
Perineuronal Nets in the CNS: Architects of Memory and Potential Therapeutic Target in Neuropsychiatric Disorders

 ,
, 
{kind=link}
{kind=link}
Abstract
1. Introduction
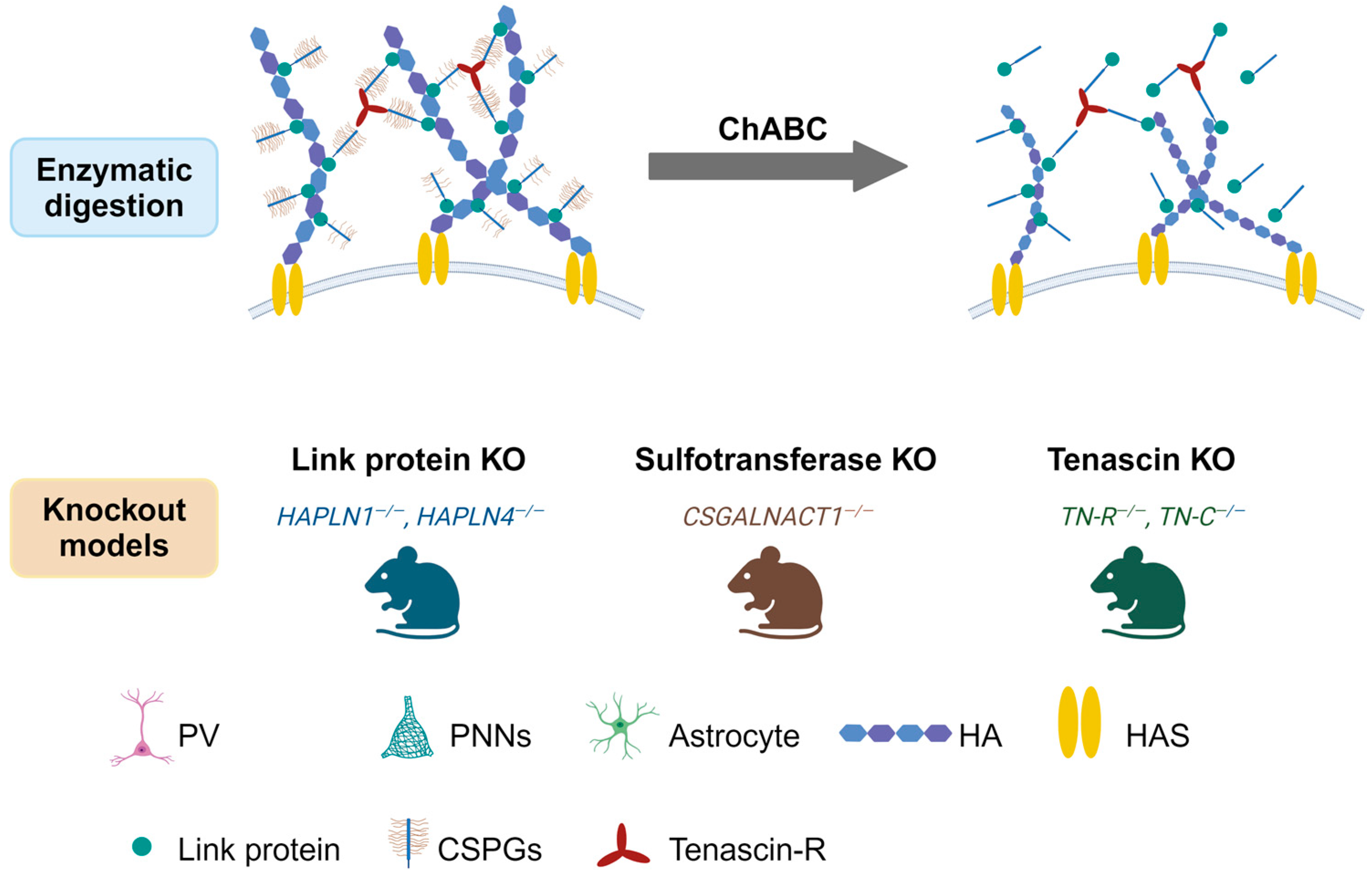
2. PNNs’ Structure and Functions
2.1. Composition and Distributions
2.2. Physiological Functions
3. Roles of PNNs in Learning and Memory
3.1. Object Recognition (OR) Memory
3.2. Fear Memory
3.3. Spatial Memory
4. Roles of PNNs in Memory-Related Dysfunctions
4.1. Addiction
4.2. Aging
4.3. Other
5. The Challenges and Prospects
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vargová, L.; Syková, E. Astrocytes and extracellular matrix in extrasynaptic volume transmission. Philos. Trans. R. Soc. London Ser. B Biol. Sci. 2014, 369, 20130608. [Google Scholar] [CrossRef]
- Pizzorusso, T.; Medini, P.; Berardi, N.; Chierzi, S.; Fawcett, J.; Maffei, L. Reactivation of ocular dominance plasticity in the adult visual cortex. Science 2002, 298, 1248–1251. [Google Scholar] [CrossRef]
- Massey, J.; Hubscher, C.; Wagoner, M.; Decker, J.; Amps, J.; Silver, J.; Onifer, S. Chondroitinase ABC digestion of the perineuronal net promotes functional collateral sprouting in the cuneate nucleus after cervical spinal cord injury. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 4406–4414. [Google Scholar] [CrossRef]
- Barritt, A.; Davies, M.; Marchand, F.; Hartley, R.; Grist, J.; Yip, P.; McMahon, S.; Bradbury, E. Chondroitinase ABC promotes sprouting of intact and injured spinal systems after spinal cord injury. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 10856–10867. [Google Scholar] [CrossRef]
- Yamada, J.; Jinno, S. Spatio-temporal differences in perineuronal net expression in the mouse hippocampus, with reference to parvalbumin. Neuroscience 2013, 253, 368–379. [Google Scholar] [CrossRef]
- O’Reilly, K.; Perica, M.; Fenton, A. Synaptic plasticity/dysplasticity, process memory and item memory in rodent models of mental dysfunction. Schizophr. Res. 2019, 207, 22–36. [Google Scholar] [CrossRef]
- Wanat, M.; Sparta, D.; Hopf, F.; Bowers, M.; Melis, M.; Bonci, A. Strain specific synaptic modifications on ventral tegmental area dopamine neurons after ethanol exposure. Biol. Psychiatry 2009, 65, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Grimwood, P.; Morris, R. Synaptic plasticity and memory: An evaluation of the hypothesis. Annu. Rev. Neurosci. 2000, 23, 649–711. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Cho, H.; Kim, M.; Oh, J.; Kang, M.; Yoo, M.; Lee, H.; Han, J. Synaptic plasticity-dependent competition rule influences memory formation. Nat. Commun. 2021, 12, 3915. [Google Scholar] [CrossRef]
- Ruzicka, J.; Dalecka, M.; Safrankova, K.; Peretti, D.; Jendelova, P.; Kwok, J.; Fawcett, J. Perineuronal nets affect memory and learning after synapse withdrawal. Transl. Psychiatry 2022, 12, 480. [Google Scholar] [CrossRef] [PubMed]
- Testa, D.; Prochiantz, A.; Di Nardo, A. Perineuronal nets in brain physiology and disease. Semin. Cell Dev. Biol. 2019, 89, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, A.; Hare, D.; Bussey, T.; Saksida, L. Perineuronal Nets: Plasticity, Protection, and Therapeutic Potential. Trends Neurosci. 2019, 42, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Bitanihirwe, B.; Woo, T. Perineuronal nets and schizophrenia: The importance of neuronal coatings. Neurosci. Biobehav. Rev. 2014, 45, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Sorg, B.; Berretta, S.; Blacktop, J.; Fawcett, J.; Kitagawa, H.; Kwok, J.; Miquel, M. Casting a Wide Net: Role of Perineuronal Nets in Neural Plasticity. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 11459–11468. [Google Scholar] [CrossRef] [PubMed]
- Bosiacki, M.; Gąssowska-Dobrowolska, M.; Kojder, K.; Fabiańska, M.; Jeżewski, D.; Gutowska, I.; Lubkowska, A. Perineuronal Nets and Their Role in Synaptic Homeostasis. Int. J. Mol. Sci. 2019, 20, 4108. [Google Scholar] [CrossRef]
- Celio, M.; Spreafico, R.; De Biasi, S.; Vitellaro-Zuccarello, L. Perineuronal nets: Past and present. Trends Neurosci. 1998, 21, 510–515. [Google Scholar] [CrossRef]
- Li, X.; Li, N.; Zhao, P.; Ren, D.; Luo, B.; Zhou, T. Perineuronal Nets: From Structure to Neurological Disorders. Curr. Med. Chem. 2024, 31. [Google Scholar] [CrossRef]
- Yong, V.W.; Power, C.; Forsyth, P.; Edwards, D.R. Metalloproteinases in biology and pathology of the nervous system. Nat. Rev. Neurosci. 2001, 2, 502–511. [Google Scholar] [CrossRef]
- Hayes, A.; Melrose, J. Neural Tissue Homeostasis and Repair Is Regulated via CS and DS Proteoglycan Motifs. Front. Cell Dev. Biol. 2021, 9, 696640. [Google Scholar] [CrossRef]
- Carbo-Gas, M.; Moreno-Rius, J.; Guarque-Chabrera, J.; Vazquez-Sanroman, D.; Gil-Miravet, I.; Carulli, D.; Hoebeek, F.; De Zeeuw, C.; Sanchis-Segura, C.; Miquel, M. Cerebellar perineuronal nets in cocaine-induced pavlovian memory: Site matters. Neuropharmacology 2017, 125, 166–180. [Google Scholar] [CrossRef]
