
KYNA Ameliorates Glutamate Toxicity of HAND by Enhancing Glutamate Uptake in A2 Astrocytes
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
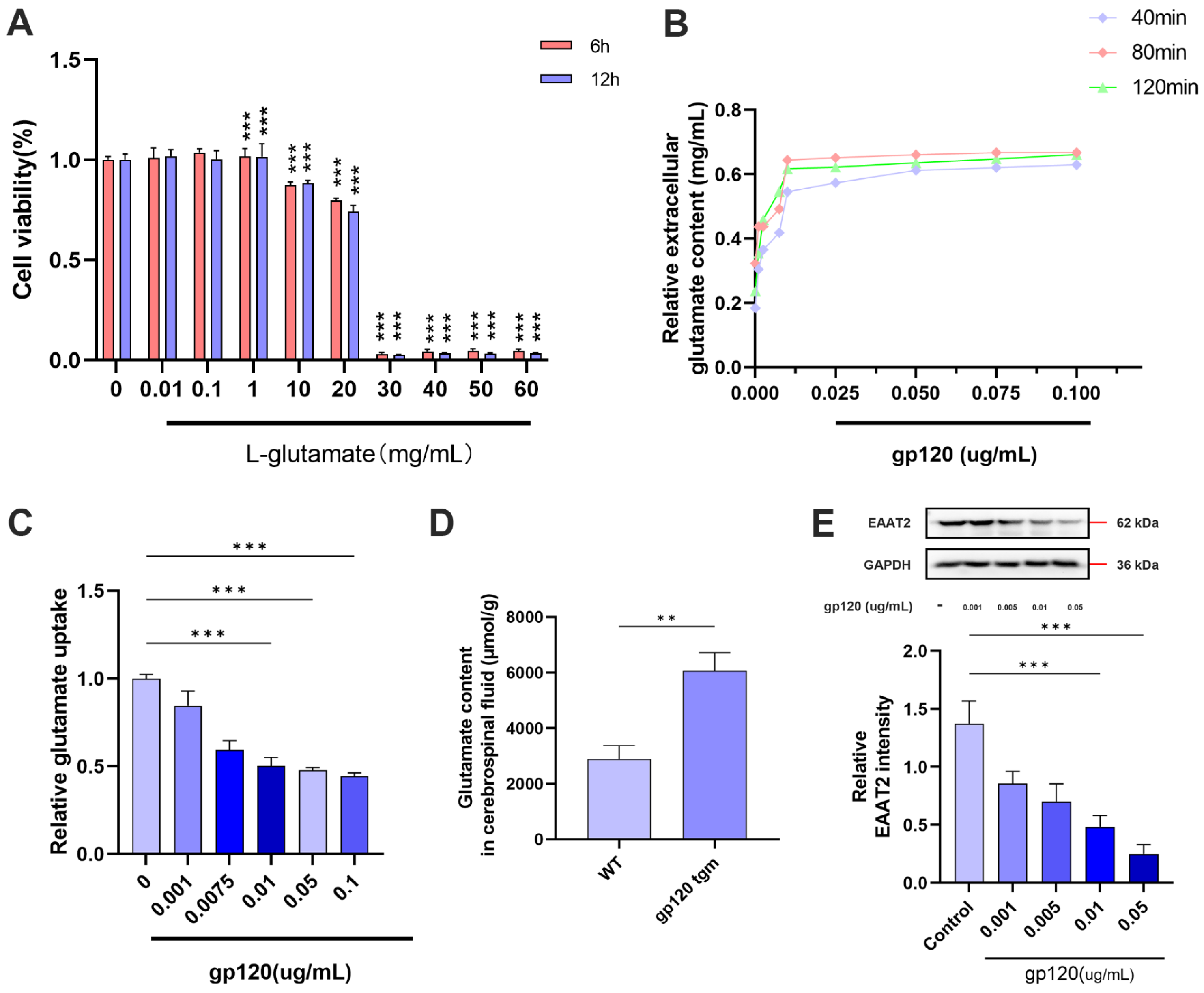
2.1. HIV-1 gp120 Inhibits