


Recombinant Arginine Deiminase from Levilactobacillus brevis Inhibits the Growth of Stomach Cancer Cells, Possibly by Activating the Intrinsic Apoptosis Pathway
 , , and
, , and
Abstract
:1. Introduction
2. Results
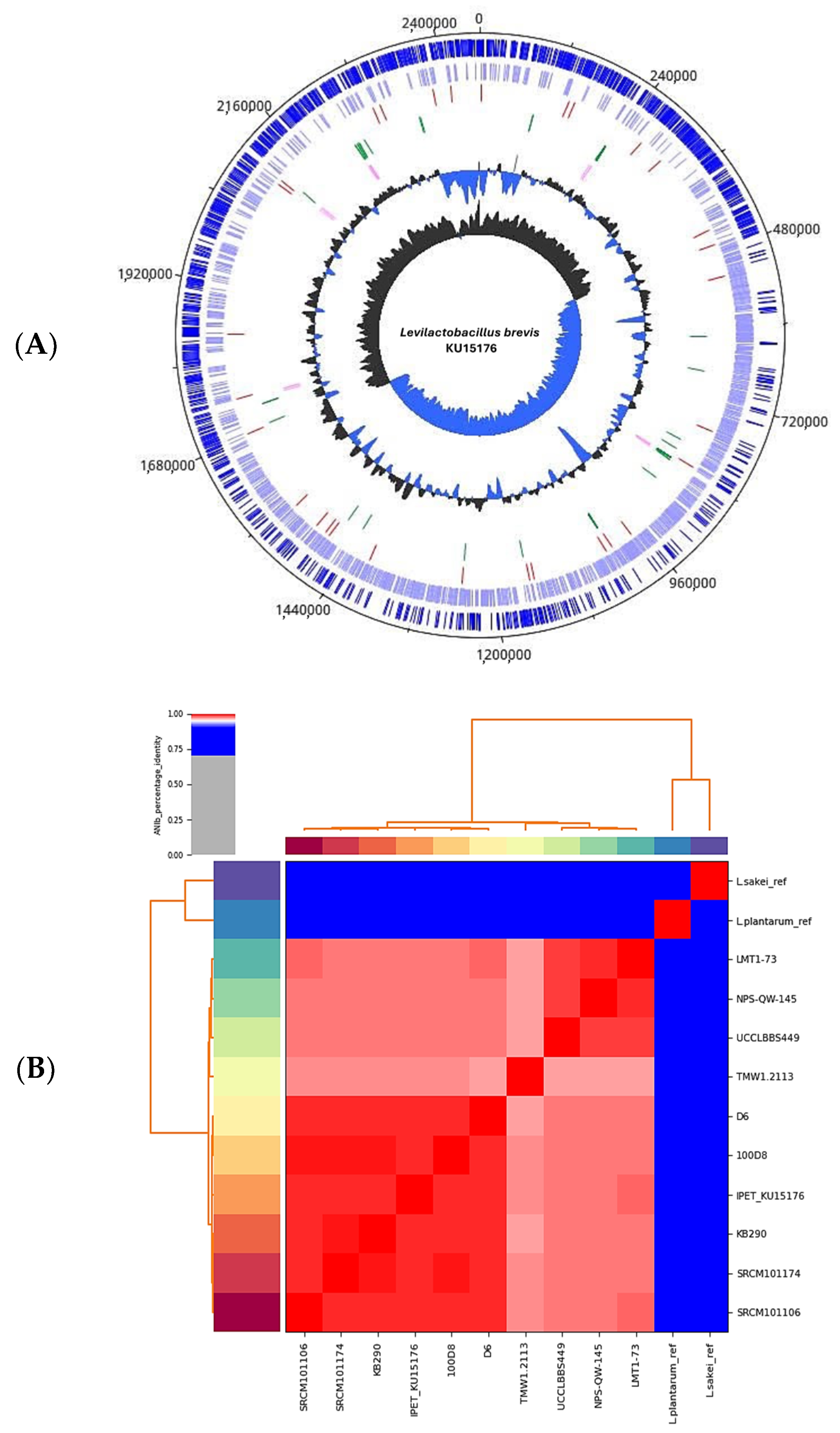
2.1. Genome Analysis, Identity Verification, and Identification of Potential Anticancer Genes and Gene Products in L. brevis KU15176
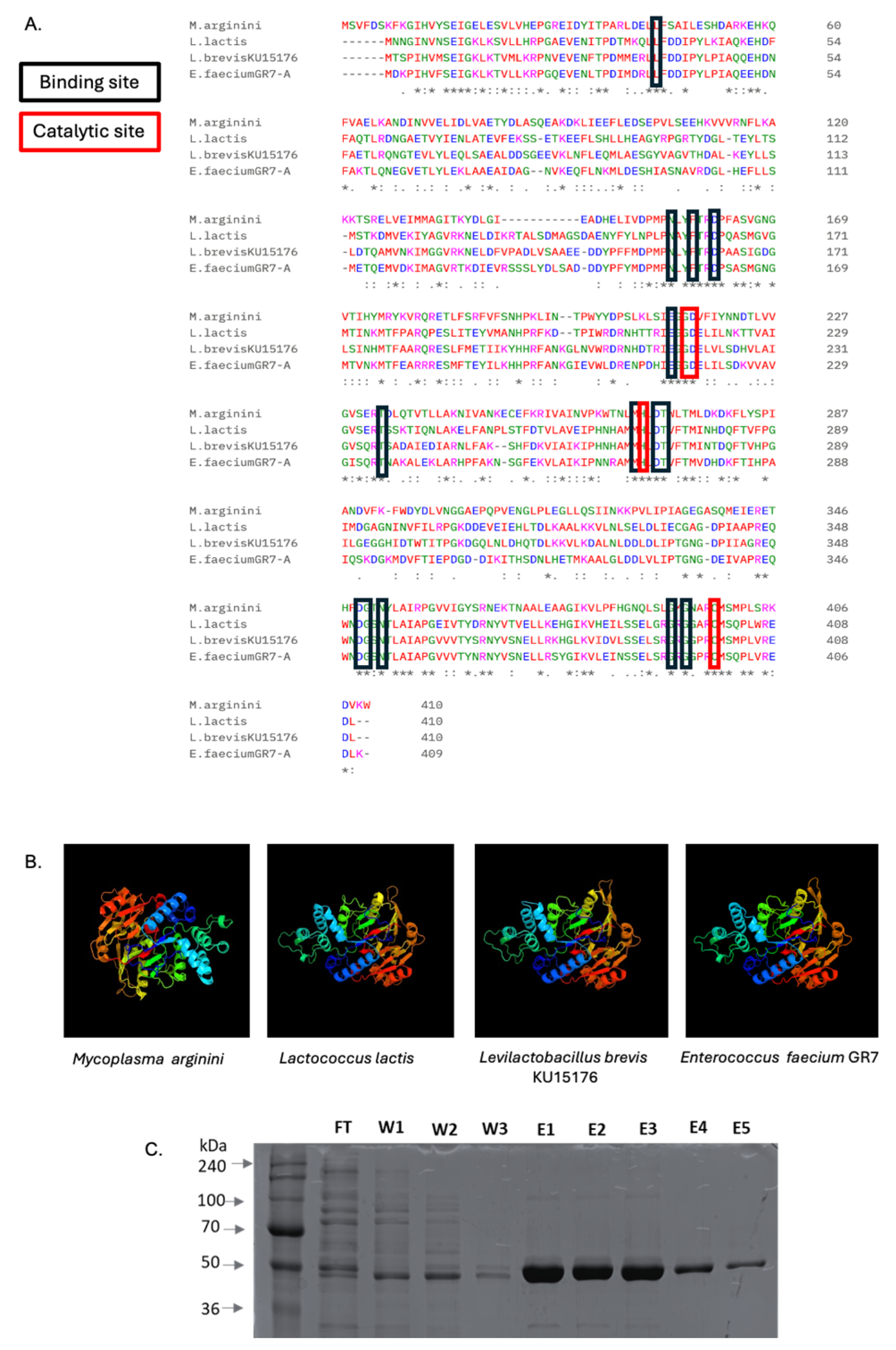
2.2. Arginine Deiminase of L. brevis KU15176
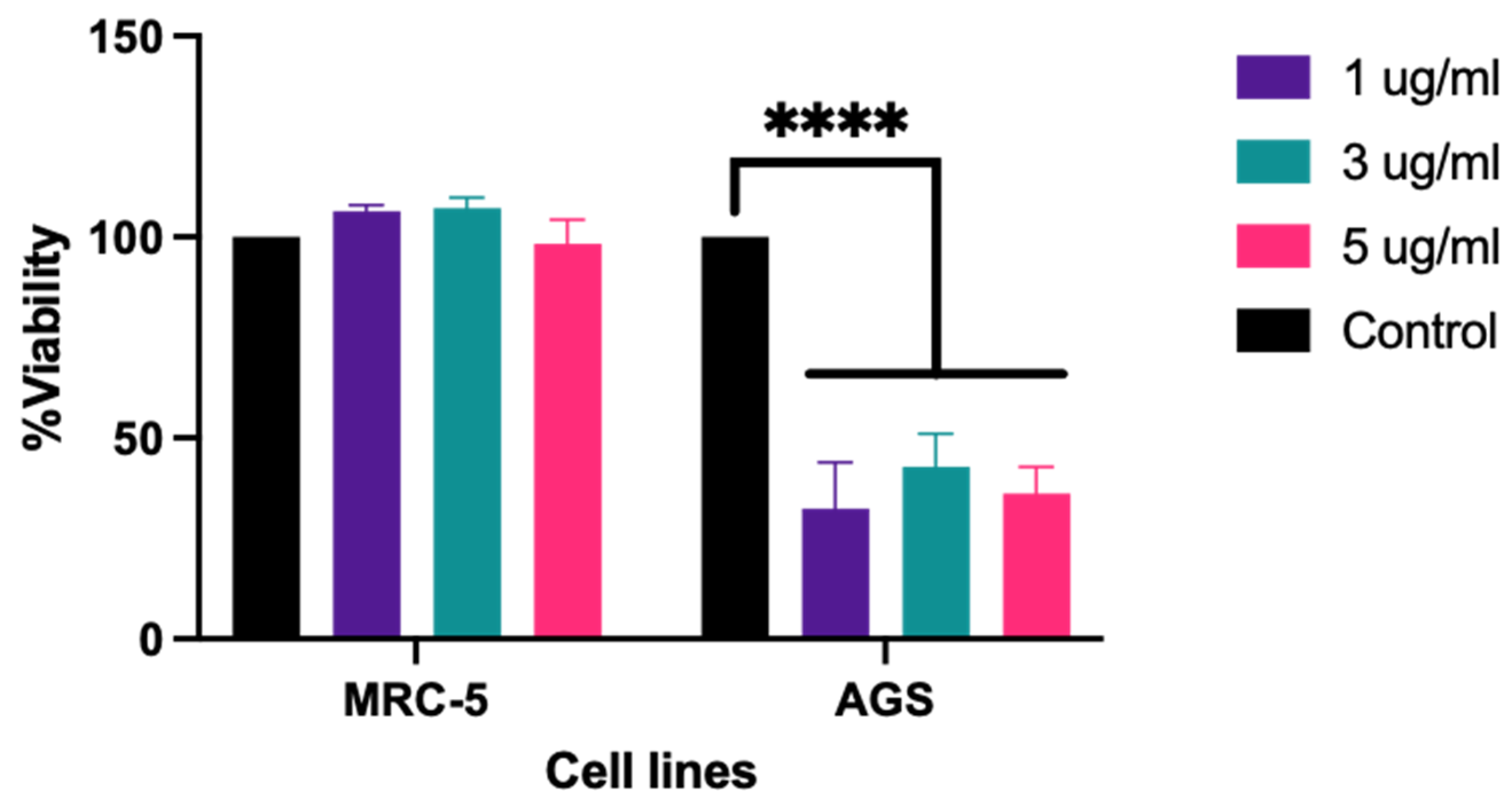
2.3. ADI_br Exhibited Antiproliferative Activity on AGS Cells
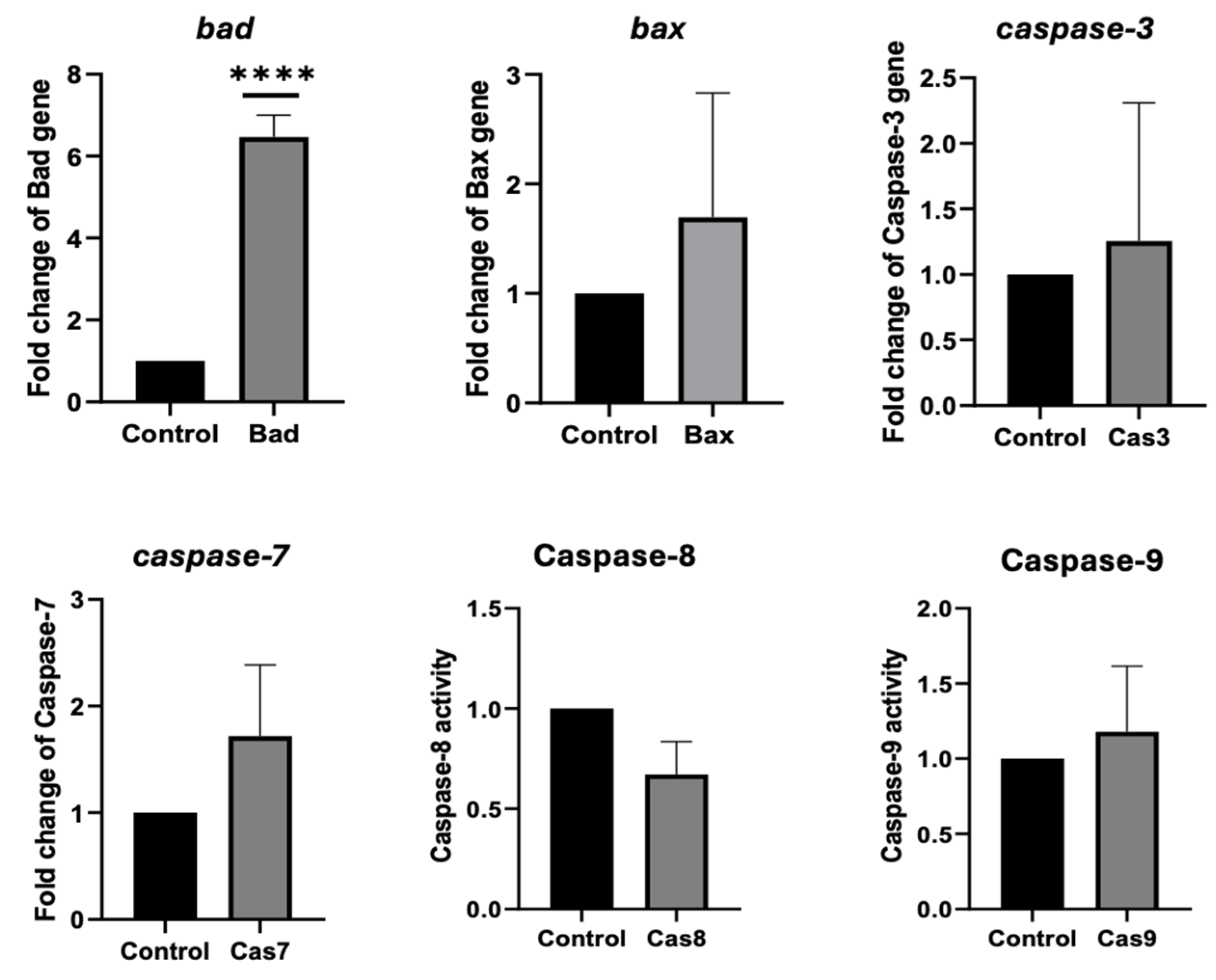
2.4. ADI_br Potentially Induced Apoptosis in AGS Cells
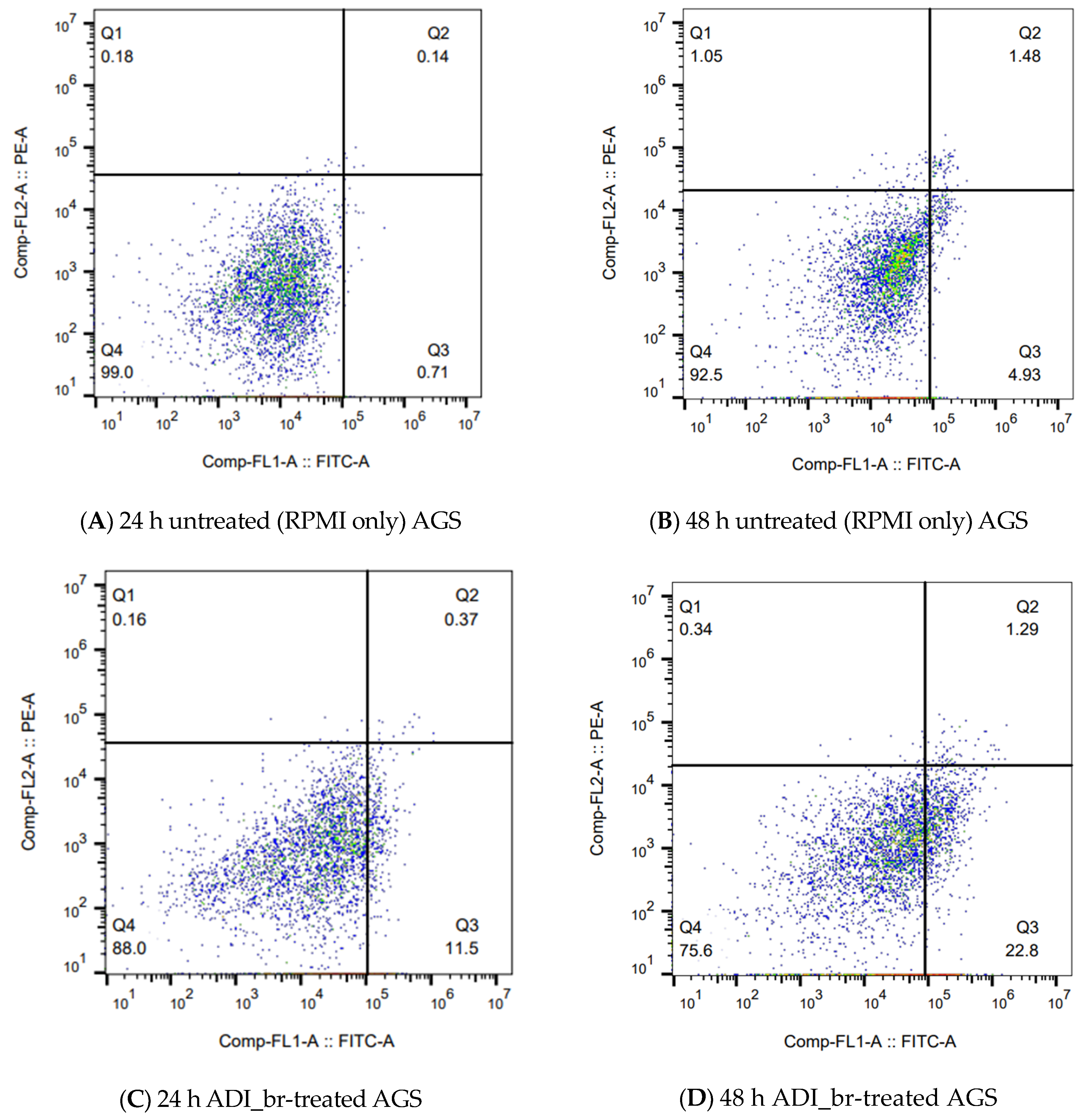
2.5. Flow Cytometry Confirmed the Induction and Progression of Apoptosis in ADI_br-Treated AGS Cells
3. Discussion
4. Materials and Methods
4.1. Cultivation of Bacterial and Mammalian Cells
4.2. DNA Extraction, Whole Genome Sequencing, Assembly, Annotation, and Gene Mining
4.3. Cloning of ADI_br
4.4. Overexpression of ADI_br
4.5. Purification of ADI_br
4.6. MTT Assay
4.7. RNA Extraction and RT-qPCR
4.8. Caspase-8 and Caspase-9 Colorimetric Assay
4.9. Apoptosis Analysis Using Flow Cytometry