- Mörgelin, M.; Heinegård, D.; Engel, J.; Paulsson, M. The cartilage proteoglycan aggregate: Assembly through combined protein-carbohydrate and protein-protein interactions. Biophys. Chem. 1994, 50, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Galtrey, C.M.; Fawcett, J.W. The role of chondroitin sulfate proteoglycans in regeneration and plasticity in the central nervous system. Brain Res. Rev. 2007, 54, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Carulli, D.; Pizzorusso, T.; Kwok, J.; Putignano, E.; Poli, A.; Forostyak, S.; Andrews, M.; Deepa, S.; Glant, T.; Fawcett, J. Animals lacking link protein have attenuated perineuronal nets and persistent plasticity. Brain A J. Neurol. 2010, 133, 2331–2347. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, N.; Miyata, S.; Tamada, A.; Watanabe, Y.; Kawasaki, A.; Kitagawa, H.; Takao, K.; Miyakawa, T.; Takeuchi, K.; Igarashi, M. Abnormalities in perineuronal nets and behavior in mice lacking CSGalNAcT1, a key enzyme in chondroitin sulfate synthesis. Mol. Brain 2017, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Geissler, M.; Gottschling, C.; Aguado, A.; Rauch, U.; Wetzel, C.; Hatt, H.; Faissner, A. Primary hippocampal neurons, which lack four crucial extracellular matrix molecules, display abnormalities of synaptic structure and function and severe deficits in perineuronal net formation. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 7742–7755. [Google Scholar] [CrossRef] [PubMed]
- Carulli, D.; Rhodes, K.E.; Brown, D.J.; Bonnert, T.P.; Pollack, S.J.; Oliver, K.; Strata, P.; Fawcett, J.W. Composition of perineuronal nets in the adult rat cerebellum and the cellular origin of their components. J. Comp. Neurol. 2006, 494, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Oohashi, T.; Edamatsu, M.; Bekku, Y.; Carulli, D. The hyaluronan and proteoglycan link proteins: Organizers of the brain extracellular matrix and key molecules for neuronal function and plasticity. Exp. Neurol. 2015, 274, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Giamanco, K.A.; Matthews, R.T. Deconstructing the Perineuronal Net: Cellular Contributions and Molecular Composition of the Neuronal Extracellular Matrix. Neuroscience 2012, 218, 367–384. [Google Scholar] [CrossRef]
- Miyata, S.; Nadanaka, S.; Igarashi, M.; Kitagawa, H. Structural Variation of Chondroitin Sulfate Chains Contributes to the Molecular Heterogeneity of Perineuronal Nets. Front. Integr. Neurosci. 2018, 12, 3. [Google Scholar] [CrossRef]
- Duncan, J.; Foster, R.; Kwok, J. The potential of memory enhancement through modulation of perineuronal nets. Br. J. Pharmacol. 2019, 176, 3611–3621. [Google Scholar] [CrossRef]
- Mulligan, K.A.; Vanbrederode, J.F.M.; Hendrickson, A.E. The Lectin Vicia-Villosa Labels a Distinct Subset of Gabaergic Cells in Macaque Visual-Cortex. Vis. Neurosci 1989, 2, 63–72. [Google Scholar] [CrossRef]
- Giamanco, K.A.; Morawski, M.; Matthews, R.T. Perineuronal net formation and structure in aggrecan knockout mice. Neuroscience 2010, 170, 1314–1327. [Google Scholar] [CrossRef] [PubMed]
- Matthews, R.; Kelly, G.; Zerillo, C.; Gray, G.; Tiemeyer, M.; Hockfield, S. Aggrecan glycoforms contribute to the molecular heterogeneity of perineuronal nets. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 7536–7547. [Google Scholar] [CrossRef] [PubMed]
- Brückner, G.; Szeöke, S.; Pavlica, S.; Grosche, J.; Kacza, J. Axon initial segment ensheathed by extracellular matrix in perineuronal nets. Neuroscience 2006, 138, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Morris, N.; Henderson, Z. Perineuronal nets ensheath fast spiking, parvalbumin-immunoreactive neurons in the medial septum/diagonal band complex. Eur. J. Neurosci. 2000, 12, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Brückner, G.; Härtig, W.; Kacza, J.; Seeger, J.; Welt, K.; Brauer, K. Extracellular matrix organization in various regions of rat brain grey matter. J. Neurocytol. 1996, 25, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Wei, X.; Wang, X.; Du, S.; Liu, W.; Song, J.; Wang, Y. Perineuronal nets protect long-term memory by limiting activity-dependent inhibition from parvalbumin interneurons. Proc. Natl. Acad. Sci. USA 2019, 116, 27063–27073. [Google Scholar] [CrossRef] [PubMed]
- Morino-Wannier, P.; Fujita, S.; Jones, E. GABAergic neuronal populations in monkey primary auditory cortex defined by co-localized calcium binding proteins and surface antigens. Exp. Brain Res. 1992, 88, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.; Brauer, K.; Arélin, C.; Härtig, W.; Fine, A.; Mäder, M.; Arendt, T.; Brückner, G. Perineuronal nets in the rhesus monkey and human basal forebrain including basal ganglia. Neuroscience 2001, 108, 285–298. [Google Scholar] [CrossRef]
- Wegner, F.; Härtig, W.; Bringmann, A.; Grosche, J.; Wohlfarth, K.; Zuschratter, W.; Brückner, G. Diffuse perineuronal nets and modified pyramidal cells immunoreactive for glutamate and the GABA(A) receptor alpha1 subunit form a unique entity in rat cerebral cortex. Exp. Neurol. 2003, 184, 705–714. [Google Scholar] [CrossRef]
- Mészár, Z.; Girard, F.; Saper, C.; Celio, M. The lateral hypothalamic parvalbumin-immunoreactive (PV1) nucleus in rodents. J. Comp. Neurol. 2012, 520, 798–815. [Google Scholar] [CrossRef]
- Horii-Hayashi, N.; Sasagawa, T.; Hashimoto, T.; Kaneko, T.; Takeuchi, K.; Nishi, M. A newly identified mouse hypothalamic area having bidirectional neural connections with the lateral septum: The perifornical area of the anterior hypothalamus rich in chondroitin sulfate proteoglycans. Eur. J. Neurosci. 2015, 42, 2322–2334. [Google Scholar] [CrossRef]