A2 Astrocyte Formation and Induces Glutamatergic Toxicity
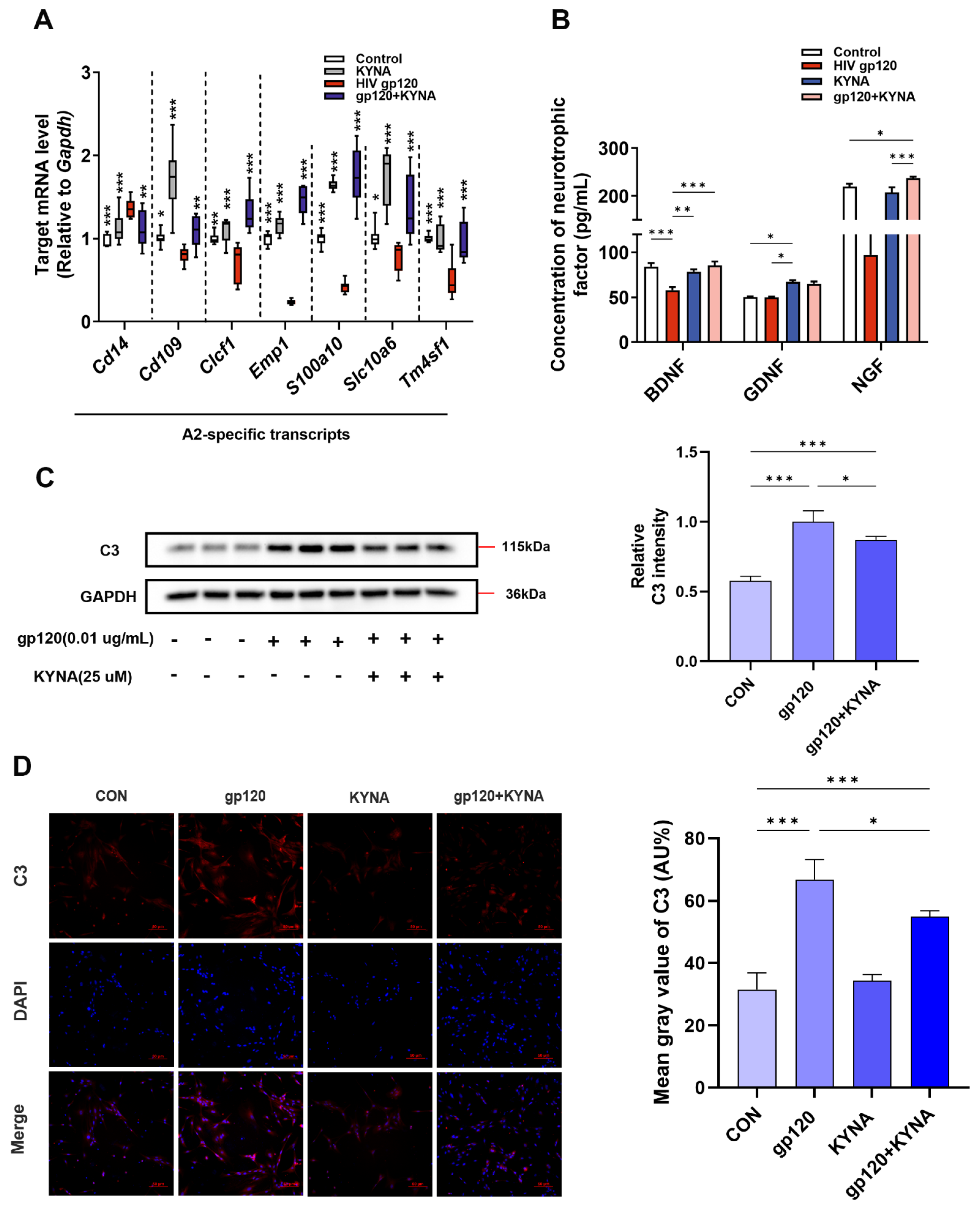
