
CgNis1’s Impact on Virulence and Stress Response in Colletotrichum gloeosporioides

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Identification and Knockout of CgNIS1
2.2. CgNIS1 Deletion Affects Vegetative Growth
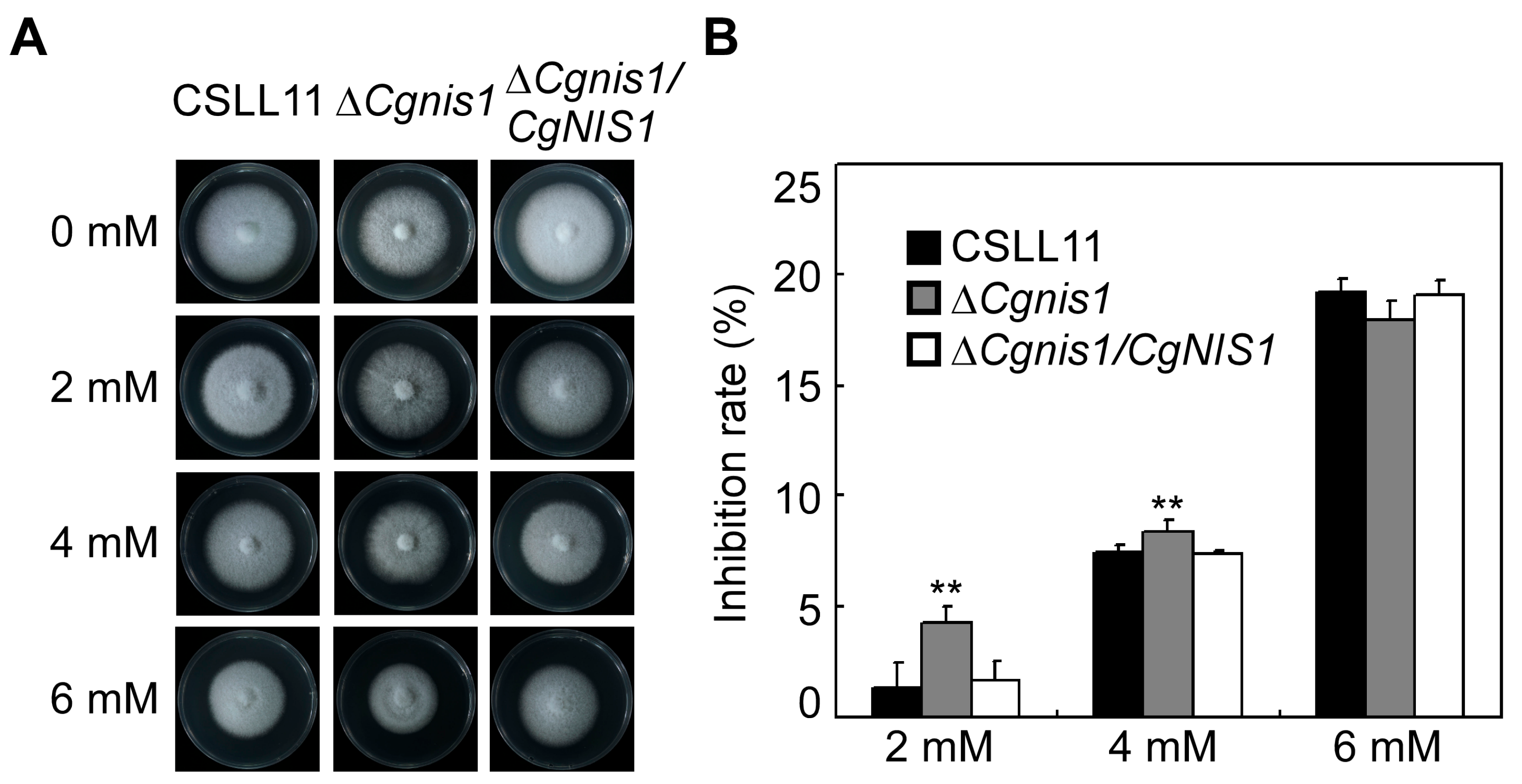
2.3. CgNis1 Is Important in H2O2 Responses
2.4. CgNIS1 Deletion Affects Cell Membrane Permeability
2.5. CgNis1 Is Involved in Conidiogenesis but Not Appressorium Formation
2.6. CgNis1 Is Required for Full Virulence
2.7. CgNis1 Induces Cell Necrosis
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. CgNIS1 Gene Disruption and ∆Cgnis1 Mutant Complementation
4.3. Southern Blotting
4.4. Vegetative Growth, Stress Response, and Cell Membrane Permeability
4.5. Conidiation and Appressorium Formation
4.6. Pathogenicity Analysis of the Gene Knockout Mutants
4.7. CgNis1 Induces Cell Necrosis
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ridzuan, R.; Rafii, M.Y.; Ismail, S.I.; Mohammad Yusoff, M.; Miah, G.; Usman, M. Breeding for Anthracnose Disease Resistance in Chili: Progress and Prospects. Int. J. Mol. Sci. 2018, 19, 3122. [Google Scholar] [CrossRef]
- Chen, Y.; Zeng, Q.; Man, Y.; Liu, S.; Ouyang, C.; Li, C.; Wu, X.; Zhang, D.; Liu, Y.; Tan, X. Simple sequence repeat markers reflect the biological phenotype differentiation and genetic diversity of Colletotrichum gloeosporioides strains from Capsicum annuum L. in China. J. Phytopathol. 2021, 169, 701–709. [Google Scholar] [CrossRef]
- Naveen, J.; Navya, H.M.; Hithamani, G.; Hariprasad, P.; Niranjana, S.R. Pathological, biochemical and molecular variability of Colletotrichum truncatum incitant of anthracnose disease in chilli (Capsicum annuum L.). Microb. Pathog. 2021, 152, 104611. [Google Scholar] [CrossRef]
- Ali, M.; Li, Q.H.; Zou, T.; Wei, A.M.; Gombojab, G.; Lu, G.; Gong, Z.H. Chitinase Gene Positively Regulates Hypersensitive and Defense Responses of Pepper to Colletotrichum acutatum Infection. Int. J. Mol. Sci. 2020, 21, 6624. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Do, J.W.; Yoon, J.B. Development of STS markers linked to the major QTLs for resistance to the pepper anthracnose caused by Colletotrichum acutatum and C. capsici. Hortic. Environ. Biotechnol. 2011, 52, 596–601. [Google Scholar] [CrossRef]
- Villafana, R.T.; Ramdass, A.C.; Rampersad, S.N. Colletotrichum brevisporum is associated with anthracnose of red bell pepper fruit in Trinidad. New Dis. Rep. 2019, 39, 11. [Google Scholar] [CrossRef]