- Balmer, T. Perineuronal Nets Enhance the Excitability of Fast-Spiking Neurons. eNeuro 2016, 3, 0112. [Google Scholar] [CrossRef]
- Yamada, J.; Ohgomori, T.; Jinno, S. Perineuronal nets affect parvalbumin expression in GABAergic neurons of the mouse hippocampus. Eur. J. Neurosci. 2015, 41, 368–378. [Google Scholar] [CrossRef]
- Carstens, K.; Phillips, M.; Pozzo-Miller, L.; Weinberg, R.; Dudek, S. Perineuronal Nets Suppress Plasticity of Excitatory Synapses on CA2 Pyramidal Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 6312–6320. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, B.B.; Madden, A.M.K.; Edwards, K.A.; Zup, S.L.; Stary, C.M. Age-dependent sexual dimorphism in hippocampal cornu ammonis-1 perineuronal net expression in rats. Brain Behav. 2019, 9, e01265. [Google Scholar] [CrossRef] [PubMed]
- Cornez, G.; Jonckers, E.; Ter Haar, S.; Van der Linden, A.; Cornil, C.; Balthazart, J. Timing of perineuronal net development in the zebra finch song control system correlates with developmental song learning. Proc. Biol. Sci. 2018, 285, 20180849. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, A.; Sugahara, K.; Faissner, A. Chondroitin sulfate “wobble motifs” modulate maintenance and differentiation of neural stem cells and their progeny. J. Biol. Chem. 2012, 287, 2935–2942. [Google Scholar] [CrossRef]
- Zhang, Z.; Dawson, P.; Piper, M.; Simmons, D. Postnatal N-acetylcysteine administration rescues impaired social behaviors and neurogenesis in Slc13a4 haploinsufficient mice. EBioMedicine 2019, 43, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Sharmin, S.; Pradhan, J.; Zhang, Z.; Bellingham, M.; Simmons, D.; Piper, M. Perineuronal net abnormalities in Slc13a4 mice are rescued by postnatal administration of N-acetylcysteine. Exp. Neurol. 2021, 342, 113734. [Google Scholar] [CrossRef] [PubMed]
- Levelt, C.; Hübener, M. Critical-period plasticity in the visual cortex. Annu. Rev. Neurosci. 2012, 35, 309–330. [Google Scholar] [CrossRef]
- Lensjø, K.; Lepperød, M.; Dick, G.; Hafting, T.; Fyhn, M. Removal of Perineuronal Nets Unlocks Juvenile Plasticity through Network Mechanisms of Decreased Inhibition and Increased Gamma Activity. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 1269–1283. [Google Scholar] [CrossRef]
- Sigal, Y.; Bae, H.; Bogart, L.; Hensch, T.; Zhuang, X. Structural maturation of cortical perineuronal nets and their perforating synapses revealed by superresolution imaging. Proc. Natl. Acad. Sci. USA 2019, 116, 7071–7076. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, K.P.; Figueroa Velez, D.X.; Habeeb, M.; Gandhi, S.P. Precocious deposition of perineuronal nets on Parvalbumin inhibitory neurons transplanted into adult visual cortex. Sci. Rep. 2018, 8, 7480. [Google Scholar] [CrossRef] [PubMed]
- Harauzov, A.; Spolidoro, M.; DiCristo, G.; De Pasquale, R.; Cancedda, L.; Pizzorusso, T.; Viegi, A.; Berardi, N.; Maffei, L. Reducing Intracortical Inhibition in the Adult Visual Cortex Promotes Ocular Dominance Plasticity. J. Neurosci. 2010, 30, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Franco, J.M.; Ouellet, L.; Kamal, B.; de Villers-Sidani, E. A Brain without Brakes: Reduced Inhibition Is Associated with Enhanced but Dysregulated Plasticity in the Aged Rat Auditory Cortex. Eneuro 2018, 5, 0051. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Franco, J.M.; de Villers-Sidani, É. Reactivation of critical period plasticity in adult auditory cortex through chemogenetic silencing of parvalbumin-positive interneurons. Proc. Natl. Acad. Sci. USA 2019, 116, 26329–26331. [Google Scholar] [CrossRef] [PubMed]
- Cope, E.; Zych, A.; Katchur, N.; Waters, R.; Laham, B.; Diethorn, E.; Park, C.; Meara, W.; Gould, E. Atypical perineuronal nets in the CA2 region interfere with social memory in a mouse model of social dysfunction. Mol. Psychiatry 2021, 27, 3520–3531. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, R.; Heine, M.; Perrais, D.; Seidenbecher, C.I.; Choquet, D.; Gundelfinger, E.D. Brain extracellular matrix affects AMPA receptor lateral mobility and short-term synaptic plasticity. Nat. Neurosci. 2009, 12, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.-H.; Brakebusch, C.; Matthies, H.; Oohashi, T.; Hirsch, E.; Moser, M.; Krug, M.; Seidenbecher, C.I.; Boeckers, T.M.; Rauch, U. Neurocan is dispensable for brain development. Mol. Cell. Biol. 2001, 21, 5970–5978. [Google Scholar] [CrossRef]
- Brakebusch, C.; Seidenbecher, C.I.; Asztely, F.; Rauch, U.; Matthies, H.; Meyer, H.; Krug, M.; Böckers, T.M.; Zhou, X.; Kreutz, M.R. Brevican-deficient mice display impaired hippocampal CA1 long-term potentiation but show no obvious deficits in learning and memory. Mol. Cell. Biol. 2002, 22, 7417–7427. [Google Scholar] [CrossRef]
- Bukalo, O.; Schachner, M.; Dityatev, A. Modification of extracellular matrix by enzymatic removal of chondroitin sulfate and by lack of tenascin-R differentially affects several forms of synaptic plasticity in the hippocampus. Neuroscience 2001, 104, 359–369. [Google Scholar] [CrossRef]
- Evers, M.R.; Salmen, B.; Bukalo, O.; Rollenhagen, A.; Bösl, M.R.; Morellini, F.; Bartsch, U.; Dityatev, A.; Schachner, M. Impairment of L-type Ca2+ channel-dependent forms of hippocampal synaptic plasticity in mice deficient in the extracellular matrix glycoprotein tenascin-C. J. Neurosci. 2002, 22, 7177–7194. [Google Scholar] [CrossRef]
