
Trimetazidine Improves Mitochondrial Dysfunction in SOD1G93A Cellular Models of Amyotrophic Lateral Sclerosis through Autophagy Activation
 , , , ,
, , , ,  ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Mitochondrial Functionality Is Impaired in Primary Spinal Cord and Cortical Cell Cultures
2.2. Acute Trimetazidine Treatments Protect Mitochondria in Primary Spinal and Cortical Cell Cultures
2.3. Ultrastructural Mitochondrial Morphology Is Preserved in Primary Spinal and Cortical Cell Cultures Administered with Trimetazidine Treatment
2.4. Trimetazidine Treatment Stimulates Autophagy and Mitophagy Processes
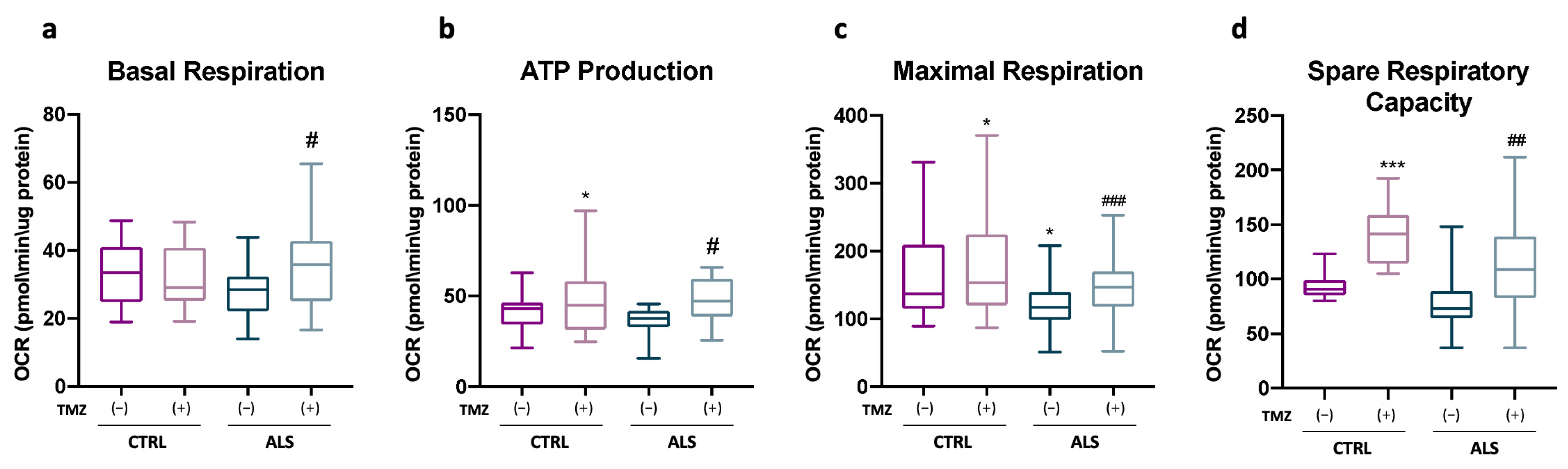
2.5. Trimetazidine Treatment Ameliorates Mitochondrial Functionality of ALS-Patient-Derived PBMCs
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Animals
4.3. Primary Cell Cultures and Cell Lines
4.4. Amyotrophic Lateral Sclerosis Patient Biosampling and PBMC Preparation
4.5. Electrophoresis and Western Blotting
4.6. Immunofluorescence Analysis
4.7. Bioenergetic Analysis
4.8. Spectrophotometric Assays
4.9. Electron Microscopic Analysis
4.10. Mitochondria Morphological Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldman, E.L.; Goutman, S.A.; Petri, S.; Mazzini, L.; Savelieff, M.G.; Shaw, P.J.; Sobue, G. Amyotrophic Lateral Sclerosis. Lancet 2022, 400, 1363–1380. [Google Scholar] [CrossRef] [PubMed]
- Genge, A.; Wainwright, S.; Velde, C. Vande Amyotrophic Lateral Sclerosis: Exploring Pathophysiology in the Context of Treatment. Amyotroph. Lateral Scler. Front. Degener. 2023, 1–12. [Google Scholar] [CrossRef]
- Levi Montalcini, R.; Feldman, E.L.; Goutman, S.A.; Hardiman, O.; Al-Chalabi, A.; Chió, A.; Savelieff, M.G.; Kiernan, M.C. Amyotrophic Lateral Sclerosis 2 Recent Advances in the Diagnosis and Prognosis of Amyotrophic Lateral Sclerosis. Ser. Lancet Neurol. 2022, 21, 480–493. [Google Scholar] [CrossRef]
- Akçimen, F.; Lopez, E.R.; Landers, J.E.; Nath, A.; Chiò, A.; Chia, R.; Traynor, B.J. Amyotrophic Lateral Sclerosis: Translating Genetic Discoveries into Therapies. Nat. Rev. Genet. 2023, 24, 642–658. [Google Scholar] [CrossRef] [PubMed]
- Talbott, E.O.; Malek, A.M.; Lacomis, D. The Epidemiology of Amyotrophic Lateral Sclerosis. Handb. Clin. Neurol. 2016, 138, 225–238. [Google Scholar] [CrossRef]
- Ferri, A.; Coccurello, R. What Is “Hyper” in the ALS Hypermetabolism? Mediat. Inflamm. 2017, 2017, 7821672. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.T.; Trotti, D. Altered Bioenergetics and Metabolic Homeostasis in Amyotrophic Lateral Sclerosis. Neurotherapeutics 2022, 19, 1102–1118. [Google Scholar] [CrossRef]
- Peter, R.S.; Rosenbohm, A.; Dupuis, L.; Brehme, T.; Kassubek, J.; Rothenbacher, D.; Nagel, G.; Ludolph, A.C. Life Course Body Mass Index and Risk and Prognosis of Amyotrophic Lateral Sclerosis: Results from the ALS Registry Swabia. Eur. J. Epidemiol. 2017, 32, 901–908. [Google Scholar] [CrossRef]