- Than, P.P.; Prihastuti, H.; Phoulivong, S.; Taylor, P.W.; Hyde, K.D. Chilli anthracnose disease caused by Colletotrichum species. J. Zhejiang Univ. Sci. B 2008, 9, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Diao, Y.Z.; Zhang, C.; Liu, F.; Wang, W.Z.; Liu, L.; Cai, L.; Liu, X.L. Colletotrichum species causing anthracnose disease of chili in China. Persoonia 2017, 38, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Li, D.; Kolattukudy, P.E. Induction of Ca2+-calmodulin signaling by hard-surface contact primes Colletotrichum gloeosporioides conidia to germinate and form appressoria. J. Bacteriol. 1998, 180, 5144–5150. [Google Scholar] [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Fang, Y.L.; Xia, L.M.; Wang, P.; Zhu, L.H.; Ye, J.R.; Huang, L. The MAPKKK CgMck1 Is Required for Cell Wall Integrity, Appressorium Development, and Pathogenicity in Colletotrichum gloeosporioides. Genes 2018, 9, 543. [Google Scholar] [CrossRef]
- Priyatno, T.P.; Abu Bakar, F.D.; Kamaruddin, N.; Mahadi, N.M.; Abdul Murad, A.M. Inactivation of the catalytic subunit of cAMP-dependent protein kinase A causes delayed appressorium formation and reduced pathogenicity of Colletotrichum gloeosporioides. Sci. World J. 2012, 2012, 545784. [Google Scholar] [CrossRef]
- Barhoom, S.; Kupiec, M.; Zhao, X.; Xu, J.R.; Sharon, A. Functional characterization of CgCTR2, a putative vacuole copper transporter that is involved in germination and pathogenicity in Colletotrichum gloeosporioides. Eukaryot. Cell 2008, 7, 1098–1108. [Google Scholar] [CrossRef]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.L.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef] [PubMed]
- Gan, P.; Ikeda, K.; Irieda, H.; Narusaka, M.; O’Connell, R.J.; Narusaka, Y.; Takano, Y.; Kubo, Y.; Shirasu, K. Comparative genomic and transcriptomic analyses reveal the hemibiotrophic stage shift of Colletotrichum fungi. New Phytol. 2013, 197, 1236–1249. [Google Scholar] [CrossRef]
- Martin, F.; Aerts, A.; Ahrén, D.; Brun, A.; Danchin, E.G.; Duchaussoy, F.; Gibon, J.; Kohler, A.; Lindquist, E.; Pereda, V.; et al. The genome of Laccaria bicolor provides insights into mycorrhizal symbiosis. Nature 2008, 452, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Kohler, A.; Murat, C.; Balestrini, R.; Coutinho, P.M.; Jaillon, O.; Montanini, B.; Morin, E.; Noel, B.; Percudani, R.; et al. Périgord black truffle genome uncovers evolutionary origins and mechanisms of symbiosis. Nature 2010, 464, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Collemare, J.; Mehrabi, R.; De Wit, P.J. Phytotoxic secondary metabolites and peptides produced by plant pathogenic Dothideomycete fungi. FEMS Microbiol. Rev. 2013, 37, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Zuccaro, A.; Lahrmann, U.; Güldener, U.; Langen, G.; Pfiffi, S.; Biedenkopf, D.; Wong, P.; Samans, B.; Grimm, C.; Basiewicz, M.; et al. Endophytic life strategies decoded by genome and transcriptome analyses of the mutualistic root symbiont Piriformospora indica. PLoS Pathog. 2011, 7, e1002290. [Google Scholar] [CrossRef]
