, N. Influence of Periosteum Location on the Bone and Cartilage in Tissue-Engineered Phalanx. J. Hand Surg. Am. 2020, 45, 62.e1–62.e10. [Google Scholar] [CrossRef]
- Moore, E.; Maridas, D.; Gamer, L.; Chen, G.; Burton, K.; Rosen, V. A periosteum-derived cell line to study the role of BMP/TGFβ signaling in periosteal cell behavior and function. Front. Physiol. 2023, 14, 1221152. [Google Scholar] [CrossRef]
- van Gastel, N.; Stegen, S.; Eelen, G.; Schoors, S.; Carlier, A.; Daniëls, V.W.; Baryawno, N.; Przybylski, D.; Depypere, M.; Stiers, P.J.; et al. Lipid availability determines fate of skeletal progenitor cells via SOX9. Nature 2020, 579, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.J.; Chen, Y.; Moesen, M.; Schrooten, J.; Luyten, F.P. Enhancement of osteogenic gene expression for the differentiation of human periosteal derived cells. Stem Cell Res. 2011, 7, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, S.Y.; Lee, H.Y.; Lee, J.H.; Rho, G.J.; Lee, H.J.; Lee, H.C.; Byun, J.H.; Oh, S.H. Oxygen-Releasing Microparticles for Cell Survival and Differentiation Ability under Hypoxia for Effective Bone Regeneration. Biomacromolecules 2019, 20, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tower, R.J.; Chandra, A.; Yao, L.; Tong, W.; Xiong, Z.; Tang, K.; Zhang, Y.; Liu, X.S.; Boerckel, J.D.; et al. Periosteal Mesenchymal Progenitor Dysfunction and Extraskeletally-Derived Fibrosis Contribute to Atrophic Fracture Nonunion. J. Bone Miner. Res. 2019, 34, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Marsell, R.; Einhorn, T.A. The biology of fracture healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Dar, H.; Perrien, D.; Pal, S.; Stoica, A.; Uppuganti, S.; Nyman, J.; Jones, R.; Weitzmann, M.; Pacifici, R. Callus γδ T cells and microbe-induced intestinal Th17 cells improve fracture healing in mice. J. Clin. Investig. 2023, 133, e166577. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Xu, C.; Shi, H.; Cheng, Q. PTH1-34 improves bone healing by promoting angiogenesis and facilitating MSCs migration and differentiation in a stabilized fracture mouse model. PLoS ONE 2019, 14, e0226163. [Google Scholar] [CrossRef]
- Shi, Y.; He, G.; Lee, W.C.; McKenzie, J.A.; Silva, M.J.; Long, F. Gli1 identifies osteogenic progenitors for bone formation and fracture repair. Nat. Commun. 2017, 8, 2043. [Google Scholar] [CrossRef]
- Lin, Z.; Fateh, A.; Salem, D.M.; Intini, G. Periosteum: Biology and applications in craniofacial bone regeneration. J. Dent. Res. 2014, 93, 109–116. [Google Scholar] [CrossRef]
- Funes, S.C.; Rios, M.; Escobar-Vera, J.; Kalergis, A.M. Implications of macrophage polarization in autoimmunity. Immunology 2018, 154, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Li, M.; Chen, K.; Fang, B.; Chen, P.; Tang, Z.; Lin, X.; Fan, S. Periosteal matrix-derived hydrogel promotes bone repair through an early immune regulation coupled with enhanced angio- and osteogenesis. Biomaterials 2020, 227, 119552. [Google Scholar] [CrossRef] [PubMed]
- Yuling, T.; Xiao, C.; Junxia, Z.; Jun, J.; Xinghua, L. Effect of different composite plates on the healing of femoral fractures. J. Mech. Behav. Biomed. Mater. 2023, 151, 106356. [Google Scholar] [CrossRef] [PubMed]
- Leucht, P.; Kim, J.B.; Amasha, R.; James, A.W.; Girod, S.; Helms, J.A. Embryonic origin and Hox status determine progenitor cell fate during adult bone regeneration. Development 2008, 135, 2845–2854. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.G.; Novak, S.; Sbrana, F.V.; Funnell, J.L.; Cao, Y.; Buckels, E.J.; Grcevic, D.; Kalajzic, I. Heterogeneity of murine periosteum progenitors involved in fracture healing. eLife 2021, 10, e58534. [Google Scholar] [CrossRef]
