
Potential Candidate Molecule of Photosystem II Inhibitor Herbicide—Brassicanate A Sulfoxide


Abstract
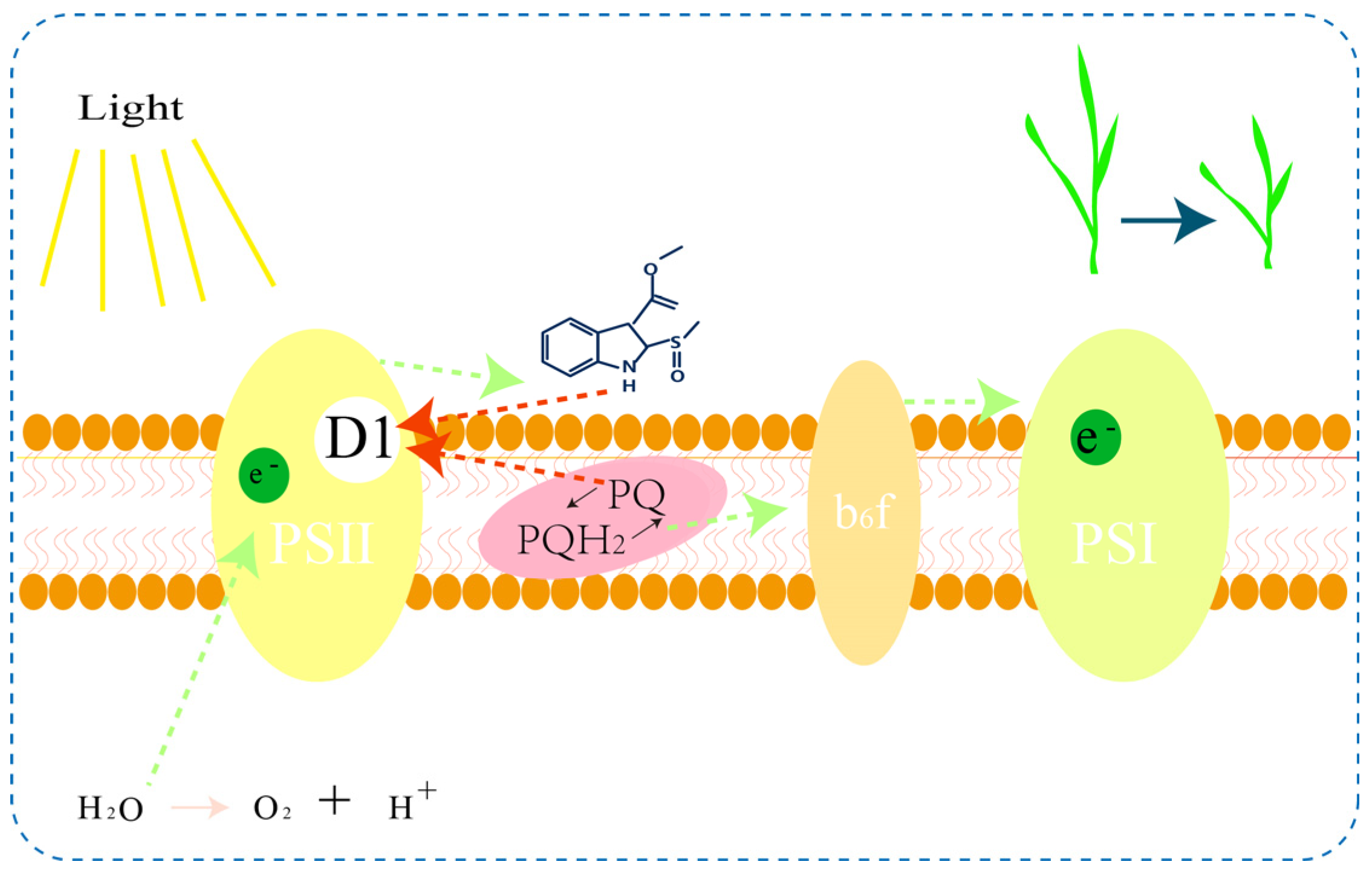
:1. Introduction
2. Results
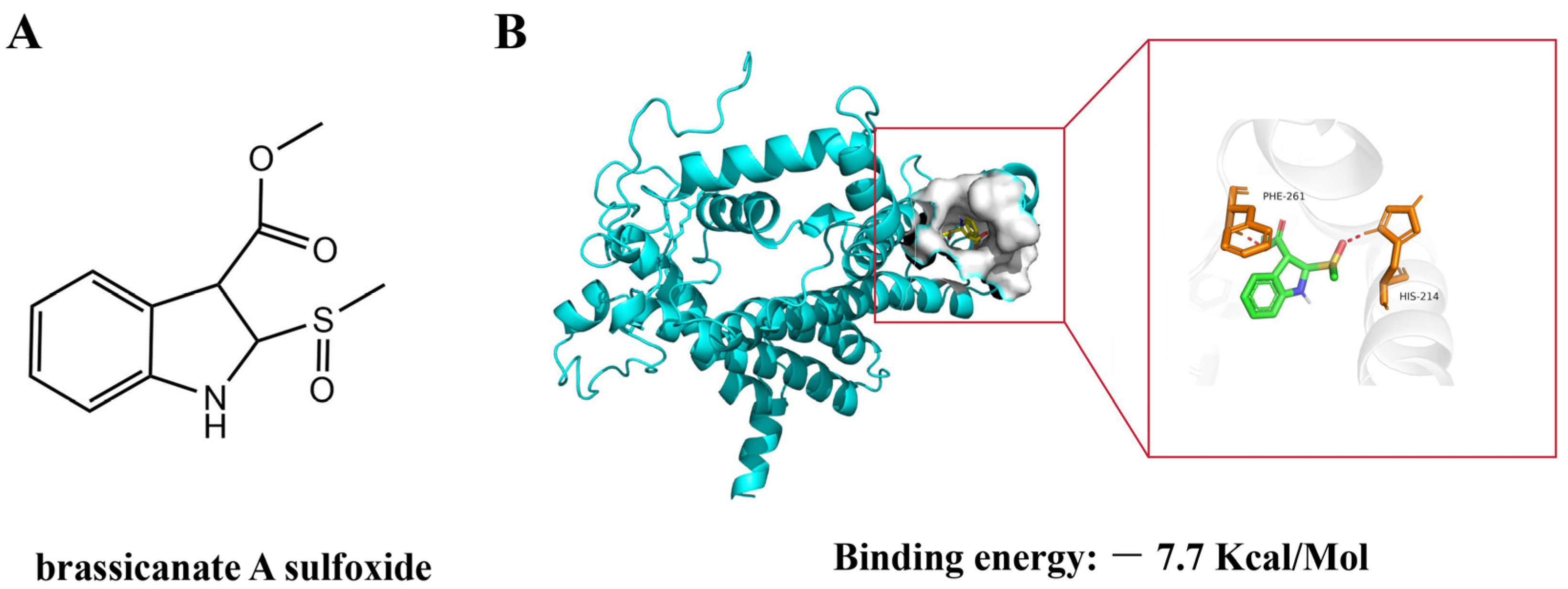
2.1. The Docking of Brassicanate A Sulfoxide with PSBD1
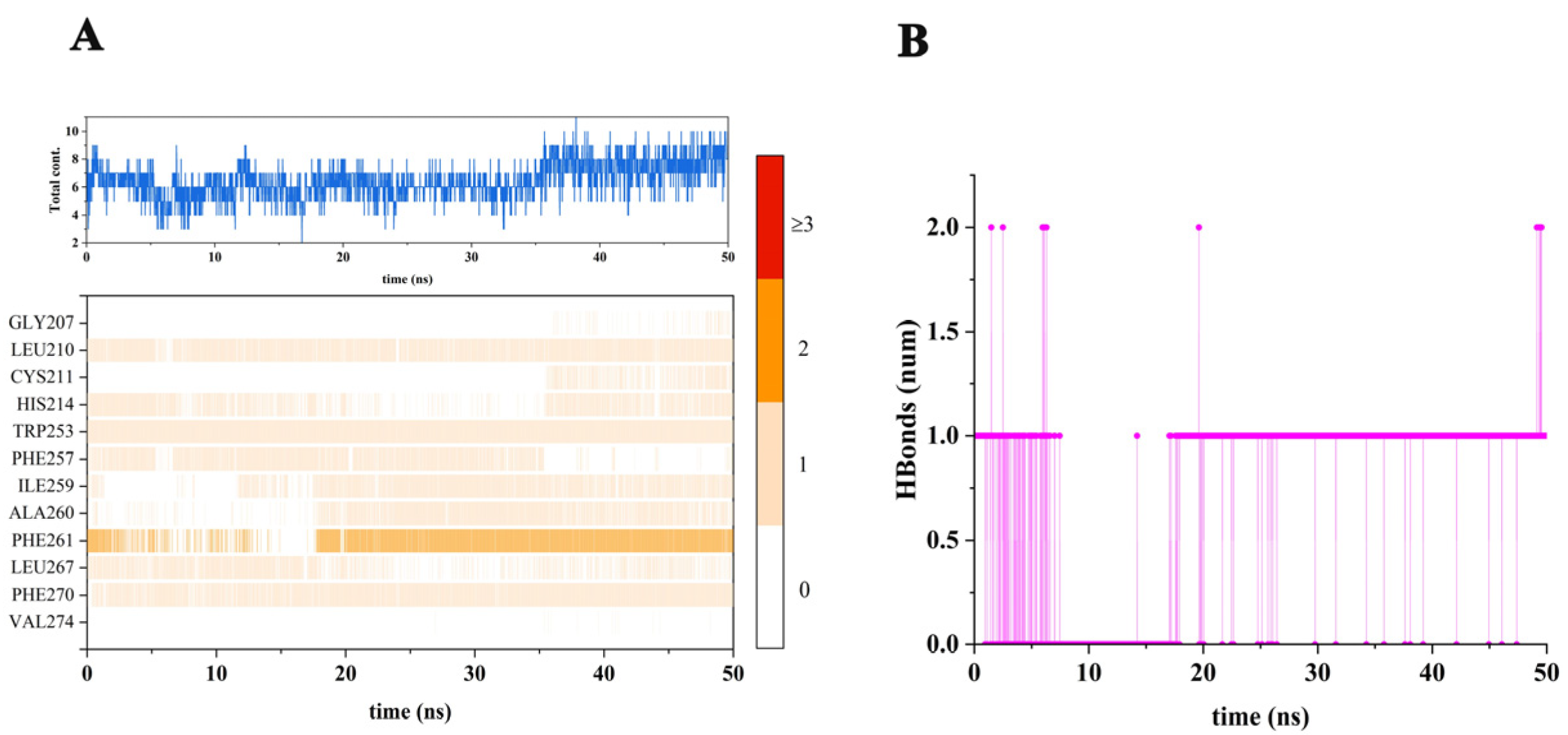
2.2. Stability of the Simulated System
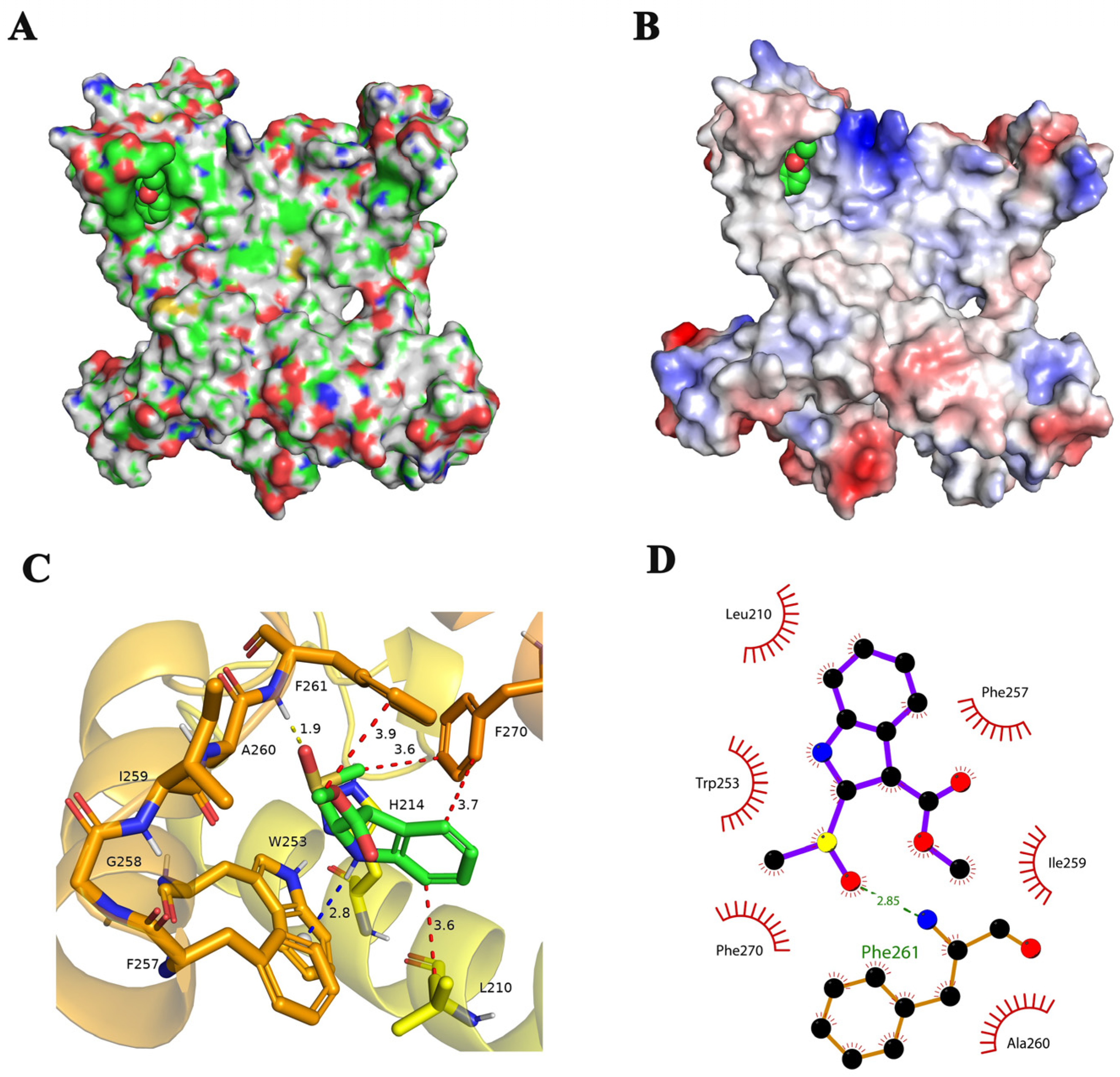
2.3. Binding Mode of Brassicanate A Sulfoxide with PSBD1
2.4. Types and Quantities of Amino Acids Involved in the Interaction between Brassicanate A Sulfoxide and PSBD1
2.5. Binding Energy of Brassicanate A Sulfoxide with PSBD1
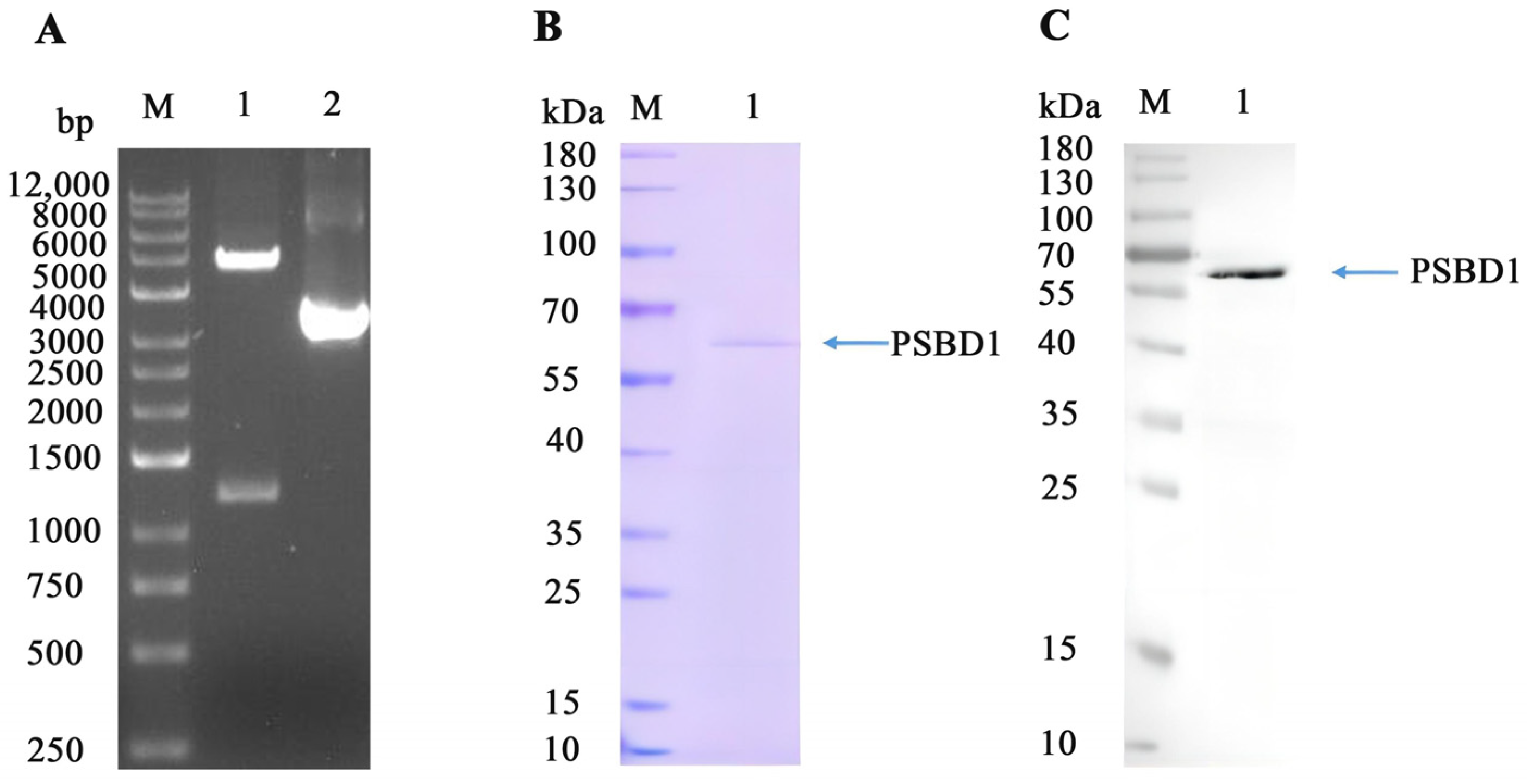
2.6. Expression and Purification of PSBD1
2.7. Validation of the Binding between PSBD1 and Brassicanate A Sulfoxide
3. Discussion
4. Materials and Methods
4.1. Molecular Docking
4.2. Preparation of PSBD1 and Brassicanate A Sulfoxide Structure
4.3. Molecular Dynamics Simulations
4.4. Binding Free Energy Calculation
4.5. The Expression of PSBD1
4.6. Purification of PSBD1
4.7. Biolayer Interferometry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nasim, G.; Shabbir, A. Invasive Weed Species—A Threat to Sustainable Agriculture. In Crop Production for Agricultural Improvement; Ashraf, M., Öztürk, M., Ahmad, M.S.A., Aksoy, A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 523–556. [Google Scholar]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical pesticides and human health: The urgent need for a new concept in agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef]