- D’amico, E.; Grosso, G.; Nieves, J.W.; Zanghì, A.; Factor-Litvak, P.; Mitsumoto, H. Metabolic Abnormalities, Dietary Risk Factors and Nutritional Management in Amyotrophic Lateral Sclerosis. Nutrients 2021, 13, 2273. [Google Scholar] [CrossRef]
- Ludolph, A.; Dupuis, L.; Kasarskis, E.; Steyn, F.; Ngo, S.; Mcdermott, C. Nutritional and Metabolic Factors in Amyotrophic Lateral Sclerosis. Nat. Rev. Neurol. 2023, 19, 511–524. [Google Scholar] [CrossRef]
- Dorst, J.; Weydt, P.; Brenner, D.; Witzel, S.; Kandler, K.; Huss, A.; Herrmann, C.; Wiesenfarth, M.; Knehr, A.; Günther, K.; et al. Metabolic Alterations Precede Neurofilament Changes in Presymptomatic ALS Gene Carriers. EBioMedicine 2023, 90, 104521. [Google Scholar] [CrossRef]
- Steyn, F.J.; Ioannides, Z.A.; Van Eijk, R.P.A.; Heggie, S.; Thorpe, K.A.; Ceslis, A.; Heshmat, S.; Henders, A.K.; Wray, N.R.; Van Den Berg, L.H.; et al. Hypermetabolism in ALS Is Associated with Greater Functional Decline and Shorter Survival. J. Neurol. Neurosurg. Psychiatry 2018, 89, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Perera, N.D.; Bradley, J.; Turner, J. AMPK Signalling and Defective Energy Metabolism in Amyotrophic Lateral Sclerosis. Neurochem. Res. 2016, 41, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Scaricamazza, S.; Salvatori, I.; Giacovazzo, G.; Loeffler, J.P.; Renè, F.; Rosina, M.; Quessada, C.; Proietti, D.; Heil, C.; Rossi, S.; et al. Skeletal-Muscle Metabolic Reprogramming in ALS-SOD1 G93A Mice Predates Disease Onset and Is A Promising Therapeutic Target. iScience 2020, 23, 101087. [Google Scholar] [CrossRef] [PubMed]
- Steyn, F.J.; Li, R.; Kirk, S.E.; Tefera, T.W.; Xie, T.Y.; Tracey, T.J.; Kelk, D.; Wimberger, E.; Garton, F.C.; Roberts, L.; et al. Altered Skeletal Muscle Glucose-Fatty Acid Flux in Amyotrophic Lateral Sclerosis. Brain Commun. 2020, 2, fcaa154. [Google Scholar] [CrossRef]
- Michels, S.; Kurz, D.; Rosenbohm, A.; Peter, R.S.; Just, S.; Bäzner, H.; Börtlein, A.; Dettmers, C.; Gold, H.J.; Kohler, A.; et al. Association of Blood Lipids with Onset and Prognosis of Amyotrophic Lateral Sclerosis: Results from the ALS Swabia Registry. J. Neurol. 2023, 270, 3082–3090. [Google Scholar] [CrossRef]
- Smith, E.F.; Shaw, P.J.; De Vos, K.J. The Role of Mitochondria in Amyotrophic Lateral Sclerosis. Neurosci. Lett. 2019, 710, 132933. [Google Scholar] [CrossRef] [PubMed]
- Genin, E.C.; Abou-Ali, M.; Paquis-Flucklinger, V. Mitochondria, a Key Target in Amyotrophic Lateral Sclerosis Pathogenesis. Genes 2023, 14, 1981. [Google Scholar] [CrossRef]
- Belosludtseva, N.V.; Matveeva, L.A.; Belosludtsev, K.N. Mitochondrial Dyshomeostasis as an Early Hallmark and a Therapeutic Target in Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2023, 24, 16833. [Google Scholar] [CrossRef]