- Djamei, A.; Schipper, K.; Rabe, F.; Ghosh, A.; Vincon, V.; Kahnt, J.; Osorio, S.; Tohge, T.; Fernie, A.R.; Feussner, I.; et al. Metabolic priming by a secreted fungal effector. Nature 2011, 478, 395–398. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Spanu, P.D.; Abbott, J.C.; Amselem, J.; Burgis, T.A.; Soanes, D.M.; Stüber, K.; Ver Loren van Themaat, E.; Brown, J.K.; Butcher, S.A.; Gurr, S.J.; et al. Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism. Science 2010, 330, 1543–1546. [Google Scholar] [CrossRef] [PubMed]
- Wicker, T.; Oberhaensli, S.; Parlange, F.; Buchmann, J.P.; Shatalina, M.; Roffler, S.; Ben-David, R.; Doležel, J.; Šimková, H.; Schulze-Lefert, P.; et al. The wheat powdery mildew genome shows the unique evolution of an obligate biotroph. Nat. Genet. 2013, 45, 1092–1096. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Zhao, L.; Liao, Z.; Feng, L.; An, B.; He, C.; Wang, Q.; Luo, H. Colletotrichum gloeosporioides Cg2LysM contributed to virulence toward rubber tree through affecting invasive structure and inhibiting chitin-triggered plant immunity. Front. Microbiol. 2023, 14, 1129101. [Google Scholar] [CrossRef]
- Irieda, H.; Inoue, Y.; Mori, M.; Yamada, K.; Oshikawa, Y.; Saitoh, H.; Uemura, A.; Terauchi, R.; Kitakura, S.; Kosaka, A.; et al. Conserved fungal effector suppresses PAMP-triggered immunity by targeting plant immune kinases. Proc. Natl. Acad. Sci. USA 2019, 116, 496–505. [Google Scholar] [CrossRef]
- Yoshino, K.; Irieda, H.; Sugimoto, F.; Yoshioka, H.; Okuno, T.; Takano, Y. Cell death of Nicotiana benthamiana is induced by secreted protein NIS1 of Colletotrichum orbiculare and is suppressed by a homologue of CgDN3. Mol. Plant Microbe Interact. 2012, 25, 625–636. [Google Scholar] [CrossRef]
- Nie, J.; Zhou, W.; Lin, Y.; Liu, Z.; Yin, Z.; Huang, L. Two NIS1-like proteins from apple canker pathogen (Valsa mali) play distinct roles in plant recognition and pathogen virulence. Stress Biol. 2022, 2, 7. [Google Scholar] [CrossRef]
- Farkas, A.; Maróti, G.; Durgő, H.; Györgypál, Z.; Lima, R.M.; Medzihradszky, K.F.; Kereszt, A.; Mergaert, P.; Kondorosi, É. Medicago truncatula symbiotic peptide NCR247 contributes to bacteroid differentiation through multiple mechanisms. Proc. Natl. Acad. Sci. USA 2014, 111, 5183–5188. [Google Scholar] [CrossRef]
- Levin, E.; Ballester, A.R.; Raphael, G.; Feigenberg, O.; Liu, Y.; Norelli, J.; Gonzalez-Candelas, L.; Ma, J.; Dardick, C.; Wisniewski, M.; et al. Identification and characterization of LysM effectors in Penicillium expansum. PLoS ONE 2017, 12, e0186023. [Google Scholar] [CrossRef]
- Monaghan, J.; Zipfel, C. Plant pattern recognition receptor complexes at the plasma membrane. Curr. Opin. Plant Biol. 2012, 15, 349–357. [Google Scholar] [CrossRef]
