
Alterations in Genome Organization in Lymphoma Cell Nuclei due to the Presence of the t(14;18) Translocation


 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Results
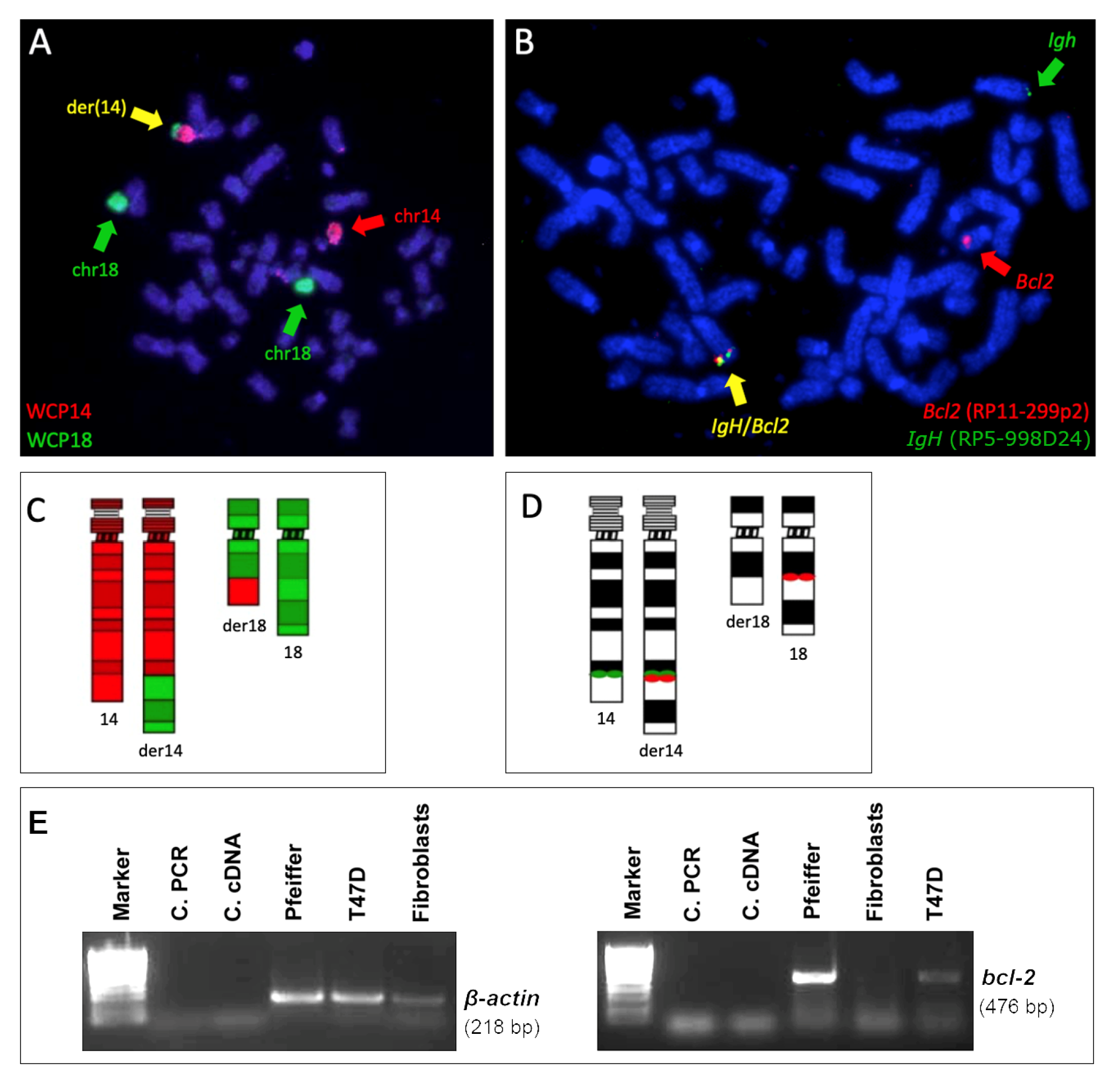
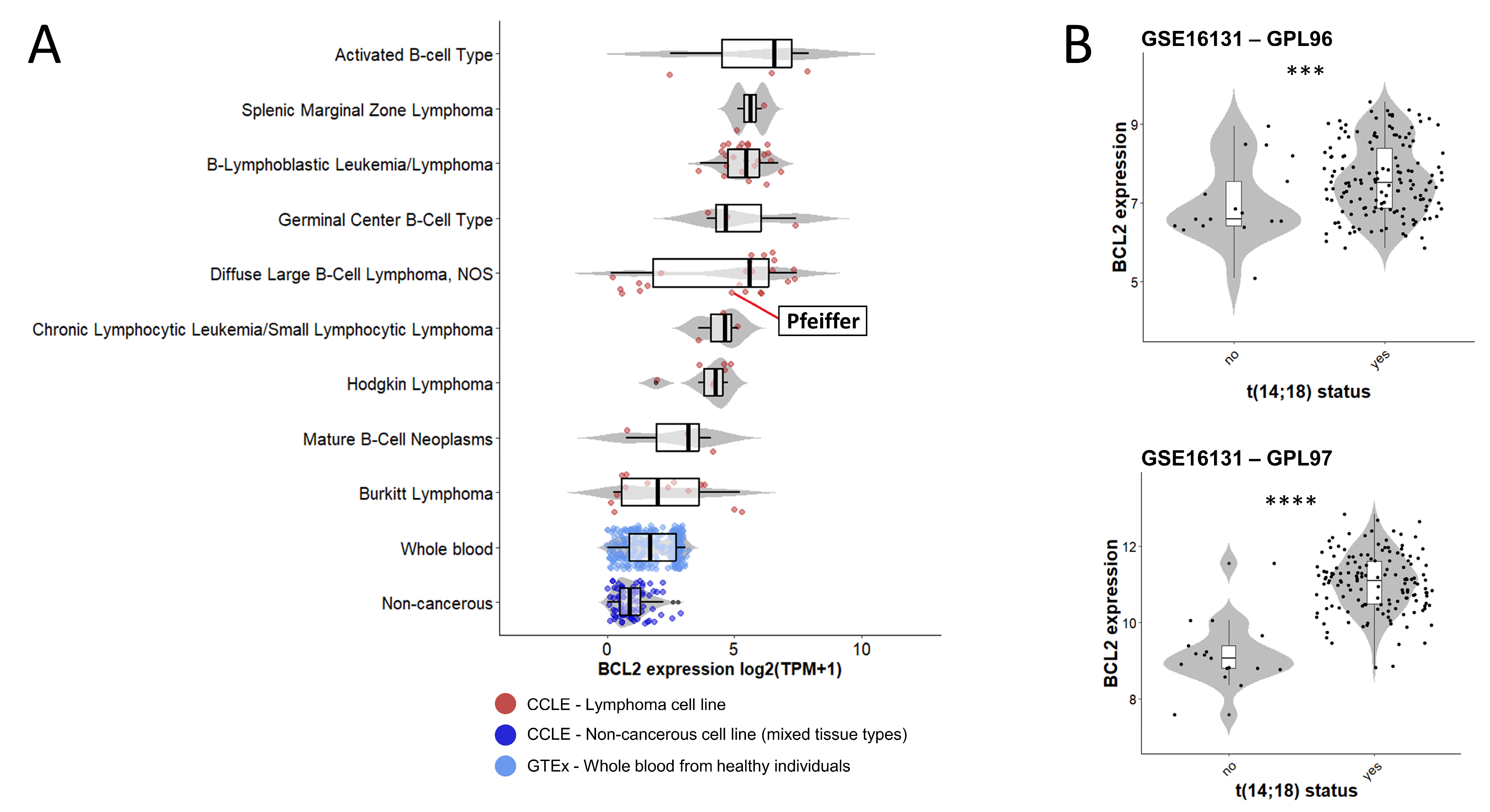
2.1. t(14;18) Translocation and BCL2 Expression in the Pfeiffer Cell Line
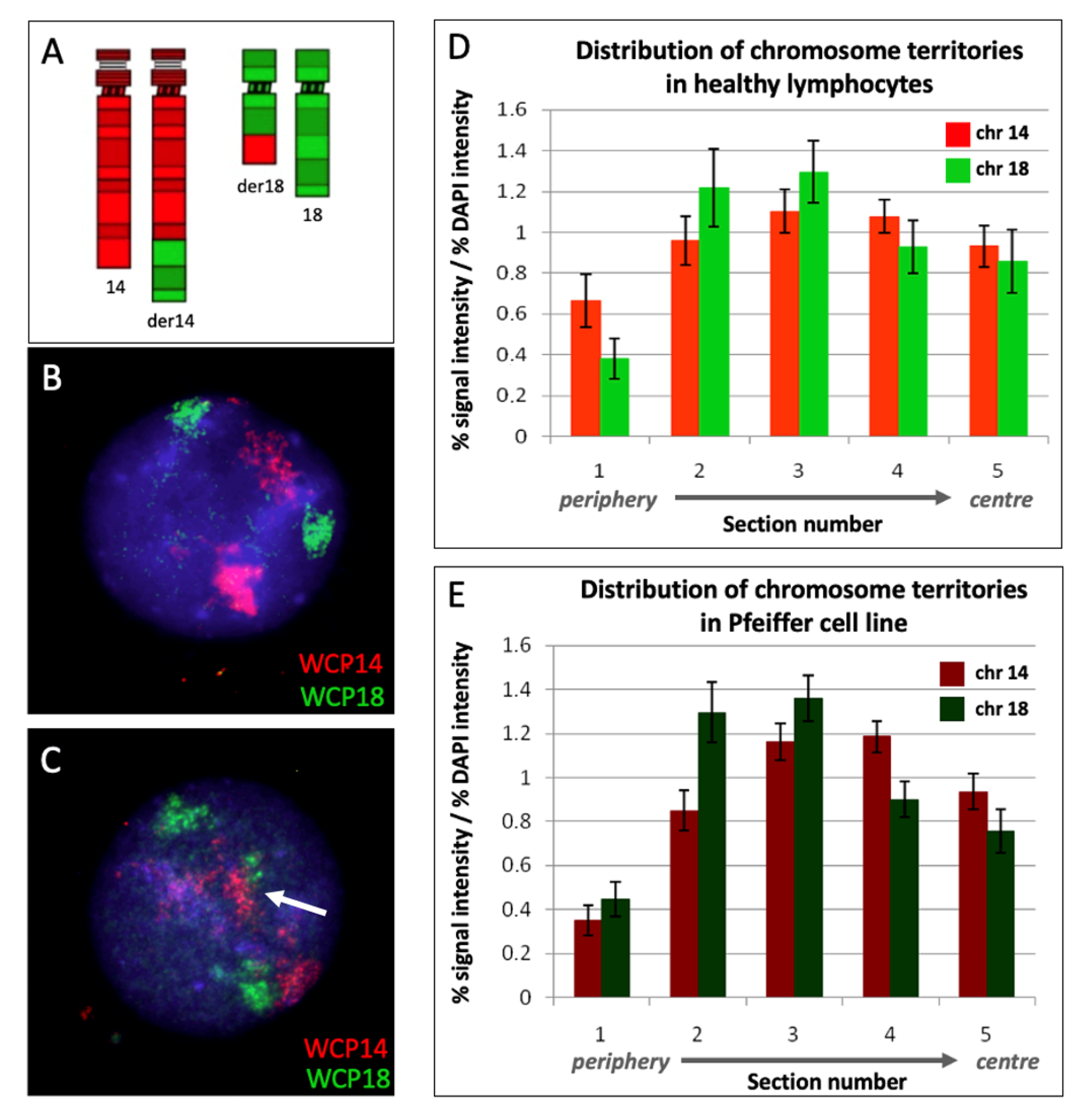
2.2. Chromosome Territories 14 and 18 Are Unaltered in the Pfeiffer Cell Line Compared to Healthy Lymphocytes
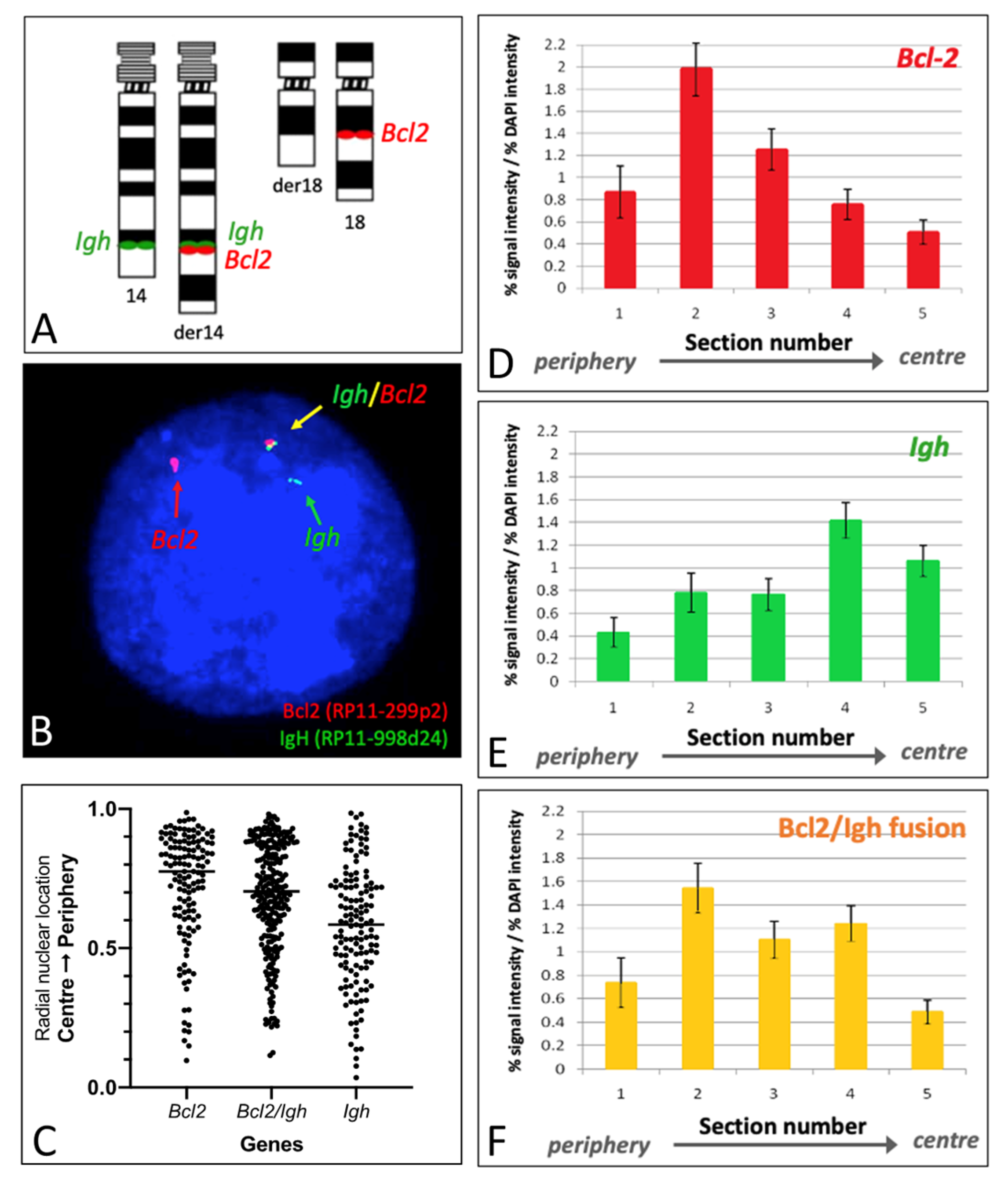
2.3. Nuclear Positioning of BCL2, IGH, and BCL2::IGH
2.4. Gene Nuclear Positioning in Relation to Their Corresponding Chromosomal Territories
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Expression Analysis of Publicly Available Data
4.3. Control Samples
4.4. Reverse Transcription (RT) PCR
4.5. Fluorescence In Situ Hybridization (FISH) on Metaphase Chromosomes and Interphase Nuclei
4.6. Nuclear Localization Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D. The 2016 Revision of the World Health Organization Classification of Lymphoid Neoplasms. Blood J. Am. Soc. Hematol. 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, E. Follicular Lymphoma: 2023 Update on Diagnosis and Management. Am. J. Hematol. 2022, 97, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Katsuyoshi, T. Highlights: Follicular Lymphoma. J. Clin. Exp. Hematop. 2023, 63, 212–213. [Google Scholar]
- Küppers, R. Mechanisms of B-Cell Lymphoma Pathogenesis. Nat. Rev. Cancer 2005, 5, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Egle, A.; Harris, A.W.; Bath, M.L.; O’Reilly, L.; Cory, S. VavP-Bcl2 Transgenic Mice Develop Follicular Lymphoma Preceded by Germinal Center Hyperplasia. Blood 2004, 103, 2276–2283. [Google Scholar] [CrossRef] [PubMed]
