
Antioxidant, Anti-Diabetic, and Anti-Inflammation Activity of Garcinia livingstonei Aqueous Leaf Extract: A Preliminary Study
 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Cytotoxicity Effect of Aqueous G. livingstonei Extract
2.2. Quantification of Specific Phenolic Compounds
2.3. Total Antioxidant Capacity
2.4. UHPLC-MS Identification of Phenolic Compounds
2.4.1. Identification of Flavan-3-Ols, Flavonols, and Flavone
2.4.2. Identification of Benzophenones
2.4.3. Identification of Hydroxycinnamic Acids
2.5. Anti-Inflammatory Activity
2.6. Glucose Uptake and Utilisation
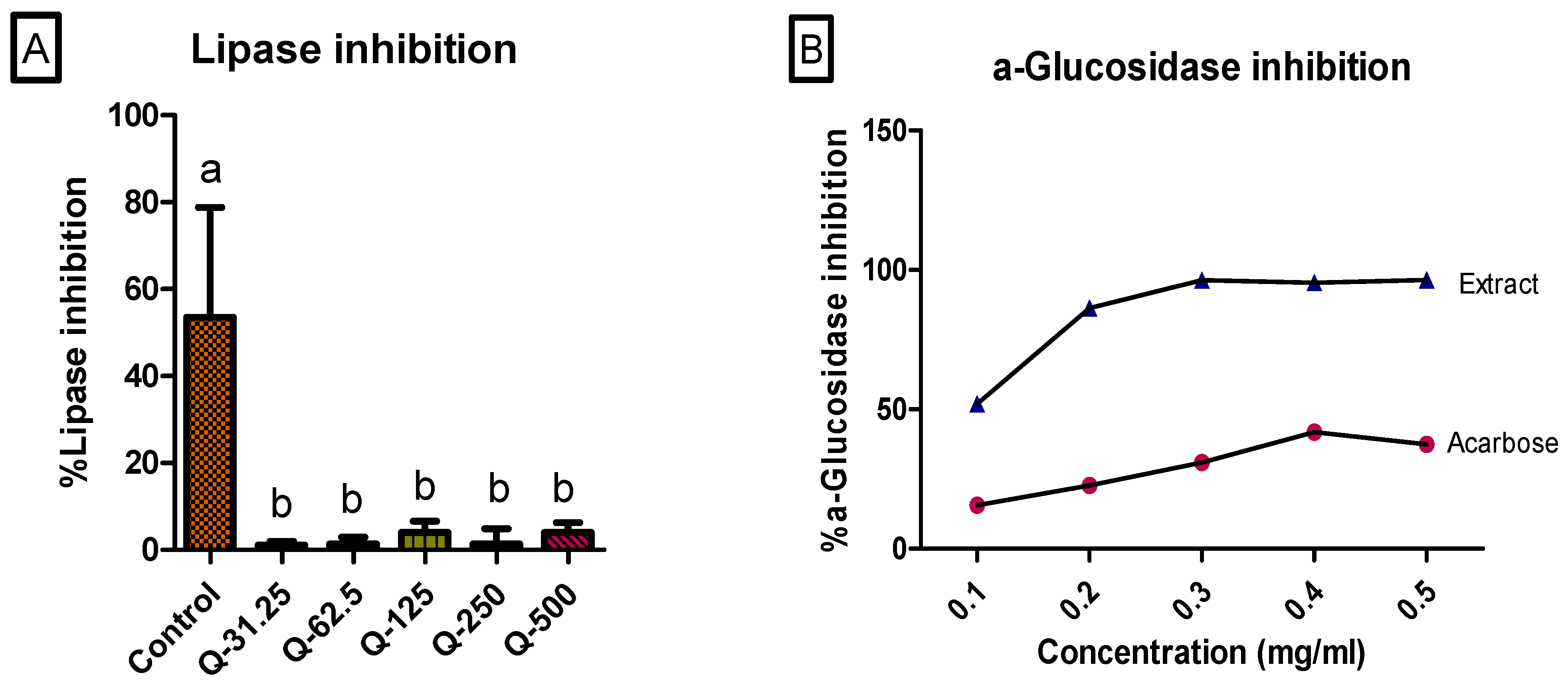
2.7. α-Glucosidase and Lipase Inhibition
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Plant Extraction
3.4. Phytochemical Analysis
3.4.1. Determination of Total Polyphenol
3.4.2. Determination of Flavanol Content
3.4.3. Determination of Total Alkaloid
3.4.4. Determination of Flavanol Content
3.5. Ultra-High Performance Liquid Chromatography Mass Spectrometry (UHPLC-MS) Analysis
3.6. Antioxidant Capacity
3.6.1. Ferric Reducing Antioxidant Power Assay
3.6.2. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
3.6.3. 2,2-diphenyl-1-picrylhydrazyl (DPPH) Assay
3.7. Anti-Inflammatory Effect Determination
3.8. Glucose Uptake and Utilisation
3.8.1. Cell Line Maintenance
3.8.2. Glucose Uptake
3.8.3. Glucose Utilisation
3.9. Cytotoxicity Determination-MTT Assay
3.10. Lipase Inhibition
3.11. α-Glucosidase Inhibition
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pant, D.; Aryal, B.; Pun, D.; Sharma, S.; Joshi, G.P. Inhibition of a-amylase and a-glucosidase activities in vitro by extracts of selected medicinal plants. Biodiversitas J. Biol. Divers. 2021, 22, 314. [Google Scholar] [CrossRef]
- Etsassala, N.G.E.R.; Badmus, J.A.; Marnewick, J.L.; Iwuoha, E.I.; Nchu, F.; Hussein, A.A. Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities, Molecular Docking, and Antioxidant Capacities of Salvia aurita Constituents. Antioxidants 2020, 9, 1149. [Google Scholar] [CrossRef]
- Jiang, S.; Young, J.L.; Wang, K.; Qian, Y.; Cai, L. Diabetic-induced alterations in hepatic glucose and lipid metabolism: The role of type 1 and type 2 diabetes mellitus (Review). Mol. Med. Rep. 2020, 22, 603–611. [Google Scholar] [CrossRef]
- Papachristoforou, E.; Lambadiari, V.; Maratou, E.; Makrilakis, K. Association of Glycemic Indices (Hyperglycemia, Glucose Variability, and Hypoglycemia) with Oxidative Stress and Diabetic Complications. J. Diabetes Res. 2020, 2020, 7489795. [Google Scholar] [CrossRef]
- Alsenosy, A.-W.A.; El-Far, A.H.; Sadek, K.M.; Ibrahim, S.A.; Atta, M.S.; Sayed-Ahmed, A.; Al Jaouni, S.K.; Mousa, S.A. Graviola (Annona muricata) attenuates behavioural alterations and testicular oxidative stress induced by streptozotocin in diabetic rats. PLoS ONE 2019, 14, e0222410. [Google Scholar] [CrossRef]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced Glycation End Products and Diabetes Mellitus: Mechanisms and Perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef]