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, J.; Zhang, Y.; Ye, L.; Wang, C. The anti-cancer effects and mechanisms of lactic acid bacteria exopolysaccharides in vitro: A review. Carbohydr. Polym. 2021, 253, 117308. [Google Scholar] [CrossRef] [PubMed]
- Garbacz, K. Anticancer activity of lactic acid bacteria. Semin. Cancer Biol. 2022, 86, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zheng, J.; Ou, X.; Han, Y. Anti-cancer Substances and Safety of Lactic Acid Bacteria in Clinical Treatment. Front. Microbiol. 2021, 12, 722052. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.; Van Der Meer, L.T.; Van Leeuwen, F.N. Amino Acid Depletion Therapies: Starving Cancer Cells to Death. Trends Endocrinol. Metab. 2021, 32, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Patil, M.; Bhaumik, J.; Babykutty, S.; Banerjee, U.; Fukumura, D. Arginine dependence of tumor cells: Targeting a chink in cancer’s armor. Oncogene 2016, 35, 4957–4972. [Google Scholar] [CrossRef] [PubMed]
- Al-Koussa, H.; El Mais, N.; Maalouf, H.; Abi-Habib, R.; El-Sibai, M. Arginine deprivation: A potential therapeutic for cancer cell metastasis? A review. Cancer Cell Int. 2020, 20, 150. [Google Scholar] [CrossRef] [PubMed]