- Yadav, I.C.; Devi, N.L. Pesticides classification and its impact on human and environment. Environ. Sci. Eng. 2017, 6, 140–158. [Google Scholar]
- Singh, H.; Batish, D.R.; Kohli, R. Allelopathic interactions and allelochemicals: New possibilities for sustainable weed management. Crit. Rev. Plant Sci. 2003, 22, 239–311. [Google Scholar] [CrossRef]
- Islam, A.M.; Yeasmin, S.; Qasem, J.R.S.; Juraimi, A.S.; Anwar, M.P. Allelopathy of medicinal plants: Current status and future prospects in weed management. Agric. Sci. 2018, 9, 1569–1588. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Dong, B.; Wang, D.; Nian, Y.; Zhou, H. Isolation and Identification of Herbicidal Active Compounds from Brassica oleracea L. and Exploration of the Binding Sites of Brassicanate A Sulfoxide. Plants 2023, 12, 2576. [Google Scholar] [CrossRef] [PubMed]
- Reifler, M.J.; Szalai, V.A.; Peterson, C.N.; Brudvig, G.W. Effects of tail-like substituents on the binding of competitive inhibitors to the QB site of photosystem II. J. Mol. Recognit. 2001, 14, 157–165. [Google Scholar] [CrossRef]
- Yaduraju, N.; Sharma, A.; Rao, A. Weeds in Indian Agriculture: Problems and prospects to become self sufficient. Indian Farming 2015, 65, 2–6. [Google Scholar]
- Lambrev, P.; Goltsev, V. pH dependence of the effects of diuron, atrazine and dinoseb on the luminescent properties of thylakoid membranes. Bulg. J. Plant Physiol. 2001, 27, 85–100. [Google Scholar]