- Lima, J.E.; Moreira, N.C.; Sakamoto-Hojo, E.T. Mechanisms underlying the pathophysiology of type 2 diabetes: From risk factors to oxidative stress, metabolic dysfunction, and hyperglycemia. Mutat. Res. Toxicol. Environ. Mutagen. 2021, 874–875, 503437. [Google Scholar] [CrossRef]
- Teh, H.X.; Phang, S.J.; Looi, M.L.; Kuppusamy, U.R.; Arumugam, B. Molecular pathways of NF-ĸB and NLRP3 inflammasome as potential targets in the treatment of inflammation in diabetic wounds: A review. Life Sci. 2023, 334, 122228. [Google Scholar] [CrossRef]
- Rayego-Mateos, S.; Morgado-Pascual, J.L.; Opazo-Ríos, L.; Guerrero-Hue, M.; García-Caballero, C.; Vázquez-Carballo, C.; Mas, S.; Sanz, A.B.; Herencia, C.; Mezzano, S.; et al. Pathogenic Pathways and Therapeutic Approaches Targeting Inflammation in Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 3798. [Google Scholar] [CrossRef]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2022, 21, 1049–1079. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P.; Slama, P.; Roychoudhury, S. Oxidative Stress, Testicular Inflammatory Pathways, and Male Reproduction. Int. J. Mol. Sci. 2021, 22, 10043. [Google Scholar] [CrossRef]
- Maresch, C.C.; Stute, D.C.; Fleming, T.; Lin, J.; Hammes, H.-P.; Linn, T. Hyperglycemia induces spermatogenic disruption via major pathways of diabetes pathogenesis. Sci. Rep. 2019, 9, 13074. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Erzse, A.; Stacey, N.; Chola, L.; Tugendhaft, A.; Freeman, M.; Hofman, K. The direct medical cost of type 2 diabetes mellitus in South Africa: A cost of illness study. Glob. Health Action 2019, 12, 1636611. [Google Scholar] [CrossRef]
- Grundlingh, N.; Zewotir, T.T.; Roberts, D.J.; Manda, S. Assessment of prevalence and risk factors of diabetes and pre-diabetes in South Africa. J. Health Popul. Nutr. 2022, 41, 7. [Google Scholar] [CrossRef]
- Wang, X.; Li, Q.; Han, X.; Gong, M.; Yu, Z.; Xu, B. Electroacupuncture Alleviates Diabetic Peripheral Neuropathy by Regulating Glycolipid-Related GLO/AGEs/RAGE Axis. Front. Endocrinol. 2021, 12, 655591. [Google Scholar] [CrossRef]
- Liu, T.-T.; Liu, X.-T.; Chen, Q.-X.; Shi, Y. Lipase Inhibitors for Obesity: A Review. Biomed. Pharmacother. 2020, 128, 110314. [Google Scholar] [CrossRef]
- Khan, M.; Ahad, G.; Alam, A.; Ullah, S.; Khan, A.; Kanwal; Salar, U.; Wadood, A.; Ajmal, A.; Khan, K.M.; et al. Synthesis of new bis(dimethylamino)benzophenone hydrazone for diabetic management: In-vitro and in-silico approach. Heliyon 2024, 10, e23323. [Google Scholar] [CrossRef]
- Gutierrez, R.M.P.; Velazquez, E.G. Glucopyranoside flavonoids isolated from leaves of Spinacia oleracea (spinach) inhibit the formation of advanced glycation end products (AGEs) and aldose reductase activity (RLAR). Biomed. Pharmacother. 2020, 128, 110299. [Google Scholar] [CrossRef]
- Mudau, T.; Olowoyo, J.; Amoo, S. Ethnobotanical assessment of medicinal plants used traditionally for treating diabetes in Vhembe district, Limpopo Province, South Africa. S. Afr. J. Bot. 2022, 146, 304–324. [Google Scholar] [CrossRef]