- Bonzheim, I.; Salaverria, I.; Haake, A.; Gastl, G.; Adam, P.; Siebert, R.; Fend, F.; Quintanilla-Martinez, L. A Unique Case of Follicular Lymphoma Provides Insights to the Clonal Evolution from Follicular Lymphoma in Situ to Manifest Follicular Lymphoma. Blood J. Am. Soc. Hematol. 2011, 118, 3442–3444. [Google Scholar] [CrossRef] [PubMed]
- Devan, J.; Janikova, A.; Mraz, M. New Concepts in Follicular Lymphoma Biology: From BCL2 to Epigenetic Regulators and Non-Coding RNAs. Semin. Oncol. 2018, 45, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Mozas, P.; López, C.; Grau, M.; Nadeu, F.; Clot, G.; Valle, S.; Kulis, M.; Navarro, A.; Ramis-Zaldivar, J.E.; González-Farré, B. Genomic Landscape of Follicular Lymphoma Across a Wide Spectrum of Clinical Behaviors. Hematol. Oncol. 2023, 41, 631–643. [Google Scholar] [CrossRef]
- Federico, C.; Bruno, F.; Ragusa, D.; Clements, C.S.; Brancato, D.; Henry, M.P.; Bridger, J.M.; Tosi, S.; Saccone, S. Chromosomal Rearrangements and Altered Nuclear Organization: Recent Mechanistic Models in Cancer. Cancers 2021, 13, 5860. [Google Scholar] [CrossRef]
- Bridger, J.M.; Pereira, R.T.; Pina, C.; Tosi, S.; Lewis, A. Alterations to Genome Organisation in Stem Cells, Their Differentiation and Associated Diseases. In Nuclear, Chromosomal, and Genomic Architecture in Biology and Medicine; Springer: Berlin/Heidelberg, Germany, 2022; pp. 71–102. [Google Scholar]
- Fritz, A.J.; Sehgal, N.; Pliss, A.; Xu, J.; Berezney, R. Chromosome Territories and the Global Regulation of the Genome. Genes Chromosomes Cancer 2019, 58, 407–426. [Google Scholar] [CrossRef]