- Lev-Ram, V.; Palida, S.; Deerinck, T.J.; Bushong, E.A.; Toyoma, B.H.; Perez, A.; McClatchy, D.B.; Prichard, D.R.; Savas, J.N.; Ellisman, M.H. Do perineuronal nets stabilize the engram of a synaptic circuit? bioRxiv 2023, bioRxiv:2023.2004.2009.536164. [Google Scholar]
- Cabungcal, J.; Steullet, P.; Morishita, H.; Kraftsik, R.; Cuenod, M.; Hensch, T.; Do, K. Perineuronal nets protect fast-spiking interneurons against oxidative stress. Proc. Natl. Acad. Sci. USA 2013, 110, 9130–9135. [Google Scholar] [CrossRef] [PubMed]
- Morawski, M.; Brückner, M.; Riederer, P.; Brückner, G.; Arendt, T. Perineuronal nets potentially protect against oxidative stress. Exp. Neurol. 2004, 188, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Suttkus, A.; Rohn, S.; Weigel, S.; Glöckner, P.; Arendt, T.; Morawski, M. Aggrecan, link protein and tenascin-R are essential components of the perineuronal net to protect neurons against iron-induced oxidative stress. Cell Death Dis. 2014, 5, e1119. [Google Scholar] [CrossRef] [PubMed]
- Dingess, P.; Harkness, J.; Slaker, M.; Zhang, Z.; Wulff, S.; Sorg, B.; Brown, T. Consumption of a High-Fat Diet Alters Perineuronal Nets in the Prefrontal Cortex. Neural Plast. 2018, 2018, 2108373. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, A.; Gibson, G.; Abbott, K.; Hare, D. A high-fat high-sugar diet in adolescent rats impairs social memory and alters chemical markers characteristic of atypical neuroplasticity and parvalbumin interneuron depletion in the medial prefrontal cortex. Food Funct. 2019, 10, 1985–1998. [Google Scholar] [CrossRef]
- Morikawa, S.; Ikegaya, Y.; Narita, M.; Tamura, H. Activation of perineuronal net-expressing excitatory neurons during associative memory encoding and retrieval. Sci. Rep. 2017, 7, 46024. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Cacquevel, M.; Saksida, L.; Bussey, T.; Schneider, B.; Aebischer, P.; Melani, R.; Pizzorusso, T.; Fawcett, J.; Spillantini, M. Perineuronal net digestion with chondroitinase restores memory in mice with tau pathology. Exp. Neurol. 2015, 265, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Okamoto, M.; Aoki, S.; Ishihara, T. Postnatal development of GABAergic interneurons and perineuronal nets in mouse temporal cortex subregions. Int. J. Dev. Neurosci. 2017, 63, 27–37. [Google Scholar] [CrossRef]
- Morishita, H.; Cabungcal, J.; Chen, Y.; Do, K.; Hensch, T. Prolonged Period of Cortical Plasticity upon Redox Dysregulation in Fast-Spiking Interneurons. Biol. Psychiatry 2015, 78, 396–402. [Google Scholar] [CrossRef]
- Tsien, R. Very long-term memories may be stored in the pattern of holes in the perineuronal net. Proc. Natl. Acad. Sci. USA 2013, 110, 12456–12461. [Google Scholar] [CrossRef]
- McRae, P.A.; Porter, B.E. The perineuronal net component of the extracellular matrix in plasticity and epilepsy. Neurochem. Int 2012, 61, 963–972. [Google Scholar] [CrossRef]
- Goode, T.; Tanaka, K.; Sahay, A.; McHugh, T. An Integrated Index: Engrams, Place Cells, and Hippocampal Memory. Neuron 2020, 107, 805–820. [Google Scholar] [CrossRef]
- Giorgi, C.; Marinelli, S. Roles and Transcriptional Responses of Inhibitory Neurons in Learning and Memory. Front. Mol. Neurosci. 2021, 14, 689952. [Google Scholar] [CrossRef]
- Morrison, D.; Rashid, A.; Yiu, A.; Yan, C.; Frankland, P.; Josselyn, S. Parvalbumin interneurons constrain the size of the lateral amygdala engram. Neurobiol. Learn. Mem. 2016, 135, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Barron, H.; Vogels, T.; Behrens, T.; Ramaswami, M. Inhibitory engrams in perception and memory. Proc. Natl. Acad. Sci. USA 2017, 114, 6666–6674. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Berretta, S. In Sickness and in Health: Perineuronal Nets and Synaptic Plasticity in Psychiatric Disorders. Neural Plast. 2016, 2016, 9847696. [Google Scholar] [CrossRef]
- Kwok, J.; Dick, G.; Wang, D.; Fawcett, J. Extracellular matrix and perineuronal nets in CNS repair. Dev. Neurobiol. 2011, 71, 1073–1089. [Google Scholar] [CrossRef]
- Favuzzi, E.; Marques-Smith, A.; Deogracias, R.; Winterflood, C.; Sánchez-Aguilera, A.; Mantoan, L.; Maeso, P.; Fernandes, C.; Ewers, H.; Rico, B. Activity-Dependent Gating of Parvalbumin Interneuron Function by the Perineuronal Net Protein Brevican. Neuron 2017, 95, 639–655.e610. [Google Scholar] [CrossRef]
- Dityatev, A.; Brückner, G.; Dityateva, G.; Grosche, J.; Kleene, R.; Schachner, M. Activity-dependent formation and functions of chondroitin sulfate-rich extracellular matrix of perineuronal nets. Dev. Neurobiol. 2007, 67, 570–588. [Google Scholar] [CrossRef]
- Donato, F.; Chowdhury, A.; Lahr, M.; Caroni, P. Early- and late-born parvalbumin basket cell subpopulations exhibiting distinct regulation and roles in learning. Neuron 2015, 85, 770–786. [Google Scholar] [CrossRef]
- Romberg, C.; Yang, S.; Melani, R.; Andrews, M.; Horner, A.; Spillantini, M.; Bussey, T.; Fawcett, J.; Pizzorusso, T.; Saksida, L. Depletion of perineuronal nets enhances recognition memory and long-term depression in the perirhinal cortex. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 7057–7065. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G. Hippocampal sharp wave-ripple: A cognitive biomarker for episodic memory and planning. Hippocampus 2015, 25, 1073–1188. [Google Scholar] [CrossRef]