- Abdul-Rahman, A.M.; Elwekeel, A.; Alruhaimi, R.S.; Kamel, E.M.; Bin-Ammar, A.; Mahmoud, A.M.; Moawad, A.S.; Zaki, M.A. Multi-target action of Garcinia livingstonei extract and secondary metabolites against fatty acid synthase, α-glucosidase, and xanthine oxidase. Saudi Pharm. J. 2023, 31, 101762. [Google Scholar] [CrossRef]
- Ramadwa, T.; Meddows-Taylor, S. Garcinia livingstonei T. Anderson (Clusiaceae): A comprehensive review of traditional uses, phytochemistry, pharmacology and toxicity. S. Afr. J. Bot. 2023, 162, 767–782. [Google Scholar] [CrossRef]
- Joseph, K.S.; Bolla, S.; Joshi, K.; Bhat, M.; Naik, K.; Patil, S.; Bendre, S.; Gangappa, B.; Haibatti, V.; Payamalle, S.; et al. Determination of Chemical Composition and Nutritive Value with Fatty Acid Compositions of African Mangosteen (Garcinia Livingstonei). Erwerbs-Obstbau 2016, 59, 195–202. [Google Scholar] [CrossRef]
- Yang, H.; Figueroa, M.; To, S.; Baggett, S.; Jiang, B.; Basile, M.J.; Weinstein, I.B.; Kennelly, E.J. Benzophenones and Biflavonoids from Garcinia livingstonei Fruits. J. Agric. Food Chem. 2010, 58, 4749–4755. [Google Scholar] [CrossRef]
- Martín, M.; Ramos, S. Impact of Dietary Flavanols on Microbiota, Immunity and Inflammation in Metabolic Diseases. Nutrients 2021, 13, 850. [Google Scholar] [CrossRef]
- Okaiyeto, K.; Kerebba, N.; Rautenbach, F.; Singh, S.K.; Dua, K.; Oguntibeju, O.O. UPLC-ESI-QTOF-MS phenolic compounds identification and quantification from ethanolic extract of Myrtus communis ‘Variegatha’: In vitro antioxidant and antidiabetic potentials. Arab. J. Chem. 2023, 16, 104447. [Google Scholar] [CrossRef]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S.; et al. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complement. Med. 2017, 8, 361–376. [Google Scholar] [CrossRef]
- Nna, V.U.; Abu Bakar, A.B.; Ahmad, A.; Eleazu, C.O.; Mohamed, M. Oxidative Stress, NF-κB-Mediated Inflammation and Apoptosis in the Testes of Streptozotocin–Induced Diabetic Rats: Combined Protective Effects of Malaysian Propolis and Metformin. Antioxidants 2019, 8, 465. [Google Scholar] [CrossRef]
- Ambika, S.; Saravanan, R.; Thirumavalavan, K. Antidiabetic and antihyperlipidemic effect of p-hydroxycinnamic acid on streptozotocin-induced diabetic Wistar rats. Biomed. Aging Pathol. 2013, 3, 253–257. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular Mechanisms Linking Oxidative Stress and Diabetes Mellitus. Oxidative Med. Cell. Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef]
- Budniak, L.; Vasenda, M.; Slobodianiuk, L. Determination of flavonoids and hydroxycinnamic acids in tablets with thick extract of primula denticulata Smith. Pharmacologyonline 2021, 2, 1244–1253. [Google Scholar]
- Mulholland, D.A.; Mwangi, E.M.; Dlova, N.C.; Plant, N.; Crouch, N.R.; Coombes, P.H. Non-toxic melanin production inhibitors from Garcinia livingstonei (Clusiaceae). J. Ethnopharmacol. 2013, 149, 570–575. [Google Scholar] [CrossRef]