- Endicott, M.; Jones, M.; Hull, J. Amino acid metabolism as a therapeutic target in cancer: A review. Amino Acids 2021, 53, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Wilder, C.S.; Chen, Z.; DiGiovanni, J. Pharmacologic approaches to amino acid depletion for cancer therapy. Mol. Carcinog. 2022, 61, 127–152. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Kaur, R. Purification of a dimeric arginine deiminase from Enterococcus faecium GR7 and study of its anti-cancerous activity. Protein Expr. Purif. 2016, 125, 53–60. [Google Scholar] [CrossRef]
- Kim, J.E.; Jeong, D.W.; Lee, H.J. Expression, purification, and characterization of arginine deiminase from Lactococcus lactis ssp. lactis ATCC 7962 in Escherichia coli BL21. Protein Expr. Purif. 2007, 53, 9–15. [Google Scholar] [CrossRef]
- Xiong, L.; Teng, J.L.L.; Botelho, M.G.; Lo, R.C.; Lau, S.K.P.; Woo, P.C.Y. Arginine Metabolism in Bacterial Pathogenesis and Cancer Therapy. Int. J. Mol. Sci. 2016, 17, 363. [Google Scholar] [CrossRef] [PubMed]
- Takaku, H.; Matsumoto, M.; Misawa, S.; Miyazaki, K. Anti-tumor activity of arginine deiminase from Mycoplasma arginini and its growth-inhibitory mechanism. Jpn. J. Cancer Res. 1995, 86, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Di Marzio, L.; Russo, F.P.; Simona, D.A.; Biordi, L.; Ulisse, S.; Amicosante, G.; Simone, C.D.; Cifone, M.G. Apoptotic