- Dhasmana, S.; Dhasmana, A.; Kotnala, S.; Mangtani, V.; Narula, A.S.; Haque, S.; Jaggi, M.; Yallapu, M.M.; Chauhan, S.C. Boosting Mitochondrial Potential: An Imperative Therapeutic Intervention in Amyotrophic Lateral Sclerosis. Curr. Neuropharmacol. 2022, 21, 1117–1138. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Huo, Z.; Chen, Y.; Liu, J.; Zhao, Z.; Meng, F.; Su, Q.; Bao, W.; Zhang, L.; et al. The Impact of Mitochondrial Dysfunction in Amyotrophic Lateral Sclerosis. Cells 2022, 11, 2049. [Google Scholar] [CrossRef]
- Shu, H.; Peng, Y.; Hang, W.; Zhou, N.; Wang, D.W. Trimetazidine in Heart Failure. Front. Pharmacol. 2021, 11, 569132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; You, B.; Qi, D.; Qiu, L.; Ripley-Gonzalez, J.W.; Zheng, F.; Fu, S.; Li, C.; Dun, Y.; Liu, S. Trimetazidine and Exercise Provide Comparable Improvements to High Fat Diet-Induced Muscle Dysfunction through Enhancement of Mitochondrial Quality Control. Sci. Rep. 2021, 11, 19116. [Google Scholar] [CrossRef] [PubMed]
- Kantor, P.F.; Lucien, A.; Kozak, R.; Lopaschuk, G.D. The Antianginal Drug Trimetazidine Shifts Cardiac Energy Metabolism from Fatty Acid Oxidation to Glucose Oxidation by Inhibiting Mitochondrial Long-Chain 3-Ketoacyl Coenzyme A Thiolase. Circ. Res. 2000, 86, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Kalucka, J.; Bierhansl, L.; Conchinha, N.V.; Missiaen, R.; Elia, I.; Brüning, U.; Scheinok, S.; Treps, L.; Cantelmo, A.R.; Dubois, C.; et al. Quiescent Endothelial Cells Upregulate Fatty Acid β-Oxidation for Vasculoprotection via Redox Homeostasis. Cell Metab. 2018, 28, 881–894.e13. [Google Scholar] [CrossRef]
- Argaud, L.; Gomez, L.; Gateau-Roesch, O.; Couture-Lepetit, E.; Loufouat, J.; Robert, D.; Ovize, M. Trimetazidine Inhibits Mitochondrial Permeability Transition Pore Opening and Prevents Lethal Ischemia-Reperfusion Injury. J. Mol. Cell Cardiol. 2005, 39, 893–899. [Google Scholar] [CrossRef]
- Morin, D.; Elimadi, A.; Sapena, R.; Crevat, A.; Carrupt, P.A.; Testa, B.; Tillement, J.P. Evidence for the Existence of [3H]-Trimetazidine Binding Sites Involved in the Regulation of the Mitochondrial Permeability Transition Pore. Br. J. Pharmacol. 1998, 123, 1385–1394. [Google Scholar] [CrossRef]
- Marzilli, M.; Vinereanu, D.; Lopaschuk, G.; Chen, Y.; Dalal, J.J.; Danchin, N.; Etriby, E.; Ferrari, R.; Gowdak, L.H.; Lopatin, Y.; et al. Trimetazidine in Cardiovascular Medicine. Int. J. Cardiol. 2019, 293, 39–44. [Google Scholar] [CrossRef]
- Abozguia, K.; Clarke, K.; Lee, L.; Frenneaux, M. Modification of Myocardial Substrate Use as a Therapy for Heart Failure. Nat. Clin. Pract. Cardiovasc. Med. 2006, 3, 490–498. [Google Scholar] [CrossRef]
- Trounce, I.A.; Kim, Y.L.; Jun, A.S.; Wallace, D.C. Assessment of Mitochondrial Oxidative Phosphorylation in Patient Muscle Biopsies, Lymphoblasts, and Transmitochondrial Cell Lines. Methods Enzymol. 1996, 264, 484–509. [Google Scholar] [CrossRef]
- Molinari, F.; Pin, F.; Gorini, S.; Chiandotto, S.; Pontecorvo, L.; Penna, F.; Rizzuto, E.; Pisu, S.; Musarò, A.; Costelli, P.; et al. The Mitochondrial Metabolic Reprogramming Agent Trimetazidine as an “exercise Mimetic” in Cachectic C26-Bearing Mice. J. Cachexia Sarcopenia Muscle 2017, 8, 954–973. [Google Scholar] [CrossRef]