- Breton, F.; Piletska, E.V.; Karim, K.; Rouillon, R.; Piletsky, S.A. Mimicking the Plastoquinone-Binding Pocket of Photosystem II Using Molecularly Imprinted Polymers. In Biotechnological Applications of Photosynthetic Proteins: Biochips, Biosensors and Biodevices; Giardi, M.T., Piletska, E.V., Eds.; Springer: Boston, MA, USA, 2006; pp. 155–165. [Google Scholar]
- Sobolev, V.; Edelman, M. Modeling the quinone-B binding site of the photosystem-II reaction center using notions of complementarity and contact-surface between atoms. Proteins Struct. Funct. Bioinform. 1995, 21, 214–225. [Google Scholar] [CrossRef]
- Petersen, R.L. Strategies using bio-layer interferometry biosensor technology for vaccine research and development. Biosensors 2017, 7, 49. [Google Scholar] [CrossRef]
- Auer, S.; Azizi, L.; Faschinger, F.; Blazevic, V.; Vesikari, T.; Gruber, H.J.; Hytönen, V.P. Stable immobilisation of His-tagged proteins on BLI biosensor surface using cobalt. Sens. Actuators B Chem. 2017, 243, 104–113. [Google Scholar] [CrossRef]
- Chiodi, E.; Marn, A.M.; Bakhshpour, M.; Lortlar Ünlü, N.; Ünlü, M.S. The effects of three-dimensional ligand immobilization on kinetic measurements in biosensors. Polymers 2022, 14, 241. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Abdoli, A. Biotransformation of rutabaga phytoalexins by the fungus Alternaria brassicicola: Unveiling the first hybrid metabolite derived from a phytoalexin and a fungal polyketide. Bioorganic Med. Chem. 2017, 25, 557–567. [Google Scholar] [CrossRef]