- Chen, R.; Dong, H.; Raval, D.; Maridas, D.; Baroi, S.; Chen, K.; Hu, D.; Berry, S.; Baron, R.; Greenblatt, M.; et al. Sfrp4 is required to maintain Ctsk-lineage periosteal stem cell niche function. Proc. Natl. Acad. Sci. USA 2023, 120, e2312677120. [Google Scholar] [CrossRef] [PubMed]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef]
- Park, J.H.; Koh, E.B.; Seo, Y.J.; Oh, H.S.; Byun, J.H. BMP-9 Improves the Osteogenic Differentiation Ability over BMP-2 through p53 Signaling In Vitro in Human Periosteum-Derived Cells. Int. J. Mol. Sci. 2023, 24, 15252. [Google Scholar] [CrossRef]
- Salazar, V.S.; Capelo, L.P.; Cantù, C.; Zimmerli, D.; Gosalia, N.; Pregizer, S.; Cox, K.; Ohte, S.; Feigenson, M.; Gamer, L.; et al. Reactivation of a developmental Bmp2 signaling center is required for therapeutic control of the murine periosteal niche. eLife 2019, 8, e42386. [Google Scholar] [CrossRef]
- McColl, L.; Chen, X.; Solga, M.; Schlegel, K.; Haughey, S.; Lobo, P.; Fread, K.; Zunder, E.; Cha, R.; Park, S.; et al. BMP-6 promotes type 2 immune response during enhancement of rat mandibular bone defect healing. Front. Immunol. 2023, 14, 1064238. [Google Scholar] [CrossRef] [PubMed]
- Romero-Torrecilla, J.A.; Lamo-Espinosa, J.M.; Ripalda-Cemboráin, P.; López-Martínez, T.; Abizanda, G.; Riera-Álvarez, L.; de Galarreta-Moriones, S.R.; López-Barberena, A.; Rodríguez-Flórez, N.; Elizalde, R.; et al. An engineered periosteum for efficient delivery of rhBMP-2 and mesenchymal progenitor cells during bone regeneration. npj Regen. Med. 2023, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, C.; Xue, M.; Zhang, X. Expression of endogenous BMP-2 in periosteal progenitor cells is essential for bone healing. Bone 2011, 48, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Chappuis, V.; Gamer, L.; Cox, K.; Lowery, J.W.; Bosshardt, D.D.; Rosen, V. Periosteal BMP2 activity drives bone graft healing. Bone 2012, 51, 800–809. [Google Scholar] [CrossRef]
- Böhm, A.M.; Dirckx, N.; Tower, R.J.; Peredo, N.; Vanuytven, S.; Theunis, K.; Nefyodova, E.; Cardoen, R.; Lindner, V.; Voet, T.; et al. Activation of Skeletal Stem and Progenitor Cells for Bone Regeneration Is Driven by PDGFRβ Signaling. Devel-opmental cell. Developmental cell 2019, 51, 236–254. [Google Scholar] [CrossRef] [PubMed]
- Hung, B.P.; Hutton, D.L.; Kozielski, K.L.; Bishop, C.J.; Naved, B.; Green, J.J.; Caplan, A.I.; Gimble, J.M.; Dorafshar, A.H.; Grayson, W.L. Platelet-Derived Growth Factor BB Enhances Osteogenesis of Adipose-Derived but Not Bone Marrow-Derived Mesenchymal Stromal/Stem Cells. Stem Cells 2015, 33, 2773–2784. [Google Scholar] [CrossRef]
- Novak, S.; Madunic, J.; Shum, L.; Vucetic, M.; Wang, X.; Tanigawa, H.; Ghosh, M.; Sanjay, A.; Kalajzic, I. PDGF inhibits BMP2-induced bone healing. npj Regen. Med. 2023, 8, 3. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, K.H.; Shin, S.Y.; Koo, K.T.; Lee, Y.M.; Seol, Y.J. Dual delivery of rhPDGF-BB and bone marrow mesenchymal stromal cells expressing the BMP2 gene enhance bone formation in a critical-sized defect model. Tissue Eng. Part. A 2013, 19, 2495–2505. [Google Scholar] [CrossRef]
- Orikasa, S.; Matsushita, Y.; Manabe, H.; Fogge, M.; Lee, Z.; Mizuhashi, K.; Sakagami, N.; Ono, W.; Ono, N. Hedgehog activation promotes osteogenic fates of growth plate resting zone chondrocytes through transient clonal competency. JCI Insight 2024, 9, e165619. [Google Scholar] [CrossRef]
- Koosha, E.; Brenna, C.; Ashique, A.; Jain, N.; Ovens, K.; Koike, T.; Kitagawa, H.; Eames, B.F. Proteoglycan inhibition of canonical BMP-dependent cartilage maturation delays endochondral ossification. Development 2024, 151, dev201716. [Google Scholar] [CrossRef]