- Belli, R.; Bonato, A.; De Angelis, L.; Mirabilii, S.; Ricciardi, M.R.; Tafuri, A.; Molfino, A.; Leigheb, M.; Costelli, P.; Caruso, M.; et al. Metabolic Reprogramming Promotes Myogenesis During Aging. Front. Physiol. 2019, 10, 442450. [Google Scholar] [CrossRef] [PubMed]
- Scaricamazza, S.; Salvatori, I.; Amadio, S.; Nesci, V.; Torcinaro, A.; Giacovazzo, G.; Primiano, A.; Gloriani, M.; Candelise, N.; Pieroni, L.; et al. Repurposing of Trimetazidine for Amyotrophic Lateral Sclerosis: A Study in SOD1G93A Mice. Br. J. Pharmacol. 2022, 179, 1732. [Google Scholar] [CrossRef]
- Davoli, A.; Greco, V.; Spalloni, A.; Guatteo, E.; Neri, C.; Rizzo, G.R.; Cordella, A.; Romigi, A.; Cortese, C.; Bernardini, S.; et al. Evidence of Hydrogen Sulfide Involvement in Amyotrophic Lateral Sclerosis. Ann. Neurol. 2015, 77, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Spalloni, A.; Albo, F.; Ferrari, F.; Mercuri, N.; Bernardi, G.; Zona, C.; Longone, P. Cu/Zn-Superoxide Dismutase (GLY93→ALA) Mutation Alters AMPA Receptor Subunit Expression and Function and Potentiates Kainate-Mediated Toxicity in Motor Neurons in Culture. Neurobiol. Dis. 2004, 15, 340–350. [Google Scholar] [CrossRef]
- Chang, Q.; Martin, L.J. Voltage-Gated Calcium Channels Are Abnormal in Cultured Spinal Motoneurons in the G93A-SOD1 Transgenic Mouse Model of ALS. Neurobiol. Dis. 2016, 93, 78–95. [Google Scholar] [CrossRef]
- Zhang, J.; He, X.; Bai, X.; Sun, Y.; Jiang, P.; Wang, X.; Li, W.; Zhang, Y. Protective Effect of Trimetazidine in Radiation-Induced Cardiac Fibrosis in Mice. J. Radiat. Res. 2020, 61, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, N.; Chen, T.; Zhang, C.; Liu, L.; Qi, Y.; Bu, P. Trimetazidine Ameliorates Sunitinib-Induced Cardiotoxicity in Mice via the AMPK/MTOR/Autophagy Pathway. Pharm. Biol. 2019, 57, 625–631. [Google Scholar] [CrossRef]
- El-khodary, N.M.; Ghoneim, A.I.; El-tayaar, A.A.; El-touny, E.M. The Impact of Trimetazidine on Cardiac Fibrosis, Inflammation, and Function in Ischemic Cardiomyopathy Patients. Cardiovasc. Drugs Ther. 2023, 37, 955–964. [Google Scholar] [CrossRef]
- Li, H.; Ma, Z.; Zhai, Y.; Lv, C.; Yuan, P.; Zhu, F.; Wei, L.; Li, Q.; Qi, X. Trimetazidine Ameliorates Myocardial Metabolic Remodeling in Isoproterenol-Induced Rats Through Regulating Ketone Body Metabolism via Activating AMPK and PPAR α. Front. Pharmacol. 2020, 11, 568526. [Google Scholar] [CrossRef]
- Cavar, M.; Ljubkovic, M.; Bulat, C.; Bakovic, D.; Fabijanic, D.; Kraljevic, J.; Karanovic, N.; Dujic, Z.; Lavie, C.J.; Wisloff, U.; et al. Trimetazidine Does Not Alter Metabolic Substrate Oxidation in Cardiac Mitochondria of Target Patient Population. Br. J. Pharmacol. 2016, 173, 1529–1540. [Google Scholar] [CrossRef]
- Jiang, L.; Shen, X.; Dun, Y.; Xie, M.; Fu, S.; Zhang, W.; Qiu, L.; Ripley-Gonzalez, J.W.; Liu, S. Exercise Combined with Trimetazidine Improves Anti-Fatal Stress Capacity through Enhancing Autophagy and Heat Shock Protein 70 of Myocardium in Mice. Int. J. Med. Sci. 2021, 18, 1680–1686. [Google Scholar] [CrossRef]