- de Souza, J.M.; Fazolo, B.R.; Lacerda, J.W.F.; Moura, M.S.; Dos Santos, A.C.R.; de Vasconcelos, L.G.; de Sousa Junior, P.T.; Dall’Oglio, E.L.; Ali, A.; Sampaio, O.M.; et al. Rational Design, Synthesis and Evaluation of Indole Nitrogen Hybrids as Photosystem II Inhibitors. Photochem. Photobiol. 2020, 96, 1233–1242. [Google Scholar] [CrossRef]
- Devendar, P.; Yang, G.F. Sulfur-Containing Agrochemicals. Top. Curr. Chem. 2017, 375, 82. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hwang, I.; Ryu, S.; Han, K.; Kwon, Y. Aryl sulfoxide scaffold useful as herbicide. Appl. Biol. Chem. 2023, 66, 67. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, B.; Wang, D.; Jia, X.; Zhang, Q.; Liu, W.; Zhou, H. Investigation into the Binding Site of (-)-Spirobrassinin for Herbicidal Activity Using Molecular Docking and Molecular Dynamics Simulations. Appl. Sci. 2023, 13, 7287. [Google Scholar] [CrossRef]
- Tice, C.M. Selecting the right compounds for screening: Does Lipinski’s Rule of 5 for pharmaceuticals apply to agrochemicals? Pest Manag. Sci. 2001, 57, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Clarke, E.; Delaney, J. Physical and Molecular Properties of Agrochemicals: An Analysis of Screen Inputs, Hits, Leads, and Products. CHIMIA Int. J. Chem. 2003, 57, 731–734. [Google Scholar] [CrossRef]
- Szigeti, Z.; Tóth, E.; Paless, G. Mode of action of photosynthesis inhibiting 4-hydroxy-benzonitriles containing nitro group. Photosynth. Res. 1982, 3, 347–356. [Google Scholar] [CrossRef]