- Parada, L.A.; Sotiriou, S.; Misteli, T. Spatial Genome Organization. Exp. Cell Res. 2004, 296, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.; Grosschedl, R. Dynamics and Interplay of Nuclear Architecture, Genome Organization, and Gene Expression. Genes Dev. 2007, 21, 3027–3043. [Google Scholar] [CrossRef] [PubMed]
- Bickmore, W.A.; Van Steensel, B. Genome Architecture: Domain Organization of Interphase Chromosomes. Cell 2013, 152, 1270–1284. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.M.; Arican-Gotkas, H.D.; Foster, H.A.; Godwin, L.S.; Harvey, A.; Kill, I.R.; Knight, M.; Mehta, I.S.; Ahmed, M.H. The Non-Random Repositioning of Whole Chromosomes and Individual Gene Loci in Interphase Nuclei and its Relevance in Disease, Infection, Aging, and Cancer. Adv. Exp. Med. Biol. 2014, 773, 263–279. [Google Scholar] [PubMed]
- Szczepińska, T.; Rusek, A.M.; Plewczynski, D. Intermingling of Chromosome Territories. Genes Chromosomes Cancer 2019, 58, 500–506. [Google Scholar] [CrossRef]
- Finlan, L.E.; Sproul, D.; Thomson, I.; Boyle, S.; Kerr, E.; Perry, P.; Ylstra, B.; Chubb, J.R.; Bickmore, W.A. Recruitment to the Nuclear Periphery can Alter Expression of Genes in Human Cells. PLoS Genet. 2008, 4, e1000039. [Google Scholar] [CrossRef]
- Guerreiro, I.; Kind, J. Spatial Chromatin Organization and Gene Regulation at the Nuclear Lamina. Curr. Opin. Genet. Dev. 2019, 55, 19–25. [Google Scholar] [CrossRef]
- Lemaître, C.; Bickmore, W.A. Chromatin at the Nuclear Periphery and the Regulation of Genome Functions. Histochem. Cell Biol. 2015, 144, 111–122. [Google Scholar] [CrossRef]
- Kumar, Y.; Sengupta, D.; Bickmore, W.A. Recent Advances in the Spatial Organization of the Mammalian Genome. J. Biosci. 2020, 45, 1–15. [Google Scholar] [CrossRef]