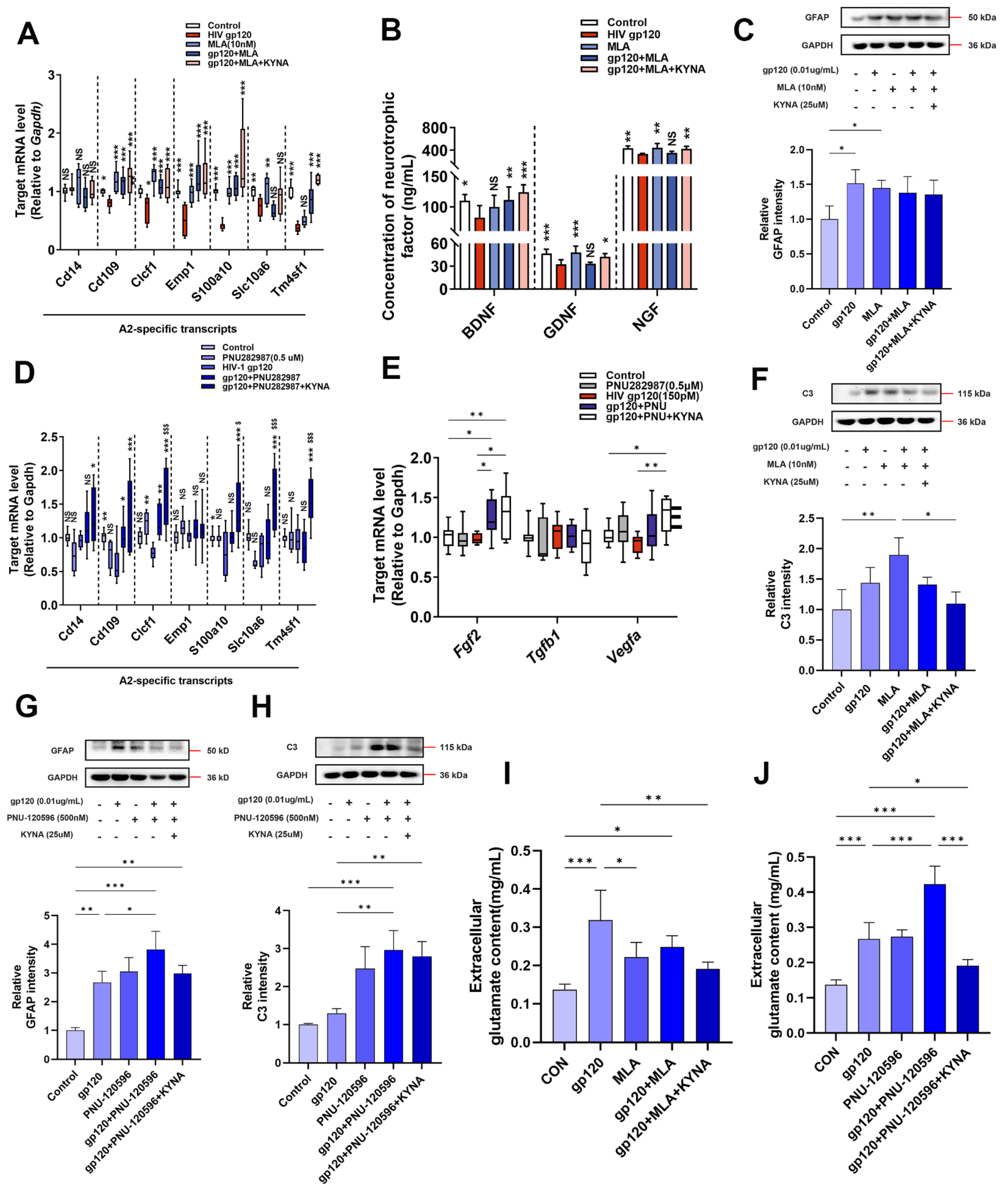
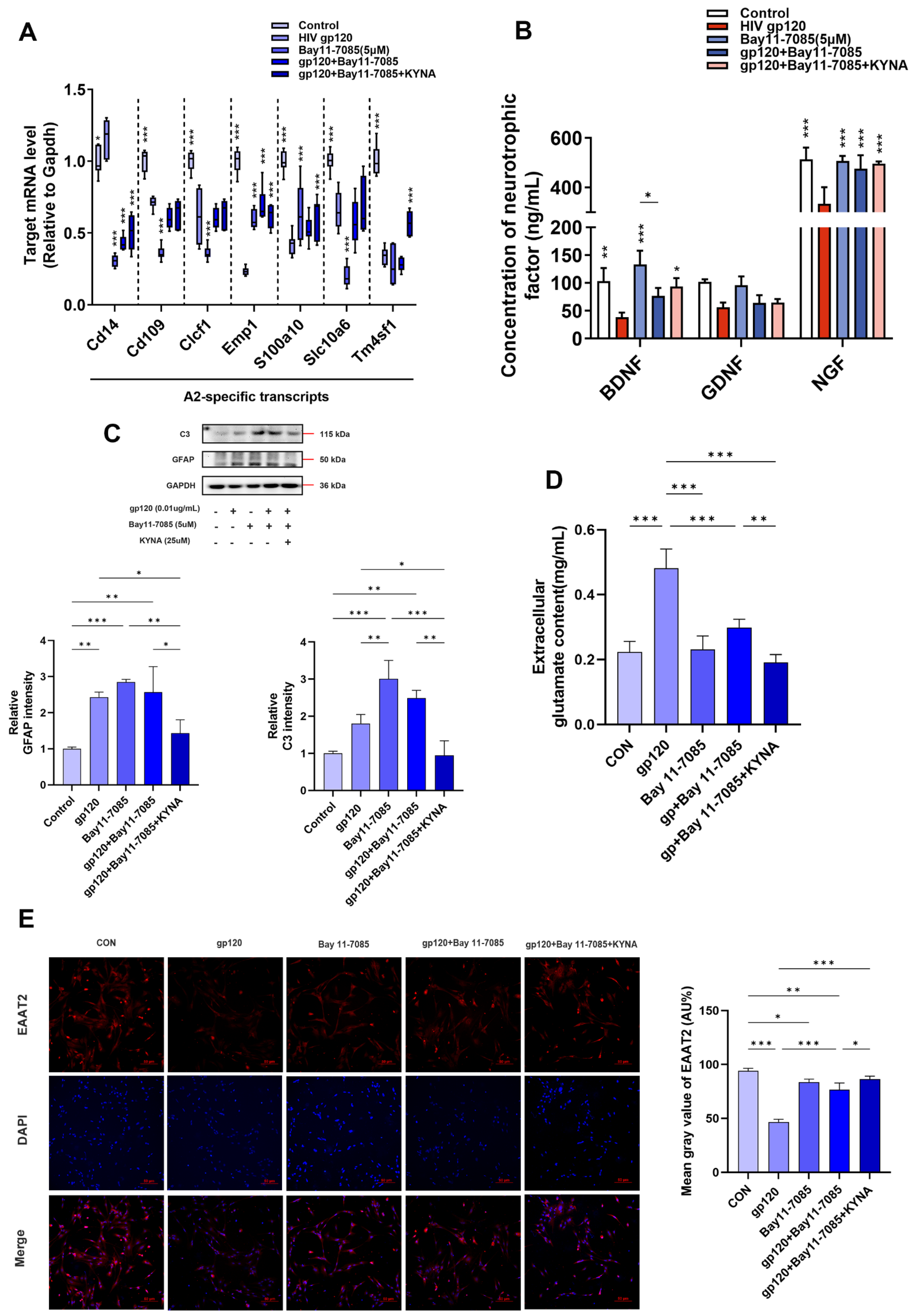
2.2. KYNA Promotes A2 Astrocyte