- Nain-Perez, A.; Barbosa, L.C.; Maltha, C.R.; Giberti, S.; Forlani, G. Tailoring natural abenquines to inhibit the photosynthetic electron transport through interaction with the D1 protein in photosystem II. J. Agric. Food Chem. 2017, 65, 11304–11311. [Google Scholar] [CrossRef]
- Mengistu, L.W.; Christoffers, M.J.; Lym, R.G. A psbA mutation in Kochia scoparia (L) Schrad from railroad rights-of-way with resistance to diuron, tebuthiuron and metribuzin. Pest Manag. Sci. Former. Pestic. Sci. 2005, 61, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.C.; Donnelly-Vanderloo, M.J.; Hume, D.J. Triazine-resistant crops: The agronomic impact and physiological consequences of chloroplast mutation. In Herbicide-Resistant Crops; CRC Press: Boca Raton, FL, USA, 2018; pp. 107–126. [Google Scholar]
- King, L. The Baculovirus Expression System: A Laboratory Guide; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Wiseman, D.N.; Otchere, A.; Patel, J.H.; Uddin, R.; Pollock, N.L.; Routledge, S.J.; Rothnie, A.J.; Slack, C.; Poyner, D.R.; Bill, R.M. Expression and purification of recombinant G protein-coupled receptors: A review. Protein Expr. Purif. 2020, 167, 105524. [Google Scholar] [CrossRef] [PubMed]
- Douris, V.; Swevers, L.; Labropoulou, V.; Andronopoulou, E.; Georgoussi, Z.; Iatrou, K. Stably transformed insect cell lines: Tools for expression of secreted and membrane-anchored proteins and high-throughput screening platforms for drug and insecticide discovery. Adv. Virus Res. 2006, 68, 113–156. [Google Scholar] [PubMed]