- Sun, Z.; Bozzelli, P.; Caccavano, A.; Allen, M.; Balmuth, J.; Vicini, S.; Wu, J.; Conant, K. Disruption of perineuronal nets increases the frequency of sharp wave ripple events. Hippocampus 2018, 28, 42–52. [Google Scholar] [CrossRef]
- Donato, F.; Rompani, S.; Caroni, P. Parvalbumin-expressing basket-cell network plasticity induced by experience regulates adult learning. Nature 2013, 504, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Gogolla, N.; Caroni, P.; Lüthi, A.; Herry, C. Perineuronal nets protect fear memories from erasure. Science 2009, 325, 1258–1261. [Google Scholar] [CrossRef]
- Kawano, H.; Kimura-Kuroda, J.; Komuta, Y.; Yoshioka, N.; Li, H.; Kawamura, K.; Li, Y.; Raisman, G. Role of the lesion scar in the response to damage and repair of the central nervous system. Cell Tissue Res. 2012, 349, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Snow, D.; Lemmon, V.; Carrino, D.; Caplan, A.; Silver, J. Sulfated proteoglycans in astroglial barriers inhibit neurite outgrowth in vitro. Exp. Neurol. 1990, 109, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Laabs, T.; Wang, H.; Katagiri, Y.; McCann, T.; Fawcett, J.; Geller, H. Inhibiting glycosaminoglycan chain polymerization decreases the inhibitory activity of astrocyte-derived chondroitin sulfate proteoglycans. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 14494–14501. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Shuto, M.; Funakoshi, K. Chondroitin Sulfate Expression in Perineuronal Nets after Goldfish Spinal Cord Lesion. Front. Cell. Neurosci. 2018, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Starkey, M.; Bartus, K.; Barritt, A.; Bradbury, E. Chondroitinase ABC promotes compensatory sprouting of the intact corticospinal tract and recovery of forelimb function following unilateral pyramidotomy in adult mice. Eur. J. Neurosci. 2012, 36, 3665–3678. [Google Scholar] [CrossRef] [PubMed]
- Orlando, C.; Raineteau, O. Integrity of cortical perineuronal nets influences corticospinal tract plasticity after spinal cord injury. Brain Struct. Funct. 2015, 220, 1077–1091. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Iwanaga, H.; Murakami, T.; Abe, K. Three-dimensional microanatomy of perineuronal proteoglycan nets enveloping motor neurons in the rat spinal cord. J. Neurocytol. 1998, 27, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Arbat-Plana, A.; Torres-Espín, A.; Navarro, X.; Udina, E. Activity dependent therapies modulate the spinal changes that motoneurons suffer after a peripheral nerve injury. Exp. Neurol. 2015, 263, 293–305. [Google Scholar] [CrossRef]
- Leger, M.; Quiedeville, A.; Bouet, V.; Haelewyn, B.; Boulouard, M.; Schumann-Bard, P.; Freret, T. Object recognition test in mice. Nat. Protoc. 2013, 8, 2531–2537. [Google Scholar] [CrossRef]
- Polydoro, M.; Acker, C.; Duff, K.; Castillo, P.; Davies, P. Age-dependent impairment of cognitive and synaptic function in the htau mouse model of tau pathology. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 10741–10749. [Google Scholar] [CrossRef]
- Baidoe-Ansah, D.; Mirzapourdelavar, H.; Carceller, H.; Perez, M.; Strackeljan, L.; Garcia, B.; Seidenbecher, C.I.; Kaushik, R.; Nacher, J.; Dityatev, A. Depletion of neurocan in the prefrontal cortex impairs temporal order recognition, cognitive flexibility and perisomatic GABAergic innervation. bioRxiv 2023, bioRxiv:2023.2004.2018.537277. [Google Scholar]
- Dubisova, J.; Burianova, J.; Svobodova, L.; Makovicky, P.; Martinez-Varea, N.; Cimpean, A.; Fawcett, J.; Kwok, J.; Kubinova, S. Oral treatment of 4-methylumbelliferone reduced perineuronal nets and improved recognition memory in mice. Brain Res. Bull. 2022, 181, 144–156. [Google Scholar] [CrossRef]
- Rowlands, D.; Lensjø, K.; Dinh, T.; Yang, S.; Andrews, M.; Hafting, T.; Fyhn, M.; Fawcett, J.; Dick, G. Aggrecan Directs Extracellular Matrix-Mediated Neuronal Plasticity. J. Neurosci. Off. J. Soc. Neurosci. 2018, 38, 10102–10113. [Google Scholar] [CrossRef]
- Lesnikova, A.; Casarotto, P.; Moliner, R.; Fred, S.; Biojone, C.; Castrén, E. Perineuronal Net Receptor PTPσ Regulates Retention of Memories. Front. Synaptic Neurosci. 2021, 13, 672475. [Google Scholar] [CrossRef]
- Shen, H. Core Concept: Perineuronal nets gain prominence for their role in learning, memory, and plasticity. Proc. Natl. Acad. Sci. USA 2018, 115, 9813–9815. [Google Scholar] [CrossRef]
- Paylor, J.; Wendlandt, E.; Freeman, T.; Greba, Q.; Marks, W.; Howland, J.; Winship, I. Impaired Cognitive Function after Perineuronal Net Degradation in the Medial Prefrontal Cortex. eNeuro 2018, 5, 1–15. [Google Scholar] [CrossRef]
- Carulli, D.; Broersen, R.; de Winter, F.; Muir, E.; Mešković, M.; de Waal, M.; de Vries, S.; Boele, H.; Canto, C.; De Zeeuw, C.; et al. Cerebellar plasticity and associative memories are controlled by perineuronal nets. Proc. Natl. Acad. Sci. USA 2020, 117, 6855–6865. [Google Scholar] [CrossRef]
- Buzsáki, G.; Wang, X. Mechanisms of gamma oscillations. Annu. Rev. Neurosci. 2012, 35, 203–225. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Park, M.; Kang, J.; Lee, E.; Jung, J.; Kim, T. Aberrant Gamma-Band Oscillations in Mice with Vitamin D Deficiency: Implications on Schizophrenia and its Cognitive Symptoms. J. Pers. Med. 2022, 12, 318. [Google Scholar] [CrossRef] [PubMed]