Formation and Reduces gp120-Induced Glutamate Toxicity
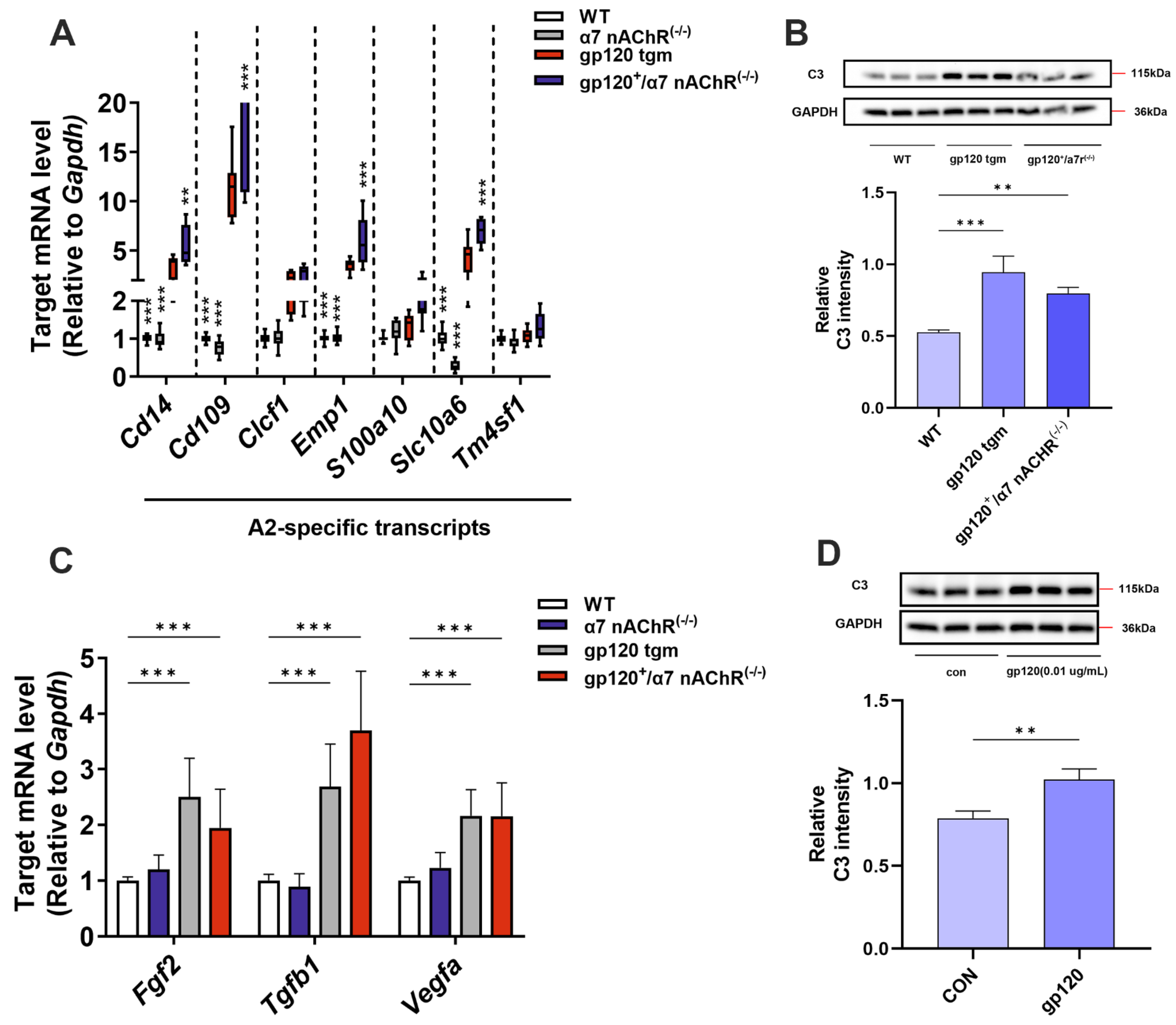