- Anitha, S.; Krishnan, S.; Senthilkumar, K.; Sasirekha, V. Theoretical investigation on the structure and antioxidant activity of (+) catechin and (−)epicatechin—A comparative study. Mol. Phys. 2020, 118, e1745917. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, S.; Kumar, B. LC-MS Identification of Proanthocyanidins in Bark and Fruit of six Terminalia species. Nat. Prod. Commun. 2018, 13, 1934578X1801300511. [Google Scholar] [CrossRef]
- Manavi, S.P.; Amiri, T.; Mozafaryan, M.J. Role of flavonoids in diabetes. J. Rev. Med. Sci. 2021, 1, 149–161. [Google Scholar] [CrossRef]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Hamid, Z.A.; Budin, S.B. The Potential Role of Flavonoids in Ameliorating Diabetic Cardiomyopathy via Alleviation of Cardiac Oxidative Stress, Inflammation and Apoptosis. Int. J. Mol. Sci. 2021, 22, 5094. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, Z.; Xiang, S.-Z.; Jin, Y.-G.; Wei, W.-Y.; Bian, Z.-Y.; Deng, W.; Tang, Q.-Z. Nobiletin attenuates cardiac dysfunction, oxidative stress, and inflammatory in streptozotocin: Induced diabetic cardiomyopathy. Mol. Cell. Biochem. 2016, 417, 87–96. [Google Scholar] [CrossRef]
- Muriithi, E.; Bojase-Moleta, G.; Majinda, R.R. Benzophenone derivatives from Garcinia livingstonei and their antioxidant activities. Phytochem. Lett. 2016, 18, 29–34. [Google Scholar] [CrossRef]
- Moon, K.M.; Yang, J.-H.; Lee, M.-K.; Kwon, E.-B.; Baek, J.; Hwang, T.; Kim, J.-I.; Lee, B. Maclurin Exhibits Antioxidant and Anti-Tyrosinase Activities, Suppressing Melanogenesis. Antioxidants 2022, 11, 1164. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Q.; Liu, M.-Y.; Ruan, J.-Y.; Yu, H.-Y.; Li, J.; Wang, T. Effects of Benzophenones from Mango Leaves on Lipid Metabolism. Chem. Pharm. Bull. 2019, 67, 634–639. [Google Scholar] [CrossRef]
- Surana, K.; Chaudhary, B.; Diwaker, M.; Sharma, S. Benzophenone: A ubiquitous scaffold in medicinal chemistry. MedChemComm 2018, 9, 1803–1817. [Google Scholar] [CrossRef]
- Ma, C.; Dastmalchi, K.; Whitaker, B.D.; Kennelly, E.J. Two New Antioxidant Malonated Caffeoylquinic Acid Isomers in Fruits of Wild Eggplant Relatives. J. Agric. Food Chem. 2011, 59, 9645–9651. [Google Scholar] [CrossRef]
- Kiliç, I.; Yeşiloğlu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Liu, W.; Li, J.; Zhang, X.; Zu, Y.; Yang, Y.; Liu, W.; Xu, Z.; Gao, H.; Sun, X.; Jiang, X.; et al. Current Advances in Naturally Occurring Caffeoylquinic Acids: Structure, Bioactivity, and Synthesis. J. Agric. Food Chem. 2020, 68, 10489–10516. [Google Scholar] [CrossRef]
- Theodosis-Nobelos, P.; Papagiouvannis, G.; Rekka, E.A. Ferulic, Sinapic, 3,4-Dimethoxycinnamic Acid and Indomethacin Derivatives with Antioxidant, Anti-Inflammatory and Hypolipidemic Functionality. Antioxidants 2023, 12, 1436. [Google Scholar] [CrossRef]
- Alam, A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13, 27. [Google Scholar] [CrossRef]