- Li, L.; Li, M.; Yu, L.; Zhou, Z.; Liang, X.; Liu, Z.; Cai, G.; Gao, L.; Zhang, X.; Wang, Y.; et al. The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity. Cell Host Microbe 2014, 15, 329–338. [Google Scholar] [CrossRef]
- Takano, Y.; Komeda, K.; Kojima, K.; Okuno, T. Proper regulation of cyclic AMP-dependent protein kinase is required for growth, conidiation, and appressorium function in the anthracnose fungus Colletotrichum lagenarium. Mol. Plant Microbe Interact. 2001, 14, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhou, Y.; Li, H.; Bao, Y.; Duan, Z.; Wang, C.; Powell, C.A.; Wang, K.; Hu, Q.; Chen, B.; et al. Identification of common fungal extracellular membrane (CFEM) proteins in Fusarium sacchari that inhibit plant immunity and contribute to virulence. Microbiol. Spectr. 2023, 11, e0145223. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Magne, M.; Chen, S.; Côté, O.; Stare, B.G.; Obradovic, N.; Jamshaid, L.; Wang, X.; Bélair, G.; Moffett, P. Analysis of putative apoplastic effectors from the nematode, Globodera rostochiensis, and identification of an expansin-like protein that can induce and suppress host defenses. PLoS ONE 2015, 10, e0115042. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, W.; Xiang, T.; Liu, Z.; Laluk, K.; Ding, X.; Zou, Y.; Gao, M.; Zhang, X.; Chen, S.; et al. Receptor-like cytoplasmic kinases integrate signaling from multiple plant immune receptors and are targeted by a Pseudomonas syringae effector. Cell Host Microbe 2010, 7, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; He, P.; Li, J.; Heese, A.; Peck, S.C.; Nürnberger, T.; Martin, G.B.; Sheen, J. Bacterial effectors target the common signaling partner BAK1 to disrupt multiple MAMP receptor-signaling complexes and impede plant immunity. Cell Host Microbe 2008, 4, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wu, S.; Chen, X.; Liu, C.; Sheen, J.; Shan, L.; He, P. The Pseudomonas syringae effector HopF2 suppresses Arabidopsis immunity by targeting BAK1. Plant J. 2014, 77, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Yang, F.; Rong, W.; Wu, X.; Zhang, J.; Chen, S.; He, C.; Zhou, J.M. A Xanthomonas uridine 5′-monophosphate transferase inhibits plant immune kinases. Nature 2012, 485, 114–118. [Google Scholar] [CrossRef]
- Roux, M.; Schwessinger, B.; Albrecht, C.; Chinchilla, D.; Jones, A.; Holton, N.; Malinovsky, F.G.; Tör, M.; de Vries, S.; Zipfel, C. The Arabidopsis leucine-rich repeat receptor-like kinases BAK1/SERK3 and BKK1/SERK4 are required for innate immunity to hemibiotrophic and biotrophic pathogens. Plant Cell 2011, 23, 2440–2455. [Google Scholar] [CrossRef]