- Wang, M.; Nasiri, A.R.; Broadus, A.E.; Tommasini, S.M. Periosteal PTHrP Regulates Cortical Bone Remodeling During Fracture Healing. Bone 2015, 81, 104–111. [Google Scholar] [CrossRef]
- Jeffery, E.; Mann, T.; Pool, J.; Zhao, Z.; Morrison, S.J. Bone marrow and periosteal skeletal stem/progenitor cells make distinct contributions to bone maintenance and repair. Cell Stem Cell 2022, 29, 1547–1561.e6. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, S.T.; Serowoky, M.A.; Vakhshori, V.; Tripuraneni, N.; Hegde, N.V.; Lieberman, J.R.; Crump, J.G.; Mariani, F.V. Sox9+ messenger cells orchestrate large-scale skeletal regeneration in the mammalian rib. eLife 2019, 8, e40715. [Google Scholar] [CrossRef] [PubMed]
- Tsukasaki, M.; Komatsu, N.; Negishi-Koga, T.; Huynh, N.; Muro, R.; Ando, Y.; Seki, Y.; Terashima, A.; Pluemsakunthai, W.; Nitta, T.; et al. Periosteal stem cells control growth plate stem cells during postnatal skeletal growth. Nat. Commun. 2022, 13, 4166. [Google Scholar] [CrossRef] [PubMed]
- Chijimatsu, R.; Saito, T. Mechanisms of synovial joint and articular cartilage development. Cell. Mol. Life Sci. 2019, 76, 3939–3952. [Google Scholar] [CrossRef] [PubMed]
- Joeng, K.S.; Long, F. Wnt7b can replace Ihh to induce hypertrophic cartilage vascularization but not osteoblast differentiation during endochondral bone development. Bone Res. 2014, 2, 14004. [Google Scholar] [CrossRef] [PubMed]
- Leucht, P.; Lee, S.; Yim, N. Wnt signaling and bone regeneration: Can’t have one without the other. Biomaterials 2019, 196, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Youngstrom, D.W.; Dishowitz, M.I.; Bales, C.B.; Carr, E.; Mutyaba, P.L.; Kozloff, K.M.; Shitaye, H.; Hankenson, K.D.; Loomes, K.M. Jagged1 expression by osteoblast-lineage cells regulates trabecular bone mass and periosteal expansion in mice. Bone 2016, 91, 64–74. [Google Scholar] [CrossRef]
- Lawal, R.A.; Zhou, X.; Batey, K.; Hoffman, C.M.; Georger, M.A.; Radtke, F.; Hilton, M.J.; Xing, L.; Frisch, B.J.; Calvi, L.M. The Notch Ligand Jagged1 Regulates the Osteoblastic Lineage by Maintaining the Osteoprogenitor Pool. J. Bone Miner. Res. 2017, 32, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Olsen, B.R. Osteoblast-derived VEGF regulates osteoblast differentiation and bone formation during bone repair. J. Clin. Investig. 2016, 126, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Buettmann, E.G.; McKenzie, J.A.; Migotsky, N.; Sykes, D.A.; Hu, P.; Yoneda, S.; Silva, M.J. VEGFA From Early Osteoblast Lineage Cells (Osterix+) Is Required in Mice for Fracture Healing. J. Bone Miner. Res. 2019, 34, 1690–1706. [Google Scholar] [CrossRef] [PubMed]
- Hah, Y.S.; Jun, J.S.; Lee, S.G.; Park, B.W.; Kim, D.R.; Kim, U.K.; Kim, J.R.; Byun, J.H. Vascular endothelial growth factor stimulates osteoblastic differentiation of cultured human periosteal-derived cells expressing vascular endothelial growth factor receptors. Mol. Biol. Rep. 2011, 38, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Bravo, D.; Josephson, A.M.; Bradaschia-Correa, V.; Wong, M.Z.; Yim, N.L.; Neibart, S.S.; Lee, S.N.; Huo, J.; Coughlin, T.; Mizrahi, M.M.; et al. Temporary inhibition of the plasminogen activator inhibits periosteal chondrogenesis and promotes periosteal osteogenesis during appendicular bone fracture healing. Bone 2018, 112, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Granero-Moltó, F.; Weis, J.A.; Miga, M.I.; Landis, B.; Myers, T.J.; O’Rear, L.; Longobardi, L.; Jansen, E.D.; Mortlock, D.P.; Spagnoli, A. Regenerative effects of transplanted mesenchymal stem cells in fracture healing. Stem Cells 2009, 27, 1887–1898. [Google Scholar] [CrossRef]