effects of selected strains of lactic acid bacteria on a human T leukemia cell line are associated with bacterial arginine deiminase and/or sphingomyelinase activities. Nutr. Cancer. 2001, 40, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.H.; Lee, N.K.; Paik, H.D. The Anti-Cancer Potential of Heat-Killed Lactobacillus brevis KU15176 upon AGS Cell Lines through Intrinsic Apoptosis Pathway. Int. J. Mol. Sci. 2022, 23, 4073. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Kim, S.Y.; Lee, K.W.; Lee, H.J. Arginine deiminase originating from Lactococcus lactis ssp. lactis American Type Culture Collection (ATCC) 7962 induces G 1-phase cell-cycle arrest and apoptosis in SNU-1 stomach adenocarcinoma cells. Br. J. Nut. 2009, 102, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, R.M.; Kim, S.H.; Vasquez, R.; Hwang, I.C.; Park, Y.S.; Paik, H.D.; Kang, D.K. Bioinformatics and its role in the study of the evolution and probiotic potential of lactic acid bacteria. Food Sci. Biotechnol. 2022, 32, 389–412. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-J.; Jiang, S.S.; Hung, W.C.; Borthakur, G.; Lin, S.F.; Pemmaraju, N.; Jabbour, E.; Bomalaski, J.S.; Chen, Y.P.; Hsiao, H.H.; et al. A Phase II Study of Arginine Deiminase (ADI-PEG20) in Relapsed/Refractory or Poor-Risk Acute Myeloid Leukemia Patients. Sci. Rep. 2017, 7, 11253. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Kanneganti, T.D. Caspase-7: A protease involved in apoptosis and inflammation. Int. J. Biochem.Cell Biol. 2010, 42, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Wlodkowic, D.; Skommer, J.; Darzynkiewicz, Z. Flow cytometry-based apoptosis detection. Methods Mol. Biol. 2009, 559, 19–32. [Google Scholar] [PubMed]
- Amer, M.N.; Mansour, N.M.; El-Diwany, A.I.; Dawoud, I.E.; Rashad, F.M. Isolation of probiotic lactobacilli strains harboring l-asparaginase and arginine deiminase genes from human infant feces for their potential application in cancer prevention. Ann. Microbiol. 2013, 63, 1121–1129. [Google Scholar] [CrossRef]
- Han, K.J.; Lee, J.E.; Lee, N.K.; Paik, H.D. Antioxidant and Anti-inflammatory effect of probiotic Lactobacillus plantarum KU15149 derived from Korean homemade Diced-radish kimchi. J. Microbiol. Biotechnol. 2020, 30, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive κ-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Terry, D.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Seemann, T. Genome analysis Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. Genome analysis QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.; Parkhill, J.; Crook, J.; Horsnell, T.; Rice, P.; Rajandream, M.-A.; Barrell, B. Artemis: Sequence visualization and annotation. Bioinformatics 2000, 16, 944–945. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction, and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef] [PubMed]
- Rho, H.-W.; Lee, B.-C.; Choi, E.-S.; Choi, I.-J.; Lee, Y.-S.; Goh, S.-H. Identification of valid reference genes for gene expression studies of human stomach cancer by reverse transcription-qPCR. BMC Cancer 2010, 10, 240. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anticancer-Associated Metabolites/Genes | Annotation/Gene Product |
|---|---|
| Arginine deiminase | |
| arcA | Arginine deiminase |
| S-layer protein | |
| LbrevisKU15176_01469 | S-layer protein |
| Exopolysaccharide (Biosynthesis) | |
| YwqE | Mn-dependent protein-tyrosine phosphatase |
| EpsC | Tyrosine-protein kinase transmembrane modulator |
| YwqD | Tyrosine-protein kinase |
| EpsE | Undecaprenyl-phosphate galactosephosphorotransferase |
| EpsF | Putative glycosyltransferase |
| EpsH | Putative glycosyltransferase |
| EpsL | Putative sugar transferase |
| LbrevisKU15176_01006 | dTDP-glucose 4,6-dehydratase |