- Zhou, X.; Li, C.; Xu, W.; Chen, J. Trimetazidine Protects against Smoking-Induced Left Ventricular Remodeling via Attenuating Oxidative Stress, Apoptosis, and Inflammation. PLoS ONE 2012, 7, e40424. [Google Scholar] [CrossRef]
- Eid, B.G.; El-Shitany, N.A.E.A.; Neamatallah, T. Trimetazidine Improved Adriamycin-Induced Cardiomyopathy by Downregulating TNF-α, BAX, and VEGF Immunoexpression via an Antioxidant Mechanism. Environ. Toxicol. 2021, 36, 1217–1225. [Google Scholar] [CrossRef]
- Ferraro, E.; Pin, F.; Gorini, S.; Pontecorvo, L.; Ferri, A.; Mollace, V.; Costelli, P.; Rosano, G. Improvement of Skeletal Muscle Performance in Ageing by the Metabolic Modulator Trimetazidine. J. Cachexia. Sarcopenia Muscle 2016, 7, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Bortolasci, C.C.; Kidnapillai, S.; Spolding, B.; Truong, T.T.T.; Connor, T.; Swinton, C.; Panizzutti, B.; Liu, Z.S.J.; Sanigorski, A.; Dean, O.M.; et al. Use of a Gene Expression Signature to Identify Trimetazidine for Repurposing to Treat Bipolar Depression. Bipolar Disord. 2023, 25, 661–670. [Google Scholar] [CrossRef]
- Dhote, V.; Mandloi, A.S.; Singour, P.K.; Kawadkar, M.; Ganeshpurkar, A.; Jadhav, M.P. Neuroprotective Effects of Combined Trimetazidine and Progesterone on Cerebral Reperfusion Injury. Curr. Res. Pharmacol. Drug Discov. 2022, 3, 100108. [Google Scholar] [CrossRef]
- Al-Shorbagy, M.Y.; Wadie, W.; El-Tanbouly, D.M. Trimetazidine Modulates Mitochondrial Redox Status and Disrupted Glutamate Homeostasis in a Rat Model of Epilepsy. Front. Pharmacol. 2021, 12, 735165. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.A.E.; Abdel-Rahman, R.F.; Mahmoud, S.S.; Khattab, M.M.; Safar, M.M. Metformin and Trimetazidine Ameliorate Diabetes-Induced Cognitive Impediment in Status Epileptic Rats. Epilepsy Behav. 2020, 104, 106893. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; Su, W.; Zhang, Y.; Li, Z.; Deng, C.; Zhuo, Y. Trimetazidine Protects Retinal Ganglion Cells from Acute Glaucoma via the Nrf2/Ho-1 Pathway. Clin. Sci. 2017, 131, 2363–2375. [Google Scholar] [CrossRef]
- Serarslan, Y.; Bal, R.; Altug, M.E.; Kontaş, T.; Keleş, O.N.; Unal, D.; Unal, B. Effects of Trimetazidine on Crush Injury of the Sciatic Nerve in Rats: A Biochemical and Stereological Study. Brain Res. 2009, 1247, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Browne, S.E.; Yang, L.; DiMauro, J.P.; Fuller, S.W.; Licata, S.C.; Beal, M.F. Bioenergetic Abnormalities in Discrete Cerebral Motor Pathways Presage Spinal Cord Pathology in the G93A SOD1 Mouse Model of ALS. Neurobiol. Dis. 2006, 22, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Ferri, A.; Cozzolino, M.; Crosio, C.; Nencini, M.; Casciati, A.; Gralla, E.B.; Rotilio, G.; Valentine, J.S.; Carri, M.T. Familial ALS-Superoxide Dismutases Associate with Mitochondria and Shift Their Redox Potentials. Proc. Natl. Acad. Sci. USA 2006, 103, 13860–13865. [Google Scholar] [CrossRef]