- Riga, D.; Kramvis, I.; Koskinen, M.; van Bokhoven, P.; van der Harst, J.; Heistek, T.; Jaap Timmerman, A.; van Nierop, P.; van der Schors, R.; Pieneman, A.; et al. Hippocampal extracellular matrix alterations contribute to cognitive impairment associated with a chronic depressive-like state in rats. Sci. Transl. Med. 2017, 9, eaai8753. [Google Scholar] [CrossRef]
- Slaker, M.; Blacktop, J.M.; Sorg, B.A. Caught in the Net: Perineuronal Nets and Addiction. Neural Plast. 2016, 2016, 7538208. [Google Scholar] [CrossRef]
- Nabel, E.; Morishita, H. Regulating critical period plasticity: Insight from the visual system to fear circuitry for therapeutic interventions. Front. Psychiatry 2013, 4, 146. [Google Scholar] [CrossRef]
- Yang, W.; Liu, T.; Cao, J.; Chen, X.; Liu, X.; Wang, M.; Su, X.; Zhang, S.; Qiu, B.; Hu, W.; et al. Fear Erasure Facilitated by Immature Inhibitory Neuron Transplantation. Neuron 2016, 92, 1352–1367. [Google Scholar] [CrossRef]
- Fawcett, J.; Oohashi, T.; Pizzorusso, T. The roles of perineuronal nets and the perinodal extracellular matrix in neuronal function. Nat. Rev. Neurosci. 2019, 20, 451–465. [Google Scholar] [CrossRef]
- Reinhard, S.; Rais, M.; Afroz, S.; Hanania, Y.; Pendi, K.; Espinoza, K.; Rosenthal, R.; Binder, D.; Ethell, I.; Razak, K. Reduced perineuronal net expression in Fmr1 KO mice auditory cortex and amygdala is linked to impaired fear-associated memory. Neurobiol. Learn. Mem. 2019, 164, 107042. [Google Scholar] [CrossRef]
- Banerjee, S.; Gutzeit, V.; Baman, J.; Aoued, H.; Doshi, N.; Liu, R.; Ressler, K. Perineuronal Nets in the Adult Sensory Cortex Are Necessary for Fear Learning. Neuron 2017, 95, 169–179.e163. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.; Terstege, D.; Scott, G.; Tsutsui, M.; Epp, J. Neurogenesis mediated plasticity is associated with reduced neuronal activity in CA1 during context fear memory retrieval. Sci. Rep. 2022, 12, 7016. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.; Lensjø, K.; Wigestrand, M.; Malthe-Sørenssen, A.; Hafting, T.; Fyhn, M. Removal of perineuronal nets disrupts recall of a remote fear memory. Proc. Natl. Acad. Sci. USA 2018, 115, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Hylin, M.; Orsi, S.; Moore, A.; Dash, P. Disruption of the perineuronal net in the hippocampus or medial prefrontal cortex impairs fear conditioning. Learn. Mem. 2013, 20, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Kochlamazashvili, G.; Henneberger, C.; Bukalo, O.; Dvoretskova, E.; Senkov, O.; Lievens, P.; Westenbroek, R.; Engel, A.; Catterall, W.; Rusakov, D.; et al. The extracellular matrix molecule hyaluronic acid regulates hippocampal synaptic plasticity by modulating postsynaptic L-type Ca(2+) channels. Neuron 2010, 67, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, Y.; Men, S.; Li, X.; Hou, S.; Ju, J. Elimination of perineuronal nets in CA1 disrupts GABA release and long-term contextual fear memory retention. Hippocampus 2023, 33, 862–871. [Google Scholar] [CrossRef]
- Jovasevic, V.; Zhang, H.; Sananbenesi, F.; Guedea, A.; Soman, K.; Wiktorowicz, J.; Fischer, A.; Radulovic, J. Primary cilia are required for the persistence of memory and stabilization of perineuronal nets. iScience 2021, 24, 102617. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Gisabella, B.; Rexrode, L.; Benefield, D.; Yildiz, E.; Seltzer, P.; Valeri, J.; Chelini, G.; Reich, A.; Ardelt, M.; et al. Circadian Rhythms of Perineuronal Net Composition. eNeuro 2020, 7, 1–21. [Google Scholar] [CrossRef]
- Bjerknes, T.; Moser, E.; Moser, M. Representation of geometric borders in the developing rat. Neuron 2014, 82, 71–78. [Google Scholar] [CrossRef]
- Christensen, A.; Lensjø, K.; Lepperød, M.; Dragly, S.; Sutterud, H.; Blackstad, J.; Fyhn, M.; Hafting, T. Perineuronal nets stabilize the grid cell network. Nat. Commun. 2021, 12, 253. [Google Scholar] [CrossRef] [PubMed]
- Bertocchi, I.; Mele, P.; Ferrero, G.; Oberto, A.; Carulli, D.; Eva, C. NPY-Y1 receptor signaling controls spatial learning and perineuronal net expression. Neuropharmacology 2021, 184, 108425. [Google Scholar] [CrossRef] [PubMed]
- Tajerian, M.; Hung, V.; Nguyen, H.; Lee, G.; Joubert, L.; Malkovskiy, A.; Zou, B.; Xie, S.; Huang, T.; Clark, J. The hippocampal extracellular matrix regulates pain and memory after injury. Mol. Psychiatry 2018, 23, 2302–2313. [Google Scholar] [CrossRef] [PubMed]
- Fretham, S.; Carlson, E.; Wobken, J.; Tran, P.; Petryk, A.; Georgieff, M. Temporal manipulation of transferrin-receptor-1-dependent iron uptake identifies a sensitive period in mouse hippocampal neurodevelopment. Hippocampus 2012, 22, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Blacktop, J.; Todd, R.; Sorg, B. Role of perineuronal nets in the anterior dorsal lateral hypothalamic area in the acquisition of cocaine-induced conditioned place preference and self-administration. Neuropharmacology 2017, 118, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Lasek, A.; Chen, H.; Chen, W. Releasing Addiction Memories Trapped in Perineuronal Nets. Trends Genet. TIG 2018, 34, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Mash, D.; ffrench-Mullen, J.; Adi, N.; Qin, Y.; Buck, A.; Pablo, J. Gene expression in human hippocampus from cocaine abusers identifies genes which regulate extracellular matrix remodeling. PLoS ONE 2007, 2, e1187. [Google Scholar] [CrossRef]