- Harani, A.; VijayaRatnam, J.; Dipankar, B.; Kumar, D.S.; Lalitha, M.B.; Kumar, S. Molecular interaction studies of phosphatidylcholine as drug delivery substrate for asenapine maleate. Curr. Sci. 2018, 115, 499–504. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Anandakrishnan, R.; Aguilar, B.; Onufriev, A.V. H++ 3.0: Automating p K prediction and the preparation of biomolecular structures for atomistic molecular modeling and simulations. Nucleic Acids Res. 2012, 40, W537–W541. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; MacKerell, A.D., Jr. Computational ligand-based rational design: Role of conformational sampling and force fields in model development. MedChemComm 2011, 2, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Jakalian, A.; Jack, D.B.; Bayly, C.I. Fast, efficient generation of high-quality atomic charges. AM1-BCC model: II. Parameterization and validation. J. Comput. Chem. 2002, 23, 1623–1641. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, C.; Wang, X.; Zhu, T. A test of AMBER force fields in predicting the secondary structure of α-helical and β-hairpin peptides. Chem. Phys. Lett. 2017, 679, 112–118. [Google Scholar] [CrossRef]
- Case, D.A.; Cheatham, T.E., III; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, K.; Jaeger, V.W.; Pfaendtner, J. The general AMBER force field (GAFF) can accurately predict thermodynamic and transport properties of many ionic liquids. J. Phys. Chem. B 2015, 119, 5882–5895. [Google Scholar] [CrossRef] [PubMed]