| LbrevisKU15176_01013 | UDP-galactopyranose mutase |
| MurJ | Flippase |
| YwqC | Putative capsular polysaccharide biosynthesis protein |
| Peptidoglycan (Recycling and Biosynthesis) | |
| glmM | Phosphoglucosamine mutase |
| glmU | N-acetylglucosamine-1-phosphate uridyltransferase |
| murA1_1 | UDP-N-acetylglucosamine 1-carboxyvinyltransferase 1 |
| murA1_2 | UDP-N-acetylglucosamine 1-carboxyvinyltransferase 1 |
| LbrevisKU15176_00604 | N-acetylmuramoyl-L-alanine amidase |
| LbrevisKU15176_00626 | N-acetylmuramoyl-L-alanine amidase |
| Genes | Sequence (5′-3′) | Product Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| B2M | |||
| Forward | AGATGAGTATGCCTGCCGTG | 105 | 55.5 |
| Reverse | GCGGCATCTTCAAACCTCCA | ||
| bax | |||
| Forward | CCCCCGAGAGGTCTTTTTCC | 160 | 54 |
| Reverse | TGTCCAGCCCATGATGGTTC | ||
| bad | |||
| Forward | CCTTTAAGAAGGGACTTCCTCGCC | 325 | 54 |
| Reverse | ACTTCCGATGGGACCAAGCCTTCC | ||
| caspase-3 | |||
| Forward | AGCGGATGGGTGCTATTGTG | 172 | 54 |
| Reverse | ACACCCACCGAAAACCAGAG | ||
| caspase-7 | |||
| Forward | ACGATGGCAGATGATCAGGG | 85 | 54 |
| Reverse | GGTCTGGCTTAGCATCCACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendoza, R.M.; Song, J.H.; Jung, Y.T.; Paik, H.-D.; Park, Y.-S.; Kang, D.