- Tanabe, H.; Müller, S.; Neusser, M.; von Hase, J.; Calcagno, E.; Cremer, M.; Solovei, I.; Cremer, C.; Cremer, T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc. Natl. Acad. Sci. USA 2022, 99, 4424–4429. [Google Scholar] [CrossRef] [PubMed]
- Federico, C.; Owoka, T.; Ragusa, D.; Sturiale, V.; Caponnetto, D.; Leotta, C.G.; Bruno, F.; Foster, H.A.; Rigamonti, S.; Giudici, G. Deletions of Chromosome 7q Affect Nuclear Organization and HLXB9 Gene Expression in Hematological Disorders. Cancers 2019, 11, 585. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, D.; Cicirò, Y.; Federico, C.; Saccone, S.; Bruno, F.; Saeedi, R.; Sisu, C.; Pina, C.; Sala, A.; Tosi, S. Engineered Model of T (7; 12)(q36; p13) AML Recapitulates Patient-Specific Features and Gene Expression Profiles. Oncogenesis 2022, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Leich, E.; Salaverria, I.; Bea, S.; Zettl, A.; Wright, G.; Moreno, V.; Gascoyne, R.D.; Chan, W.; Braziel, R.M.; Rimsza, L.M. Follicular Lymphomas with and without Translocation T (14; 18) Differ in Gene Expression Profiles and Genetic Alterations. Blood J. Am. Soc. Hematol. 2009, 114, 826–834. [Google Scholar]
- Clements, C.S.; Bikkul, U.; Ahmed, M.H.; Foster, H.A.; Godwin, L.S.; Bridger, J.M. Visualizing the Spatial Relationship of the Genome with the Nuclear Envelope using Fluorescence in Situ Hybridization. Nucl. Envel. Methods Protoc. 2016, 1411, 387–406. [Google Scholar]
- Gabay, C.; Ben-Bassat, H.; Schlesinger, M.; Laskov, R. Somatic Mutations and Intraclonal Variations in the Rearranged Vk Genes of B-non-Hodgkin’s Lymphoma Cell Lines. Eur. J. Haematol. 1999, 63, 180–191. [Google Scholar] [CrossRef]