- Pieri, M.; Albo, F.; Gaetti, C.; Spalloni, A.; Bengtson, C.P.; Longone, P.; Cavalcanti, S.; Zona, C. Altered Excitability of Motor Neurons in a Transgenic Mouse Model of Familial Amyotrophic Lateral Sclerosis. Neurosci. Lett. 2003, 351, 153–156. [Google Scholar] [CrossRef]
- Pieri, M.; Carunchio, I.; Curcio, L.; Mercuri, N.B.; Zona, C. Increased Persistent Sodium Current Determines Cortical Hyperexcitability in a Genetic Model of Amyotrophic Lateral Sclerosis. Exp. Neurol. 2009, 215, 368–379. [Google Scholar] [CrossRef]
- Le Masson, G.; Przedborski, S.; Abbott, L.F. A Computational Model of Motor Neuron Degeneration. Neuron 2014, 83, 975–988. [Google Scholar] [CrossRef]
- Ferraro, E.; Giammarioli, A.M.; Caldarola, S.; Lista, P.; Feraco, A.; Tinari, A.; Salvatore, A.M.; Malorni, W.; Berghella, L.; Rosano, G. The Metabolic Modulator Trimetazidine Triggers Autophagy and Counteracts Stress-Induced Atrophy in Skeletal Muscle Myotubes. FEBS J. 2013, 280, 5094–5108. [Google Scholar] [CrossRef]
- Gatta, L.; Vitiello, L.; Gorini, S.; Chiandotto, S.; Costelli, P.; Giammarioli, A.M.; Malorni, W.; Rosano, G.; Ferraro, E. Modulating the Metabolism by Trimetazidine Enhances Myoblast Differentiation and Promotes Myogenesis in Cachectic Tumorbearing C26 Mice. Oncotarget 2017, 8, 113938–113956. [Google Scholar] [CrossRef]
- De Silva, S.; Turner, B.J.; Perera, N.D. Metabolic Dysfunction in Motor Neuron Disease: Shedding Light through the Lens of Autophagy. Metabolites 2022, 12, 574. [Google Scholar] [CrossRef]
- Cirulli, E.T.; Lasseigne, B.N.; Petrovski, S.; Sapp, P.C.; Dion, P.A.; Leblond, C.S.; Couthouis, J.; Lu, Y.F.; Wang, Q.; Krueger, B.J.; et al. Exome Sequencing in Amyotrophic Lateral Sclerosis Identifies Risk Genes and Pathways. Science 2015, 347, 1436–1441. [Google Scholar] [CrossRef]
- Freischmidt, A.; Wieland, T.; Richter, B.; Ruf, W.; Schaeffer, V.; Müller, K.; Marroquin, N.; Nordin, F.; Hübers, A.; Weydt, P.; et al. Haploinsufficiency of TBK1 Causes Familial ALS and Fronto-Temporal Dementia. Nat. Neurosci. 2015, 18, 631–636. [Google Scholar] [CrossRef]
- Maruyama, H.; Kawakami, H. Optineurin and Amyotrophic Lateral Sclerosis. Geriatr. Gerontol. Int. 2013, 13, 528–532. [Google Scholar] [CrossRef]
- Evans, C.S.; Holzbaur, E.L.F. Autophagy and Mitophagy in ALS. Neurobiol. Dis. 2019, 122, 35–40. [Google Scholar] [CrossRef]
- Tak, Y.J.; Park, J.H.; Rhim, H.; Kang, S. ALS-Related Mutant SOD1 Aggregates Interfere with Mitophagy by Sequestering the Autophagy Receptor Optineurin. Int. J. Mol. Sci. 2020, 21, 7525. [Google Scholar] [CrossRef]
- Magrì, A.; Lipari, C.L.R.; Risiglione, P.; Zimbone, S.; Guarino, F.; Caccamo, A.; Messina, A. ERK1/2-Dependent TSPO Overactivation Associates with the Loss of Mitophagy and Mitochondrial Respiration in ALS. Cell Death Dis. 2023, 14, 122. [Google Scholar] [CrossRef]