- Andrade, P.B.; Leitão, R.; Seabra, R.M.; Oliveira, M.B.; Ferreira, M.A. 3,4-Dimethoxycinnamic acid levels as a tool for differentiation of Coffea canephora var. robusta and Coffea arabica. Food Chem. 1998, 61, 511. [Google Scholar] [CrossRef]
- Nozawa, Y.; Sakai, N.; Arai, K.; Kawasaki, Y.; Harada, K.-I. Reliable and sensitive analysis of amino acids in the peptidoglycan of actinomycetes using the advanced Marfey’s method. J. Microbiol. Methods 2007, 70, 306–311. [Google Scholar] [CrossRef]
- Mighri, H.; Akrout, A.; Bennour, N.; Eljeni, H.; Zammouri, T.; Neffati, M. LC/MS method development for the determination of the phenolic compounds of Tunisian Ephedra alata hydro-methanolic extract and its fractions and evaluation of their antioxidant activities. S. Afr. J. Bot. 2019, 124, 102–110. [Google Scholar] [CrossRef]
- Taamalli, A.; Arráez-Román, D.; Abaza, L.; Iswaldi, I.; Fernández-Gutiérrez, A.; Zarrouk, M.; Segura-Carretero, A. LC-MS-based metabolite profiling of methanolic extracts from the medicinal and aromatic species Mentha pulegium and Origanum majorana. Phytochem. Anal. 2015, 26, 320–330. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhong, B.; Yang, Z.; Zhao, W.; Shi, L.; Aziz, A.; Rauf, A.; Aljohani, A.S.; Alhumaydhi, F.A.; Suleria, H.A.R. LC-ESI-QTOF-MS/MS Characterization and Estimation of the Antioxidant Potential of Phenolic Compounds from Different Parts of the Lotus (Nelumbo nucifera) Seed and Rhizome. ACS Omega 2022, 7, 14630–14642. [Google Scholar] [CrossRef]
- The Metabolomics Innovation Centre. The Metabolomics Innovation Centre: FooDB (Version 1). Available online: https://foodb.ca/ (accessed on 19 May 2020).
- Mugaranja, K.P.; Kulal, A. Alpha glucosidase inhibition activity of phenolic fraction from Simarouba glauca: An in-vitro, in-silico and kinetic study. Heliyon 2020, 6, e04392. [Google Scholar] [CrossRef]
- Falleh, H.; Oueslati, S.; Guyot, S.; Ben Dali, A.; Magné, C.; Abdelly, C.; Ksouri, R. LC/ESI-MS/MS characterisation of procyanidins and propelargonidins responsible for the strong antioxidant activity of the edible halophyte Mesembryanthemum edule L. Food Chem. 2011, 127, 1732–1738. [Google Scholar] [CrossRef]
- Burlec, A.F.; Pecio, L.; Mircea, C.; Cioancă, O.; Corciovă, A.; Nicolescu, A.; Oleszek, W.; Hăncianu, M. Chemical Profile and Antioxidant Activity of Zinnia elegans Jacq. Fractions. Molecules 2019, 24, 2934. [Google Scholar] [CrossRef]
- Kiani, H.S.; Ahmad, W.; Nawaz, S.; Farah, M.A.; Ali, A. Optimized Extraction of Polyphenols from Unconventional Edible Plants: LC-MS/MS Profiling of Polyphenols, Biological Functions, Molecular Docking, and Pharmacokinetics Study. Molecules 2023, 28, 6703. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Shamilov, A.A. Catechin-O-Rhamnosides from Vaccinium vitis-idaea Stems. Chem. Nat. Compd. 2022, 58, 269–273. [Google Scholar] [CrossRef]
- Royal Society of Chemistry ChemSpider|Search and Share Chemistry. Available online: http://www.chemspider.com/ (accessed on 23 November 2018).