- Taslerová, R.; Kozubek, S.; Lukášová, E.; Jirsová, P.; Bártová, E.; Kozubek, M. Arrangement of Chromosome 11 and 22 Territories, EWSR1 and FLI1 Genes, and Other Genetic Elements of these Chromosomes in Human Lymphocytes and Ewing Sarcoma Cells. Hum. Genet. 2003, 112, 143–155. [Google Scholar] [CrossRef]
- Ballabio, E.; Cantarella, C.D.; Federico, C.; Di Mare, P.; Hall, G.; Harbott, J.; Hughes, J.; Saccone, S.; Tosi, S. Ectopic Expression of the HLXB9 Gene is Associated with an Altered Nuclear Position in T (7; 12) Leukaemias. Leukemia 2009, 23, 1179–1182. [Google Scholar] [CrossRef]
- Lukášová, E.; Kozubek, S.; Kozubek, M.; Kjeronská, J.; Rýznar, L.; Horáková, J.; Krahulcová, E.; Horneck, G. Localisation and Distance between ABL and BCR Genes in Interphase Nuclei of Bone Marrow Cells of Control Donors and Patients with Chronic Myeloid Leukaemia. Hum. Genet. 1997, 100, 525–535. [Google Scholar] [CrossRef]
- Volpi, E.V.; Chevret, E.; Jones, T.; Vatcheva, R.; Williamson, J.; Beck, S.; Campbell, R.D.; Goldsworthy, M.; Powis, S.H.; Ragoussis, J. Large-Scale Chromatin Organization of the Major Histocompatibility Complex and Other Regions of Human Chromosome 6 and its Response to Interferon in Interphase Nuclei. J. Cell. Sci. 2000, 113, 1565–1576. [Google Scholar] [CrossRef]
- Morgan, G.T. Imaging the Dynamics of Transcription Loops in Living Chromosomes. Chromosoma 2018, 127, 361–374. [Google Scholar] [CrossRef]
- Gabriele, M.; Brandão, H.B.; Grosse-Holz, S.; Jha, A.; Dailey, G.M.; Cattoglio, C.; Hsieh, T.S.; Mirny, L.; Zechner, C.; Hansen, A.S. Dynamics of CTCF-and Cohesin-Mediated Chromatin Looping Revealed by Live-Cell Imaging. Science 2022, 376, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S. A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef]