- Petersen, H.G. Accuracy and efficiency of the particle mesh Ewald method. J. Chem. Phys. 1995, 103, 3668–3679. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef]
- Miller, B.R., III; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]
- Sung, L.-Y.; Chen, C.-L.; Lin, S.-Y.; Li, K.-C.; Yeh, C.-L.; Chen, G.-Y.; Lin, C.-Y.; Hu, Y.-C. Efficient gene delivery into cell lines and stem cells using baculovirus. Nat. Protoc. 2014, 9, 1882–1899. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Energy | Value |
|---|---|
| ΔE vdw | −8.86 |
| ΔE ele | −4.17 |
| ΔE GB | 6.63 |
| ΔE surf | −4.02 |
| ΔE gas | −13.03 |
| ΔG solv | 2.61 |
| ΔG bind | −10.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, D.; Dong, B.; Hao, J.; Jia, X.; Zhou, H. Potential Candidate Molecule of Photosystem II Inhibitor Herbicide—Brassicanate A Sulfoxide. Int. J. Mol. Sci. 2024, 25, 2400. https://doi.org/10.3390/ijms25042400
Wang Y, Wang D, Dong B, Hao J, Jia X, Zhou H. Potential Candidate Molecule of Photosystem II Inhibitor Herbicide—Brassicanate A Sulfoxide. International Journal of Molecular Sciences. 2024; 25(4):2400. https://doi.org/10.3390/ijms25042400
Chicago/Turabian StyleWang, Yu, Dong Wang, Baozhu Dong, Jianxiu Hao, Xinyu Jia, and Hongyou Zhou. 2024. "Potential Candidate Molecule of Photosystem II Inhibitor Herbicide—Brassicanate A Sulfoxide" International Journal of Molecular Sciences 25, no. 4: 2400. https://doi.org/10.3390/ijms25042400