- Kumari, A.; Bhagwat, P.U.; Chhabria, D.; Kirubakaran, S.; Maurya, S.K. Comparative Study of Secondary Metabolites of Camellia sinensis and Camellia japonica Flowers Growing in the Kangra Region of India and Their Cytotoxicity Evaluation. ACS Food Sci. Technol. 2023, 3, 1484–1495. [Google Scholar] [CrossRef]
- Rai, A.; Kumari, K.; Han, S.S. Polyphenolic profiling of Victoria Amazonica using MRM LC-MS/MS: A comparative analysis of various plant parts. Sci. Hortic. 2023, 320, 112206. [Google Scholar] [CrossRef]
- Chu, M.-J.; Liu, X.-M.; Yan, N.; Wang, F.-Z.; Du, Y.-M.; Zhang, Z.-F. Partial Purification, Identification, and Quantitation of Antioxidants from Wild Rice (Zizania latifolia). Molecules 2018, 23, 2782. [Google Scholar] [CrossRef]
- Matei, A.O.; Gatea, F.; Radu, G.L. Analysis of Phenolic Compounds in Some Medicinal Herbs by LC–MS. J. Chromatogr. Sci. 2015, 53, 1147–1154. [Google Scholar] [CrossRef]
- Coppock, R.W.; Dziwenka, M. Green tea. In Nutraceuticals. Efficacy, Safety and Toxicity; Academic Press: Cambridge, MA, USA, 2021; pp. 697–723. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.; Sui, Y.; Mei, X.; Shi, J.; Cai, S.; Xiong, T.; Carrillo, C.; Castagnini, J.M.; Zhu, Z.; et al. Comparing the LC-MS Phenolic Acids Profiles of Seven Different Varieties of Brown Rice (Oryza sativa L.). Foods 2022, 11, 1552. [Google Scholar] [CrossRef]
- Jan, B.; Zahiruddin, S.; Basist, P.; Khan, M.U.; Abass, S.; Dewangan, R.P.; Ahmad, S. Separation and Identification of Antioxidant and Antihyperglycemic Constituents from Morus alba Infusion by TLC-MS-Bioautography and UPLC-MS. J. Food Nutr. Res. 2021, 9, 670–690. [Google Scholar] [CrossRef]
- Guo, X.-L.; Yang, L.; Yu, S.-L.; Zhang, K.; Wang, J.-H.; Wang, H.-Y. Analysis of secondary metabolites in Xinjiang Morus nigra leaves using different extraction methods with UPLC-Q/TOF-MS/MS technology. Open Chem. 2023, 21, 20220320. [Google Scholar] [CrossRef]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Fraczek, M.; Kurpisz, M. Mechanisms of the harmful effects of bacterial semen infection on ejaculated human spermatozoa: Potential inflammatory markers in semen. Folia Histochem. Et Cytobiol. 2015, 53, 201–217. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef]
- Sanganalmath, S.K.; Dubey, S.; Veeranki, S.; Narisetty, K.; Krishnamurthy, P. The interplay of inflammation, exosomes and Ca2+ dynamics in diabetic cardiomyopathy. Cardiovasc. Diabetol. 2023, 22, 37. [Google Scholar] [CrossRef]
- Ren, L.; Charbord, J.; Chu, L.; Kemas, A.M.; Bertuzzi, M.; Mi, J.; Xing, C.; Lauschke, V.M.; Andersson, O. Adjudin improves beta cell maturation, hepatic glucose uptake and glucose homeostasis. Diabetologia 2023, 67, 137–155. [Google Scholar] [CrossRef]
- Iheagwam, F.N.; Iheagwam, O.T.; Onuoha, M.K.; Ogunlana, O.O.; Chinedu, S.N. Terminalia catappa aqueous leaf extract reverses insulin resistance, improves glucose transport and activates PI3K/AKT signalling in high fat/streptozotocin-induced diabetic rats. Sci. Rep. 2022, 12, 10711. [Google Scholar] [CrossRef]