- Karasu, N.; Sexton, T. 4C-Seq: Interrogating Chromatin Looping with Circular Chromosome Conformation Capture. Capturing Chromosome Conform. Methods Protoc. 2021, 2157, 19–34. [Google Scholar]
- Krietenstein, N.; Abraham, S.; Venev, S.V.; Abdennur, N.; Gibcus, J.; Hsieh, T.S.; Parsi, K.M.; Yang, L.; Maehr, R.; Mirny, L.A. Ultrastructural Details of Mammalian Chromosome Architecture. Mol. Cell 2020, 78, 554–565.e7. [Google Scholar] [CrossRef]
- Roulland, S.; Lebailly, P.; Lecluse, Y.; Heutte, N.; Nadel, B.; Gauduchon, P. Long-Term Clonal Persistence and Evolution of T (14; 18)-Bearing B Cells in Healthy Individuals. Leukemia 2006, 20, 158–162. [Google Scholar] [CrossRef]
- Roulland, S.; Kelly, R.S.; Morgado, E.; Sungalee, S.; Solal-Celigny, P.; Colombat, P.; Jouve, N.; Palli, D.; Pala, V.; Tumino, R. T (14; 18) Translocation: A Predictive Blood Biomarker for Follicular Lymphoma. J. Clin. Oncol. 2014, 32, 1347. [Google Scholar] [CrossRef]
- Giannelli, F.; Moscarella, S.; Giannini, C.; Caini, P.; Monti, M.; Gragnani, L.; Romanelli, R.G.; Solazzo, V.; Laffi, G.; La Villa, G. Effect of Antiviral Treatment in Patients with Chronic HCV Infection and T (14; 18) Translocation. Blood 2003, 102, 1196–1201. [Google Scholar] [CrossRef]
- Santini, G.T.; Shah, P.P.; Karnay, A.; Jain, R. Aberrant Chromatin Organization at the Nexus of Laminopathy Disease Pathways. Nucleus 2022, 13, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Luperchio, T.R.; Wong, X.; Reddy, K.L. Genome Regulation at the Peripheral Zone: Lamina Associated Domains in Development and Disease. Curr. Opin. Genet. Dev. 2014, 25, 50–61. [Google Scholar] [CrossRef]
- Meaburn, K.J. Spatial Genome Organization and its Emerging Role as a Potential Diagnosis Tool. Front. Genet. 2016, 7, 134. [Google Scholar] [CrossRef]