- Arron, J.R.; Choi, Y. Bone versus immune system. Nature 2000, 408, 535–536. [Google Scholar] [CrossRef]
- Ito, R.; Matsumiya, T.; Kon, T.; Narita, N.; Kubota, K.; Sakaki, H.; Ozaki, T.; Imaizumi, T.; Kobayashi, W.; Kimura, H. Periosteum-derived cells respond to mechanical stretch and activate Wnt and BMP signaling pathways. Biomed. Res. 2014, 35, 69–79. [Google Scholar] [CrossRef]
- Schönmeyr, B.; Clavin, N.; Avraham, T.; Longo, V.; Mehrara, B.J. Synthesis of a tissue-engineered periosteum with acellular dermal matrix and cultured mesenchymal stem cells. Tissue Eng. Part. A 2009, 15, 1833–1841. [Google Scholar] [CrossRef]
- Hoffman, M.D.; Xie, C.; Zhang, X.; Benoit, D.S. The effect of mesenchymal stem cells delivered via hydrogel-based tissue engineered periosteum on bone allograft healing. Biomaterials 2013, 34, 8887–8898. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Ren, L.; Yang, Y. Engineering vascularized bone grafts by integrating a biomimetic periosteum and β-TCP scaffold. ACS Appl. Mater. Interfaces 2014, 6, 9622–9633. [Google Scholar] [CrossRef]
- Shi, X.; Chen, S.; Zhao, Y.; Lai, C.; Wu, H. Enhanced osteogenesis by a biomimic pseudo-periosteum-involved tissue engineering strategy. Adv. Healthc. Mater. 2013, 2, 1229–1235. [Google Scholar] [CrossRef]
- Shi, X.; Fujie, T.; Saito, A.; Takeoka, S.; Hou, Y.; Shu, Y.; Chen, M.; Wu, H. Periosteum-mimetic structures made from freestanding microgrooved nanosheets. Adv. Mater. 2014, 26, 3290–3296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Filion, T.M.; Kutikov, A.B.; Song, J. Facile Stem Cell Delivery to Bone Grafts Enabled by Smart Shape Recovery and Stiffening of Degradable Synthetic Periosteal Membranes. Adv. Funct. Mater. 2016, 27, 1604784. [Google Scholar] [CrossRef]
- Gong, M.; Chi, C.; Ye, J.; Liao, M.; Xie, W.; Wu, C.; Shi, R.; Zhang, L. Icariin-loaded electrospun PCL/gelatin nanofiber membrane as potential artificial periosteum. Colloids Surf. B Biointerfaces 2018, 170, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhai, Y.; Nuzzo, M.; Yang, X.; Yang, Y.; Zhang, X. Layer-by-layer nanofiber-enabled engineering of biomimetic periosteum for bone repair and reconstruction. Biomaterials 2018, 182, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Gu, Y.; Liu, L.; Tang, J.; Mao, J.; Xi, K.; Jiang, Z.; Zhou, Y.; Xu, Y.; Deng, L.; et al. Hierarchical micro/nanofibrous membranes of sustained releasing VEGF for periosteal regeneration. Biomaterials 2020, 227, 119555. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Deng, C.; Qi, S. Periosteum Containing Implicit Stem Cells: A Progressive Source of Inspiration for Bone Tissue Regeneration. Int. J. Mol. Sci. 2024, 25, 2162. https://doi.org/10.3390/ijms25042162
Zhang X, Deng C, Qi S. Periosteum Containing Implicit Stem Cells: A Progressive Source of Inspiration for Bone Tissue Regeneration. International Journal of Molecular Sciences. 2024; 25(4):2162. https://doi.org/10.3390/ijms25042162
Chicago/Turabian StyleZhang, Xinyuan, Chen Deng, and Shengcai Qi. 2024. "Periosteum Containing Implicit Stem Cells: A Progressive Source of Inspiration for Bone Tissue Regeneration" International Journal of Molecular Sciences 25, no. 4: 2162. https://doi.org/10.3390/ijms25042162
APA StyleZhang, X., Deng, C., & Qi, S. (2024). Periosteum Containing Implicit Stem Cells: A Progressive Source of Inspiration for Bone Tissue Regeneration. International Journal of Molecular Sciences, 25(4), 2162. https://doi.org/10.3390/ijms25042162