- Zhao, H.; Avena-Bustillos, R.J.; Wang, S.C. Extraction, Purification and In Vitro Antioxidant Activity Evaluation of Phenolic Compounds in California Olive Pomace. Foods 2022, 11, 174. [Google Scholar] [CrossRef]
- Alabi, T.D.; Brooks, N.L.; Oguntibeju, O.O. Antioxidant Capacity, Phytochemical Analysis and Identification of Active Compounds in Anchomanes difformis. Nat. Prod. J. 2019, 10, 446–458. [Google Scholar] [CrossRef]
- Zubarev, R.A.; Makarov, A. Orbitrap mass spectromentry. Anal. Chem. 2013, 85, 5288–5296. [Google Scholar] [CrossRef]
- Idris, O.A.; Kerebba, N.; Horn, S.; Maboeta, M.S.; Pieters, R. Comparative phytochemistry using UPLC-ESI-QTOF-MS phenolic compounds profile of the water and aqueous ethanol extracts of Tagetes minuta and their cytotoxicity. S. Afr. J. Bot. 2024, 164, 50–65. [Google Scholar] [CrossRef]
- van de Venter, M.; Roux, S.; Bungu, L.C.; Louw, J.; Crouch, N.R.; Grace, O.M.; Maharaj, V.; Pillay, P.; Sewnarian, P.; Bhagwandin, N.; et al. Antidiabetic screening and scoring of 11 plants traditionally used in South Africa. J. Ethnopharmacol. 2008, 119, 81–86. [Google Scholar] [CrossRef]
- Pringle, N.A.; van de Venter, M.; Koekemoer, T.C. Comprehensive in vitro antidiabetic screening of Aspalathus linearis using a target-directed screening platform and cellomics. Food Funct. 2021, 12, 1020–1038. [Google Scholar] [CrossRef]
- Kwon, Y.; Apostolidis, E.; Shetty, K. Inhibitory potential of wine and tea against α-amylase and α-glucosidase for management of hyperglycemia linked to type 2 diabetes. J. Food Biochem. 2006, 32, 15–31. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | tR (min) | UV λmax (nm) | m/z [M-H]− | MS/MS | Tentative Name | References |
|---|---|---|---|---|---|---|
| 1 | 1.54 | 215.0224/225.0520 | 179, 165, 133 | Dimethoxycinnamic acid monohydrate | [47] | |
| 2 | 1.68 | 188.9939 | 127 | 2,6-Diaminopimelic acid | [48] | |
| 3 | 2.21 | 262, 290 | 191.0101 | 179, 119, 149 | p-Coumaric acid ethyl ester | [49] |
| 4 | 14.20 | 254, 290 | 329.0814 | 167, 279 | Vanilic acid hexoside | [50] |
| 5 | 14.38 | 286, 310 | 423.0862 | 315, 291 | Unidentified | - |
| 6 | 14.65 | 278, 311 | 469.1545 | 423, 391, 343, 203 | Unidentified | - |
| 7 | 14.98 | 256, 301 | 361.0706 | 272, 193 | 2,4,6-Trihydroxy-21,51-dimethoxybenzophenone derivative | [38] |
| 8 | 15.44 | 236, 314 | 451.2101 | 351, 272, 361 | 31-β-Glucosyloxy-4,41-dihydroxy-2,6-dimethoxy-benzophenone | [38] |
| 9 | 16.13 | 271 | 305.0615 | 236, 162 | (±)-Gallocatechin | [50] |
| 10 | 16.72 | 285 sh | 583.2595 | 451, 293 | 3-hydroxyphloretin 2′-O-xylosylglucoside | [51] |
| 11 | 16.93 | 281 | 451.2140 | 387, 441, 113, 45 | Epicatechin-3-O- hexoside | [52] |
| 12 | 17.36 | 264 | 417.0993 | 159 | Kaempferol-3-O-pentoside | [53] |
| 13 | 17.89 | 280 | 577.1384 | 449 | Procyanidin B2 | [54] |