-K. Recombinant Arginine Deiminase from Levilactobacillus brevis Inhibits the Growth of Stomach Cancer Cells, Possibly by Activating the Intrinsic Apoptosis Pathway. Int. J. Mol. Sci. 2024, 25, 4163. https://doi.org/10.3390/ijms25084163
Mendoza RM, Song JH, Jung YT, Paik H-D, Park Y-S, Kang D-K. Recombinant Arginine Deiminase from Levilactobacillus brevis Inhibits the Growth of Stomach Cancer Cells, Possibly by Activating the Intrinsic Apoptosis Pathway. International Journal of Molecular Sciences. 2024; 25(8):4163. https://doi.org/10.3390/ijms25084163
Chicago/Turabian StyleMendoza, Remilyn M., Ji Hoon Song, Yong Tae Jung, Hyun-Dong Paik, Young-Seo Park, and Dae-Kyung Kang. 2024. "Recombinant Arginine Deiminase from Levilactobacillus brevis Inhibits the Growth of Stomach Cancer Cells, Possibly by Activating the Intrinsic Apoptosis Pathway" International Journal of Molecular Sciences 25, no. 8: 4163. https://doi.org/10.3390/ijms25084163
APA StyleMendoza, R. M., Song, J. H., Jung, Y. T., Paik, H.-D., Park, Y.-S., & Kang, D.-K. (2024). Recombinant Arginine Deiminase from Levilactobacillus brevis Inhibits the Growth of Stomach Cancer Cells, Possibly by Activating the Intrinsic Apoptosis Pathway. International Journal of Molecular Sciences, 25(8), 4163. https://doi.org/10.3390/ijms25084163