- Tian, X.; Wang, Y.; Zhao, F.; Liu, J.; Yin, J.; Chen, D.; Ma, W.; Ke, X. A New Classification of Interphase Nuclei Based on Spatial Organizations of Chromosome 8 and 21 for T (8; 21)(q22; q22) Acute Myeloid Leukemia by Three-Dimensional Fluorescence in Situ Hybridization. Leuk. Res. 2015, 39, 1414–1420. [Google Scholar] [CrossRef]
- Lima, M.F.d.; Lisboa, M.d.O.; Terceiro, L.E.; Rangel-Pozzo, A.; Mai, S. Chromosome Territories in Hematological Malignancies. Cells 2022, 11, 1368. [Google Scholar] [CrossRef]
- Wiech, T.; Stein, S.; Lachenmaier, V.; Schmitt, E.; Schwarz-Finsterle, J.; Wiech, E.; Hildenbrand, G.; Werner, M.; Hausmann, M. Spatial Allelic Imbalance of BCL2 Genes and Chromosome 18 Territories in Nonneoplastic and Neoplastic Cervical Squamous Epithelium. Eur. Biophys. J. 2009, 38, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Tiukacheva, E.A.; Ulianov, S.V.; Karpukhina, A.; Razin, S.V.; Vassetzky, Y. 3D Genome Alterations and Editing in Pathology. Mol. Ther. 2023, 31, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N. Visualizing and Interpreting Cancer Genomics Data Via the Xena Platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Bikkul, M.U.; Faragher, R.G.; Worthington, G.; Meinke, P.; Kerr, A.R.; Sammy, A.; Riyahi, K.; Horton, D.; Schirmer, E.C.; Hubank, M. Telomere Elongation through hTERT Immortalization Leads to Chromosome Repositioning in Control Cells and Genomic Instability in Hutchinson-Gilford Progeria Syndrome Fibroblasts, Expressing a Novel SUN1 Isoform. Genes Chromosomes Cancer 2019, 58, 341–356. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence |
|---|---|
| β-actin Forward | 5′-AAGAGAGGCATCCTCACCCT-3′ |