- Bordoni, M.; Pansarasa, O.; Scarian, E.; Cristofani, R.; Leone, R.; Fantini, V.; Garofalo, M.; Diamanti, L.; Bernuzzi, S.; Gagliardi, S.; et al. Lysosomes Dysfunction Causes Mitophagy Impairment in PBMCs of Sporadic ALS Patients. Cells 2022, 11, 1272. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.R.; Walters, R.; Waldron, F.M.; Pal, S.; Selvaraj, B.T.; Macleod, M.R.; Hardingham, G.E.; Chandran, S.; Gregory, J.M. Targeting Mitochondrial Dysfunction in Amyotrophic Lateral Sclerosis: A Systematic Review and Meta-Analysis. Brain Commun. 2019, 1, fcz009. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Miller, R.G.; Swash, M.; Munsat, T.L. El Escorial Revisited: Revised Criteria for the Diagnosis of Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler Other Mot. Neuron. Disord. 2000, 1, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Schirinzi, T.; Salvatori, I.; Zenuni, H.; Grillo, P.; Valle, C.; Martella, G.; Mercuri, N.B.; Ferri, A. Pattern of Mitochondrial Respiration in Peripheral Blood Cells of Patients with Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 10863. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, I.; Ferri, A.; Scaricamazza, S.; Giovannelli, I.; Serrano, A.; Rossi, S.; D’Ambrosi, N.; Cozzolino, M.; Di Giulio, A.; Moreno, S.; et al. Differential Toxicity of TAR DNA-Binding Protein 43 Isoforms Depends on Their Submitochondrial Localization in Neuronal Cells. J. Neurochem. 2018, 146, 585–597. [Google Scholar] [CrossRef]
- Valente, A.J.; Maddalena, L.A.; Robb, E.L.; Moradi, F.; Stuart, J.A. A Simple ImageJ Macro Tool for Analyzing Mitochondrial Network Morphology in Mammalian Cell Culture. Acta. Histochem. 2017, 119, 315–326. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Sources | Product | Dilution | |
|---|---|---|---|---|
| Primary | Cu/Zn SOD | Rabbit polyclonal | Enzo (Lausen, Switzerland) (Cat#ADI-SOD-100D)) | WB 1:1000 |
| β-actin | Mouse monoclonal | Sigma-Aldrich (Darmstadt, Germany) (Cat #MA1-140) | WB 1:1000 | |
| TOM20 | Rabbit polyclonal | Santa Cruz Biotechnology (Dallas, TX, USA) (Cat# sc-11415) | WB 1:1000 | |
| VDAC | Mouse monoclonal | Santa Cruz Biotechnology (Dallas, TX, USA) (Cat# sc-8829) | WB 1:1000 | |
| SMI-32 | Mouse monoclonal | BioLegend (San Diego, CA, USA) (formerly Covance Antibody Products) (Cat# SMI-32P) | IF 1:1000 | |
| Map2 | Mouse monoclonal | Invitrogen (Carlsbad, CA, USA) (Cat# MA5-12826) | IF 1:500 | |
| Connexin 43 | Rabbit monoclonal | Sigma-Aldrich (Darmstadt, Germany) (Cat#ZRB1179-25UL) | WB 1:1000 | |
| GFAP | Rabbit polyclonal | Invitrogen (Carlsbad, CA, USA) (Cat# PA5-16291) | IF 1:1000 | |
| LC3B | Rabbit polyclonal | Novus Biologicals (Cambridge, UK) (Cat# NB100-2220) | WB 1:1000 | |
| phospho-Ubiquitin (Ser65) | Rabbit monoclonal | Cell Signaling Technologies (Danvers, MA, USA) (Cat# BK62802) | WB 1:1000 | |
| HSP90 | Mouse monoclonal | Santa Cruz Biotechnologies (Dallas, TX, USA) (Cat# SC13119) | WB 1:1000 | |