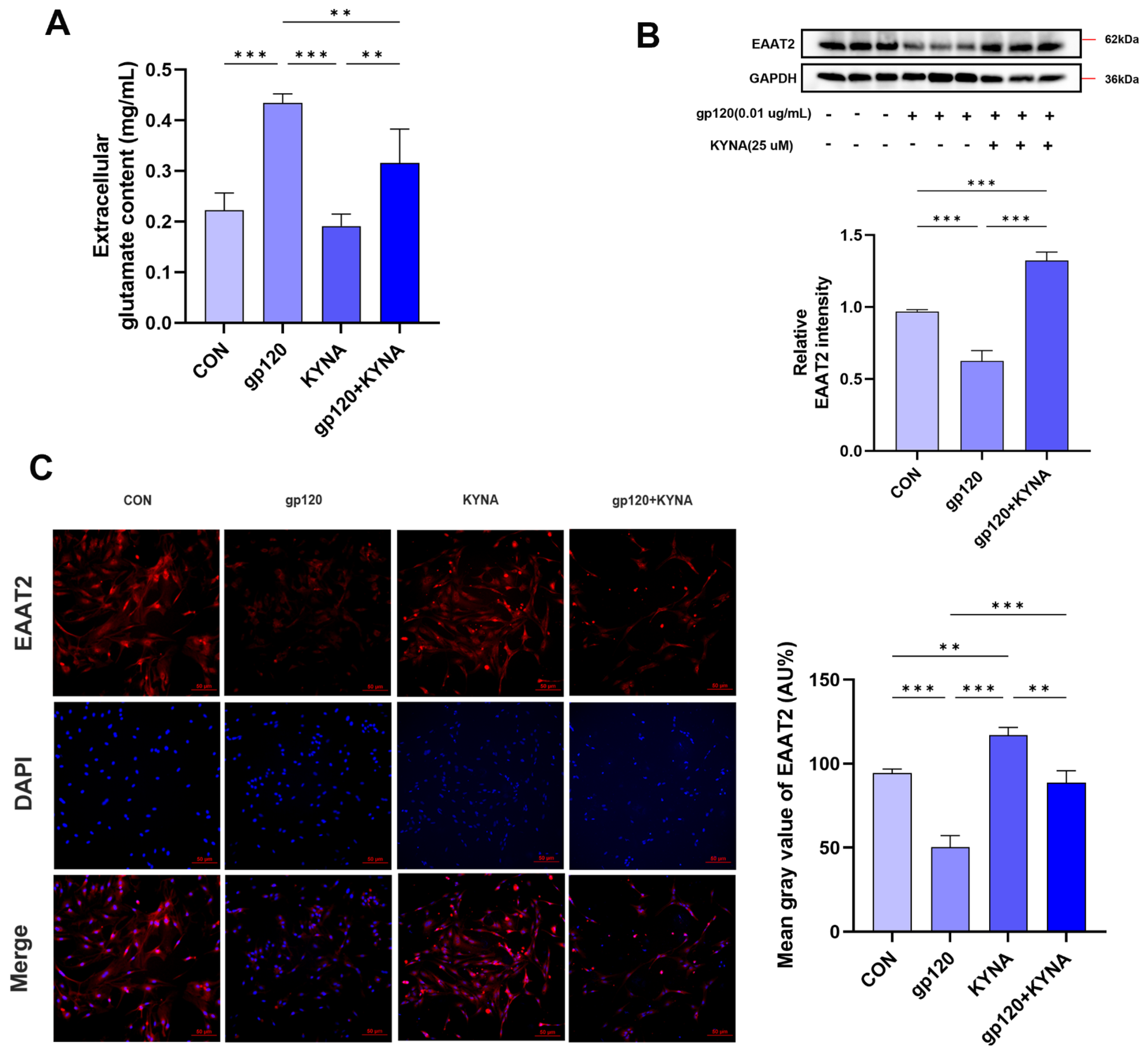
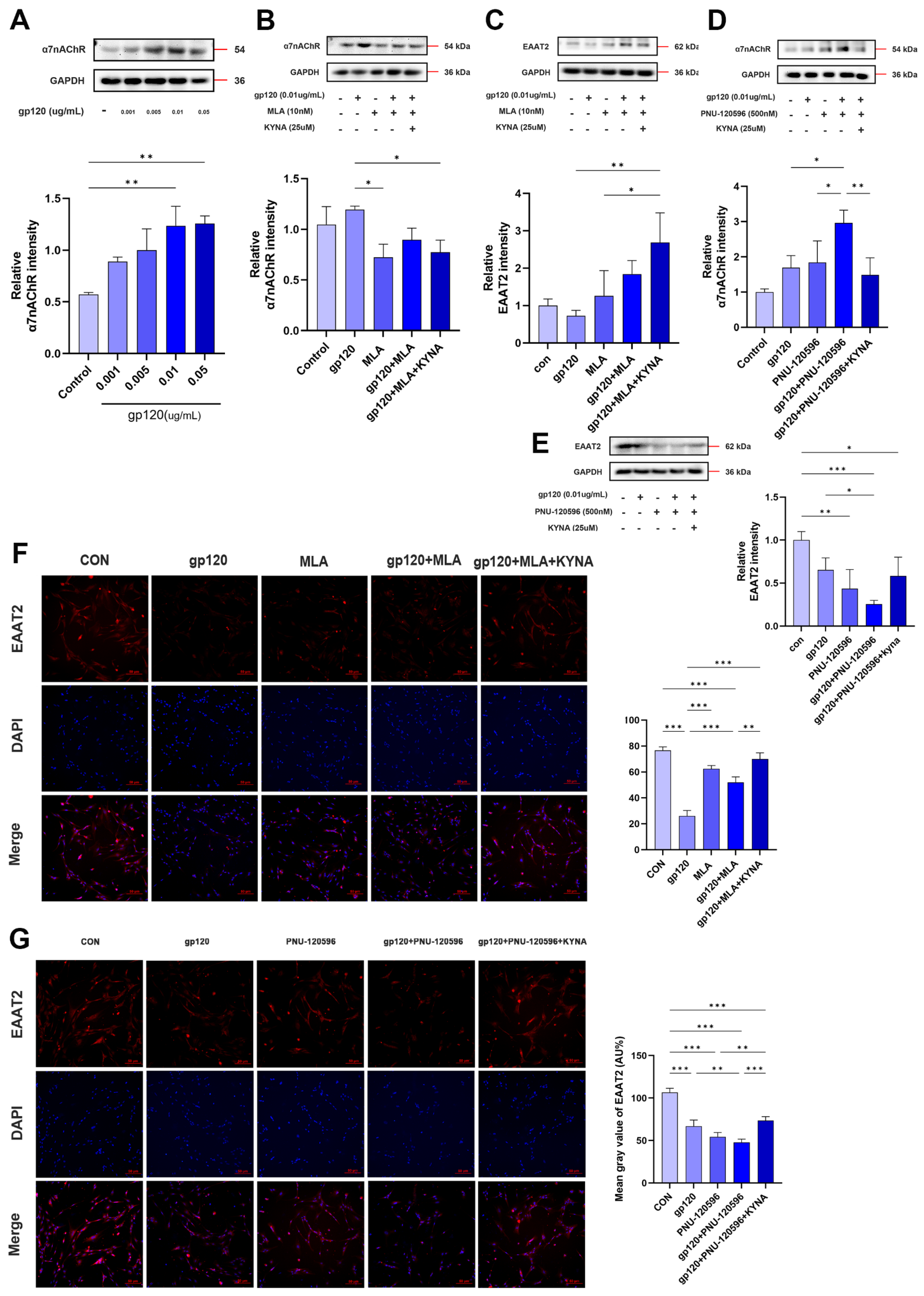
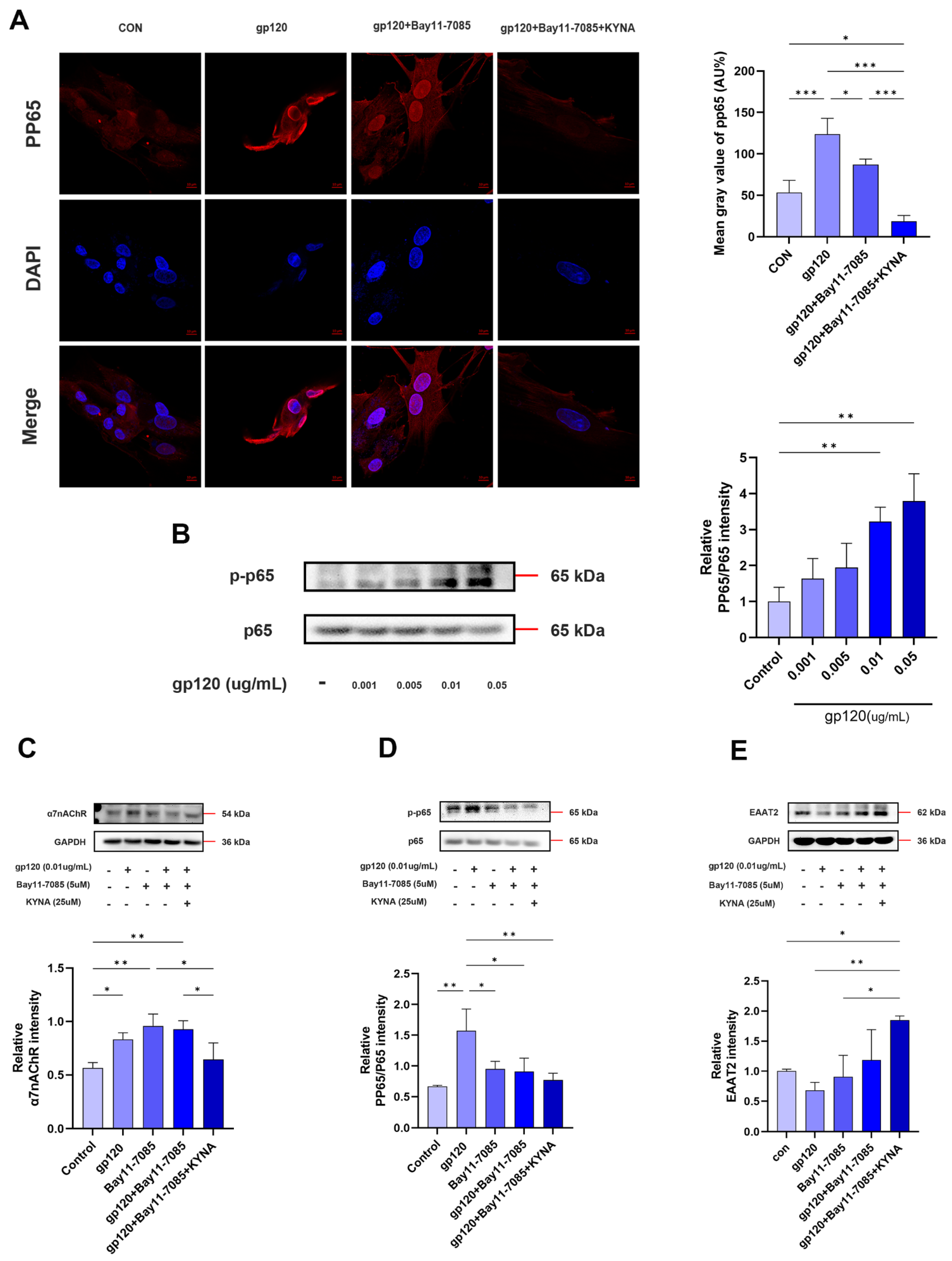