- Smith, A.; Kupchik, Y.; Scofield, M.; Gipson, C.; Wiggins, A.; Thomas, C.; Kalivas, P. Synaptic plasticity mediating cocaine relapse requires matrix metalloproteinases. Nat. Neurosci. 2014, 17, 1655–1657. [Google Scholar] [CrossRef]
- Lee, B.; Dong, Y. Cocaine-induced metaplasticity in the nucleus accumbens: Silent synapse and beyond. Neuropharmacology 2011, 61, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, R.; Chang, K.; Rasband, M.; Seidenbecher, C. Neural ECM molecules in axonal and synaptic homeostatic plasticity. Prog. Brain Res. 2014, 214, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.; Kalivas, P.W.; Quirk, G.J. Extinction circuits for fear and addiction overlap in prefrontal cortex. Learn Mem. 2009, 16, 279–288. [Google Scholar] [CrossRef]
- Melchor-Eixea, I.; Guarque-Chabrera, J.; Sanchez-Hernandez, A.; Ibáñez-Marín, P.; Pastor, R.; Miquel, M. Putting forward a model for the role of the cerebellum in cocaine-induced pavlovian memory. Front. Syst. Neurosci. 2023, 17, 1154014. [Google Scholar] [CrossRef] [PubMed]
- Guarque-Chabrera, J.; Gil-Miravet, I.; Olucha-Bordonau, F.; Melchor-Eixea, I.; Miquel, M. When the front fails, the rear wins. Cerebellar correlates of prefrontal dysfunction in cocaine-induced memory in male rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 112, 110429. [Google Scholar] [CrossRef]
- Sorg, B.A.; Wingert, J.C.; Ramos, J.D.; Reynolds, S.X.; Gonzalez, A.E.; Rose, R.M.; Hegarty, D.M.; Aicher, S.A.; Bailey, L.G.; Brown, T.E. Perineuronal nets in the rat medial prefrontal cortex alter hippocampal-prefrontal oscillations and reshape cocaine self-administration memories. bioRxiv 2024, bioRxiv:2024.2002.2005.577568. [Google Scholar]
- Slaker, M.; Churchill, L.; Todd, R.; Blacktop, J.; Zuloaga, D.; Raber, J.; Darling, R.; Brown, T.; Sorg, B. Removal of perineuronal nets in the medial prefrontal cortex impairs the acquisition and reconsolidation of a cocaine-induced conditioned place preference memory. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 4190–4202. [Google Scholar] [CrossRef]
- Jorgensen, E.; Gonzalez, A.; Harkness, J.; Hegarty, D.; Thakar, A.; Burchi, D.; Aadland, J.; Aicher, S.; Sorg, B.; Brown, T. Cocaine memory reactivation induces functional adaptations within parvalbumin interneurons in the rat medial prefrontal cortex. Addict. Biol. 2021, 26, e12947. [Google Scholar] [CrossRef]
- Xue, Y.; Xue, L.; Liu, J.; He, J.; Deng, J.; Sun, S.; Han, H.; Luo, Y.; Xu, L.; Wu, P.; et al. Depletion of perineuronal nets in the amygdala to enhance the erasure of drug memories. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 6647–6658. [Google Scholar] [CrossRef]
- Valeri, J.; Stiplosek, C.; O’Donovan, S.M.; Sinclair, D.; Grant, K.A.; Bollavarapu, R.; Platt, D.M.; Stockmeier, C.A.; Gisabella, B.; Pantazopoulos, H. Extracellular matrix abnormalities in the hippocampus of subjects with substance use disorder. Transl. Psychiatry 2024, 14, 115. [Google Scholar] [CrossRef]
- Gray, D.T.; Khattab, S.; Meltzer, J.; McDermott, K.; Schwyhart, R.; Sinakevitch, I.; Härtig, W.; Barnes, C.A. Retrosplenial cortex microglia and perineuronal net densities are associated with memory impairment in aged rhesus macaques. Cereb. Cortex 2023, 33, 4626–4644. [Google Scholar] [CrossRef]
- Erickson, C.A.; Barnes, C.A. The neurobiology of memory changes in normal aging. Exp Gerontol 2003, 38, 61–69. [Google Scholar] [CrossRef]
- Richard, A.; Tian, X.; El-Saadi, M.; Lu, X. Erasure of striatal chondroitin sulfate proteoglycan-associated extracellular matrix rescues aging-dependent decline of motor learning. Neurobiol. Aging 2018, 71, 61–71. [Google Scholar] [CrossRef]
- Brückner, G.; Hausen, D.; Härtig, W.; Drlicek, M.; Arendt, T.; Brauer, K. Cortical areas abundant in extracellular matrix chondroitin sulphate proteoglycans are less affected by cytoskeletal changes in Alzheimer’s disease. Neuroscience 1999, 92, 791–805. [Google Scholar] [CrossRef]
- Morawski, M.; Brückner, G.; Jäger, C.; Seeger, G.; Matthews, R.; Arendt, T. Involvement of perineuronal and perisynaptic extracellular matrix in Alzheimer’s disease neuropathology. Brain Pathol. 2012, 22, 547–561. [Google Scholar] [CrossRef]
- Baig, S.; Wilcock, G.; Love, S. Loss of perineuronal net N-acetylgalactosamine in Alzheimer’s disease. Acta Neuropathol. 2005, 110, 393–401. [Google Scholar] [CrossRef]
- Kobayashi, K.; Emson, P.; Mountjoy, C. Vicia villosa lectin-positive neurones in human cerebral cortex. Loss in Alzheimer-type dementia. Brain Res. 1989, 498, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, A. Is loss of perineuronal nets a critical pathological event in Alzheimer’s disease? EBioMedicine 2020, 59, 102946. [Google Scholar] [CrossRef] [PubMed]
- Alonge, K.; Logsdon, A.; Murphree, T.; Banks, W.; Keene, C.; Edgar, J.; Whittington, D.; Schwartz, M.; Guttman, M. Quantitative analysis of chondroitin sulfate disaccharides from human and rodent fixed brain tissue by electrospray ionization-tandem mass spectrometry. Glycobiology 2019, 29, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Alonge, K.; Mirzadeh, Z.; Scarlett, J.; Logsdon, A.; Brown, J.; Cabrales, E.; Chan, C.; Kaiyala, K.; Bentsen, M.; Banks, W.; et al. Hypothalamic perineuronal net assembly is required for sustained diabetes remission induced by fibroblast growth factor 1 in rats. Nat. Metab. 2020, 2, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Härtig, W.; Klein, C.; Brauer, K.; Schüppel, K.; Arendt, T.; Bigl, V.; Brückner, G. Hyperphosphorylated protein tau is restricted to neurons devoid of perineuronal nets in the cortex of aged bison. Neurobiol. Aging 2001, 22, 25–33. [Google Scholar] [CrossRef]