| 14 | 18.17 | 280 | 447.1834 | 407, 437, 261, 327 | Kaempferol-3-O-hexoside | [55] |
| 15 | 18.42 | 300 | 473.1236 | 439, 215 | 3-O-malonyl-5-O-(E)-caffeoylquinic acid derivative | [42] |
| 16 | 18.51 | 281 | 439.1740 | 239 | 3-O-malonyl-5-O-(E)-caffeoylquinic acid | [42] |
| 17 | 18.63 | 252, 289 | 431.1884 | 289, 421 | Epicatechin-3-O-xylosyl derivative | - |
| 18 | 18.71 | 279 | 289.0652 | 271, 215 | Epicatechin | [56] |
| 19 | 18.92 | 280 | 567.2673 | 431, 327, 521 | Epicatechin -3-O-(41-xylosyl) deohexoside | New |
| 20 | 19.02 | 281 | 435.2204 | 289, 425 | Catechin-3-rhamnoside | [57] |
| 21 | 19.42 | 267, 285, 337 | 435.2199 | 289, 431, 421 | Epicatechin-O-rhamnoside | [57] |
| 22 | 19.64 | 285 | 431.1862 | 385 | Hexoside of kaempferol-3-O-(deoxy) | [52,58] |
| 23 | 19.90 | 281 | 371.0909/441.1890 | 247, 309, 157 | (−)-Epicatechin-3-Ogallate | [59] |
| 24 | 21.77 | 270, 344 | 577.1569 | Un fragmented | Procyanidin B1 dimer | [60] |
| 25 | 21.84 | 268, 338 | 577.1582 | 413, 293, 311 | Procyanidin B11 dimer | [61] |
| 26 | 22.00 | 268, 335 | 431.0912 | Un fragmented | Epicatechin-3-O- xylosyl derivative | - |
| 27 | 22.34 | 270, 335 | 431.0898 | 289, 317 | Epicatechin-3-O- xylosyl derivative | - |
| 28 | 22.47 | 270, 334 | 316.9897 | 237, 207, 283 | Isorhamnetin | [62] |
| 29 | 22.68 | 292 | 300.9943/563.2272 | 221, 157 | Apigenin-C-pentoside-C-hexoside | [57,58] |
| 30 | 22.80 | 551.2685 | 447, 301, 541, 245, 505 | O-Methyl-gallocatechin-3-O-gallate-O-sulfates | [63] | |
| 31 | 22.92 | 551.2707 | 301, 447, 401, 541, 319, 169 | O-Methyl-(epi)gallocatechin-3-O-gallate-O-sulfate | [63] | |
| 32 | 23.49 | 553.2888 [M-2Na]− | 419, 507 | Quercetin-3-O-acetyl-hexoside | [58] | |
| 33 | 24.31 | 279 | 187.0867 | 169, 127 | Gallic acid | [56,64] |
| 34 | 24.69 | 227.1176 | Not identified | |||
| 35 | 25.29 | 329.0599 | 150, 145 | Fagomine derivative | [65] | |
| 36 | 25.62 | 302 | 149.9842 | Fagomine | [65,66] | |
| 37 | 27.07 | 238.8820 | Unidentified |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nethengwe, M.; Kerebba, N.; Okaiyeto, K.; Opuwari, C.S.; Oguntibeju, O.O. Antioxidant, Anti-Diabetic, and Anti-Inflammation Activity of Garcinia livingstonei Aqueous Leaf Extract: A Preliminary Study. Int. J. Mol. Sci. 2024, 25, 3184. https://doi.org/10.3390/ijms25063184
Nethengwe M, Kerebba N, Okaiyeto K, Opuwari CS, Oguntibeju OO. Antioxidant, Anti-Diabetic, and Anti-Inflammation Activity of Garcinia livingstonei Aqueous Leaf Extract: A Preliminary Study. International Journal of Molecular Sciences. 2024; 25(6):3184. https://doi.org/10.3390/ijms25063184
Chicago/Turabian StyleNethengwe, Murendeni, Nasifu Kerebba, Kunle Okaiyeto, Chinyerum S. Opuwari, and Oluwafemi O. Oguntibeju. 2024. "Antioxidant, Anti-Diabetic, and Anti-Inflammation Activity of Garcinia livingstonei Aqueous Leaf Extract: A Preliminary Study" International Journal of Molecular Sciences 25, no. 6: 3184. https://doi.org/10.3390/ijms25063184