| β-actin Reverse | 5′-TACATGGCTGGGGTGTTGAA-3′ |
| Bcl-2 Forward | 5′-ACGGAGGCTGGGATGCCTTTG-3′ |
| Bcl-2 Reverse | 5‘-ACACGAAGCGGTGCTTGGCA-3′ |
| Probe Name | Vector Type | Length, bp | Locus | Position | Labelling |
|---|---|---|---|---|---|
| RP5-998D24 | PAC | 165,821 | IGH | 14q32 | Digoxigenin-FITC |
| RP11-299P2 | BAC | 146,688 | BCL2 | 18q21 | Biotin-Texas Red |
| WCP14 * | Paint | Chr 14 | Biotin-Texas Red | ||
| WCP18 * | Paint | Chr 18 | FITC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garimberti, E.; Federico, C.; Ragusa, D.; Bruno, F.; Saccone, S.; Bridger, J.M.; Tosi, S. Alterations in Genome Organization in Lymphoma Cell Nuclei due to the Presence of the t(14;18) Translocation. Int. J. Mol. Sci. 2024, 25, 2377. https://doi.org/10.3390/ijms25042377
Garimberti E, Federico C, Ragusa D, Bruno F, Saccone S, Bridger JM, Tosi S. Alterations in Genome Organization in Lymphoma Cell Nuclei due to the Presence of the t(14;18) Translocation. International Journal of Molecular Sciences. 2024; 25(4):2377. https://doi.org/10.3390/ijms25042377
Chicago/Turabian StyleGarimberti, Elisa, Concetta Federico, Denise Ragusa, Francesca Bruno, Salvatore Saccone, Joanna Mary Bridger, and Sabrina Tosi. 2024. "Alterations in Genome Organization in Lymphoma Cell Nuclei due to the Presence of the t(14;18) Translocation" International Journal of Molecular Sciences 25, no. 4: 2377. https://doi.org/10.3390/ijms25042377
APA StyleGarimberti, E., Federico, C., Ragusa, D., Bruno, F., Saccone, S., Bridger, J. M., & Tosi, S. (2024). Alterations in Genome Organization in Lymphoma Cell Nuclei due to the Presence of the t(14;18) Translocation. International Journal of Molecular Sciences, 25(4), 2377. https://doi.org/10.3390/ijms25042377