- Sweigard, J.A.; Chumley, F.G.; Valent, B. Disruption of a Magnaporthe grisea cutinase gene. Mol. Gen. Genet. 1992, 232, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xue, C.; Kim, Y.; Xu, J.-R. A Ligation-PCR Approach for Generating Gene Replacement Constructs in Magnaporthe grisea. Fungal Genet. Rep. 2004, 51, 17–18. [Google Scholar] [CrossRef]
- Sambrook, J.F.; Fritsch, E.F.; Maniatis, T. (Eds.) Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Zhang, H.; Zhao, Q.; Guo, X.; Guo, M.; Qi, Z.; Tang, W.; Dong, Y.; Ye, W.; Zheng, X.; Wang, P.; et al. Pleiotropic function of the putative zinc-finger protein MoMsn2 in Magnaporthe oryzae. Mol. Plant Microbe Interact. 2014, 27, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, X.; Gai, W.; Xiao, L.; Gong, Z. First Report of Colletotrichum gloeosporioides Causing Anthracnose on Pepper in Shaanxi Province, China. Plant Dis. 2021, 105, 2242. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.W.; Malik, A.; Rehman, A.; Hameed, A.; Sarwar, M.; Hussain, M.; Hussain, D.; Shafeeq, T. First Record of Colletotrichum gloeosporioides Causing Anthracnose of Banana in Pakistan. Plant Dis. 2021, 105, 2013. [Google Scholar] [CrossRef]
- Oliveira, M.S.; Wang, N.Y.; Peres, N.A. Multilocus Phylogenetic Analyses of Colletotrichum gloeosporioides Species Complex Causing Crown Rot on Strawberry in Florida. Phytopathology 2022, 112, 898–906. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, B.; Zhou, Y.; Yin, H.; An, B.; Lin, D.; He, C.; Luo, H. CgNPG1 as a Novel Pathogenic Gene of Colletotrichum gloeosporioides from Hevea brasiliensis in Mycelial Growth, Conidiation, and the Invasive Structures Development. Front. Microbiol. 2021, 12, 629387. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, S.; Sun, Q.; Liu, S.; Wu, F.; Li, C.; Zhang, X.; OuYang, C.; Chen, Y.; Tan, X. CgNis1’s Impact on Virulence and Stress Response in Colletotrichum gloeosporioides. Int. J. Mol. Sci. 2024, 25, 3505. https://doi.org/10.3390/ijms25063505
Guo S, Sun Q, Liu S, Wu F, Li C, Zhang X, OuYang C, Chen Y, Tan X. CgNis1’s Impact on Virulence and Stress Response in Colletotrichum gloeosporioides. International Journal of Molecular Sciences. 2024; 25(6):3505. https://doi.org/10.3390/ijms25063505
Chicago/Turabian StyleGuo, Sheng, Qianlong Sun, Sizhen Liu, Fei Wu, Chenggang Li, Xin Zhang, Chao OuYang, Yue Chen, and Xinqiu Tan. 2024. "CgNis1’s Impact on Virulence and Stress Response in Colletotrichum gloeosporioides" International Journal of Molecular Sciences 25, no. 6: 3505. https://doi.org/10.3390/ijms25063505
APA StyleGuo, S., Sun, Q., Liu, S., Wu, F., Li, C., Zhang, X., OuYang, C., Chen, Y., & Tan, X. (2024). CgNis1’s Impact on Virulence and Stress Response in Colletotrichum gloeosporioides. International Journal of Molecular Sciences, 25(6), 3505. https://doi.org/10.3390/ijms25063505