- Díaz-Nido, J.; Wandosell, F.; Avila, J. Glycosaminoglycans and beta-amyloid, prion and tau peptides in neurodegenerative diseases. Peptides 2002, 23, 1323–1332. [Google Scholar] [CrossRef]
- Cattaud, V.; Bezzina, C.; Rey, C.; Lejards, C.; Dahan, L.; Verret, L. Early disruption of parvalbumin expression and perineuronal nets in the hippocampus of the Tg2576 mouse model of Alzheimer’s disease can be rescued by enriched environment. Neurobiol. Aging 2018, 72, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Howell, M.; Bailey, L.; Cozart, M.; Gannon, B.; Gottschall, P. Hippocampal administration of chondroitinase ABC increases plaque-adjacent synaptic marker and diminishes amyloid burden in aged APPswe/PS1dE9 mice. Acta Neuropathol. Commun. 2015, 3, 54. [Google Scholar] [CrossRef] [PubMed]
- Foscarin, S.; Raha-Chowdhury, R.; Fawcett, J.; Kwok, J. Brain ageing changes proteoglycan sulfation, rendering perineuronal nets more inhibitory. Aging 2017, 9, 1607–1622. [Google Scholar] [CrossRef]
- Miyata, S.; Komatsu, Y.; Yoshimura, Y.; Taya, C.; Kitagawa, H. Persistent cortical plasticity by upregulation of chondroitin 6-sulfation. Nat. Neurosci. 2012, 15, 414–422. [Google Scholar] [CrossRef]
- Yang, S.; Hilton, S.; Alves, J.; Saksida, L.; Bussey, T.; Matthews, R.; Kitagawa, H.; Spillantini, M.; Kwok, J.; Fawcett, J. Antibody recognizing 4-sulfated chondroitin sulfate proteoglycans restores memory in tauopathy-induced neurodegeneration. Neurobiol. Aging 2017, 59, 197–209. [Google Scholar] [CrossRef]
- Yang, S.; Gigout, S.; Molinaro, A.; Naito-Matsui, Y.; Hilton, S.; Foscarin, S.; Nieuwenhuis, B.; Tan, C.; Verhaagen, J.; Pizzorusso, T.; et al. Chondroitin 6-sulphate is required for neuroplasticity and memory in ageing. Mol. Psychiatry 2021, 26, 5658–5668. [Google Scholar] [CrossRef]
- Curley, A.A.; Lewis, D.A. Cortical basket cell dysfunction in schizophrenia. J. Physiol. 2012, 590, 715–724. [Google Scholar] [CrossRef]
- Glausier, J.; Fish, K.; Lewis, D. Altered parvalbumin basket cell inputs in the dorsolateral prefrontal cortex of schizophrenia subjects. Mol. Psychiatry 2014, 19, 30–36. [Google Scholar] [CrossRef]
- Enwright, J.F.; Sanapala, S.; Foglio, A.; Berry, R.; Fish, K.N.; Lewis, D.A. Reduced labeling of parvalbumin neurons and perineuronal nets in the dorsolateral prefrontal cortex of subjects with schizophrenia. Neuropsychopharmacology 2016, 41, 2206–2214. [Google Scholar] [CrossRef]
- Dityatev, A.; Seidenbecher, C.; Morawski, M. Brain extracellular matrix: An upcoming target in neurological and psychiatric disorders. Eur. J. Neurosci. 2021, 53, 3807–3810. [Google Scholar] [CrossRef]
- van ‘t Spijker, H.; Kwok, J. A Sweet Talk: The Molecular Systems of Perineuronal Nets in Controlling Neuronal Communication. Front. Integr. Neurosci. 2017, 11, 33. [Google Scholar] [CrossRef]
- Chelyshev, Y.; Kabdesh, I.; Mukhamedshina, Y. Extracellular Matrix in Neural Plasticity and Regeneration. Cell. Mol. Neurobiol. 2022, 42, 647–664. [Google Scholar] [CrossRef] [PubMed]
- Maleski, M.; Hockfield, S. Glial cells assemble hyaluronan-based pericellular matrices in vitro. Glia 1997, 20, 193–202. [Google Scholar] [CrossRef]
- Hayashi, N.; Mizusaki, M.; Kamei, K.; Harada, S.; Miyata, S. Chondroitin sulfate proteoglycan phosphacan associates with parallel fibers and modulates axonal extension and fasciculation of cerebellar granule cells. Mol. Cell. Neurosci. 2005, 30, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, V.; Benbenishty, A.; Blinder, P.; Sagi, I. Demystifying the extracellular matrix and its proteolytic remodeling in the brain: Structural and functional insights. Cell. Mol. Life Sci. CMLS 2019, 76, 3229–3248. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wu, X.; Lu, T.; Kuang, C.; Si, Y.; Zheng, W.; Li, Z.; Xue, Y. Perineuronal Nets in the CNS: Architects of Memory and Potential Therapeutic Target in Neuropsychiatric Disorders. Int. J. Mol. Sci. 2024, 25, 3412. https://doi.org/10.3390/ijms25063412
Li X, Wu X, Lu T, Kuang C, Si Y, Zheng W, Li Z, Xue Y. Perineuronal Nets in the CNS: Architects of Memory and Potential Therapeutic Target in Neuropsychiatric Disorders. International Journal of Molecular Sciences. 2024; 25(6):3412. https://doi.org/10.3390/ijms25063412
Chicago/Turabian StyleLi, Xue, Xianwen Wu, Tangsheng Lu, Chenyan Kuang, Yue Si, Wei Zheng, Zhonghao Li, and Yanxue Xue. 2024. "Perineuronal Nets in the CNS: Architects of Memory and Potential Therapeutic Target in Neuropsychiatric Disorders" International Journal of Molecular Sciences 25, no. 6: 3412. https://doi.org/10.3390/ijms25063412
APA StyleLi, X., Wu, X., Lu, T., Kuang, C., Si, Y., Zheng, W., Li, Z., & Xue, Y. (2024). Perineuronal Nets in the CNS: Architects of Memory and Potential Therapeutic Target in Neuropsychiatric Disorders. International Journal of Molecular Sciences, 25(6), 3412. https://doi.org/10.3390/ijms25063412