| Secondary | HRP Conjugate | Goat anti-rabbit | Bio-Rad Laboratories (Richmond, CA, USA) (Cat#1706515; RRID: AB_2617112) | WB 1:2000 |
| HRP Conjugate | Goat anti-mouse | Bio-Rad Laboratories (Richmond, CA, USA) (Cat#170-6516, RRID: AB_11125547) | WB 1:2000 | |
| Alexa Fluor 488 | Donkey anti-mouse | Invitrogen (Carlsbad, CA, USA) (Cat# A11017) | IF 1:300 | |
| Cy™3 | Donkey anti-rabbit | Jackson Immuno-Research Laboratories (Farminton, CA, USA) (Cat# 65119) | IF 1:500 |
| Sample | Gender | Age | Disease Duration (Months) | BMI | Clinical Phenotype | ALS-FRS-R | King’s Stage |
|---|---|---|---|---|---|---|---|
| sALS1 | F | 82 | 22 | 18 | Spinal | 39 | 2 |
| sALS2 | M | 68 | 23 | 28 | Spinal | 34 | 3 |
| sALS3 | F | 70 | 7 | 22.1 | Spinal | 32 | 3 |
| sALS4 | F | 74 | 73 | 21.4 | Spinal | 13 | 4a |
| TARDBP | M | 39 | 5 | 19.6 | Spinal | 44 | 2 |
| sALS6 | F | 70 | 7 | 22.1 | Spinal | 32 | 3 |
| sALS7 | F | 73 | 35 | 27.6 | Spinal | 37 | 2 |
| sALS8 | M | 66 | 36 | 26.3 | Spinal | 46 | 2 |
| sALS9 | M | 75 | 24 | 32 | Spinal | 37 | 3 |
| sALS10 | F | 76 | 24 | 25.6 | Spinal | 42 | 1 |
| sALS11 | F | 78 | 18 | 24.5 | Bulbar | 33 | 2 |
| C9Orf72 | F | 58 | 19 | 21.9 | Spinal | 32 | 3 |
| sALS13 | M | 69 | 13 | 23.7 | Spinal | 42 | 2 |
| sALS14 | M | 55 | 12 | - | Spinal | 12 | 4a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvatori, I.; Nesci, V.; Spalloni, A.; Marabitti, V.; Muzzi, M.; Zenuni, H.; Scaricamazza, S.; Rosina, M.; Fenili, G.; Goglia, M.; et al. Trimetazidine Improves Mitochondrial Dysfunction in SOD1G93A Cellular Models of Amyotrophic Lateral Sclerosis through Autophagy Activation. Int. J. Mol. Sci. 2024, 25, 3251. https://doi.org/10.3390/ijms25063251
Salvatori I, Nesci V, Spalloni A, Marabitti V, Muzzi M, Zenuni H, Scaricamazza S, Rosina M, Fenili G, Goglia M, et al. Trimetazidine Improves Mitochondrial Dysfunction in SOD1G93A Cellular Models of Amyotrophic Lateral Sclerosis through Autophagy Activation. International Journal of Molecular Sciences. 2024; 25(6):3251. https://doi.org/10.3390/ijms25063251
Chicago/Turabian StyleSalvatori, Illari, Valentina Nesci, Alida Spalloni, Veronica Marabitti, Maurizio Muzzi, Henri Zenuni, Silvia Scaricamazza, Marco Rosina, Gianmarco Fenili, Mariangela Goglia, and et al. 2024. "Trimetazidine Improves Mitochondrial Dysfunction in SOD1G93A Cellular Models of Amyotrophic Lateral Sclerosis through Autophagy Activation" International Journal of Molecular Sciences 25, no. 6: 3251. https://doi.org/10.3390/ijms25063251
APA StyleSalvatori, I., Nesci, V., Spalloni, A., Marabitti, V., Muzzi, M., Zenuni, H., Scaricamazza, S., Rosina, M., Fenili, G., Goglia, M., Boffa, L., Massa, R., Moreno, S., Mercuri, N. B., Nazio, F., Longone, P., Ferri, A., & Valle, C. (2024). Trimetazidine Improves Mitochondrial Dysfunction in SOD1G93A Cellular Models of Amyotrophic Lateral Sclerosis through Autophagy Activation. International Journal of Molecular Sciences, 25(6), 3251. https://doi.org/10.3390/ijms25063251