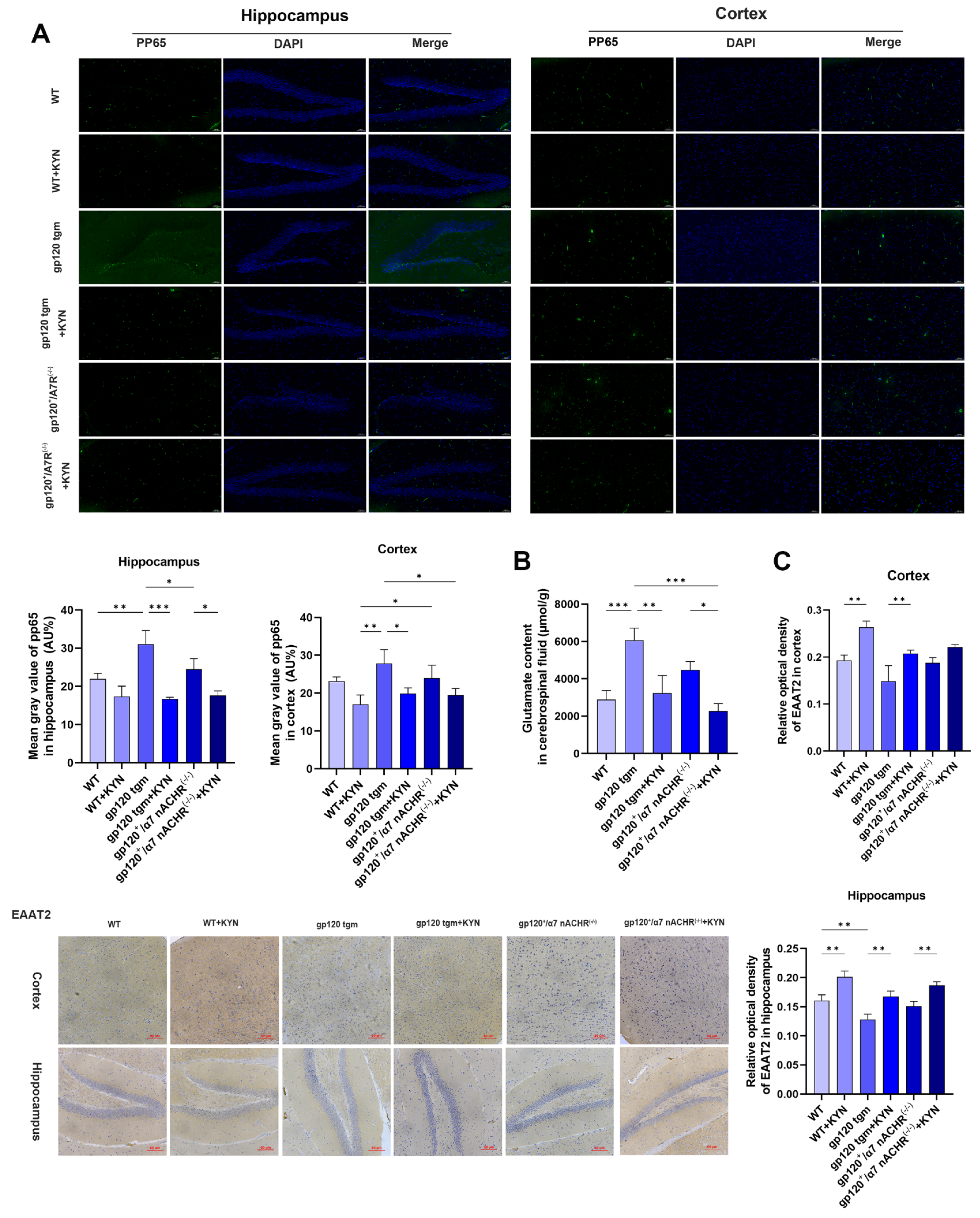
2.3. KYNA Enhances Glutamate Uptake in A2 Astrocytes by Blocking α7nAChR
2.4. KYNA Promotes A2 Astrocyte EAAT2 Expression by Blocking α7nAChR/NF-κB Signaling Activation
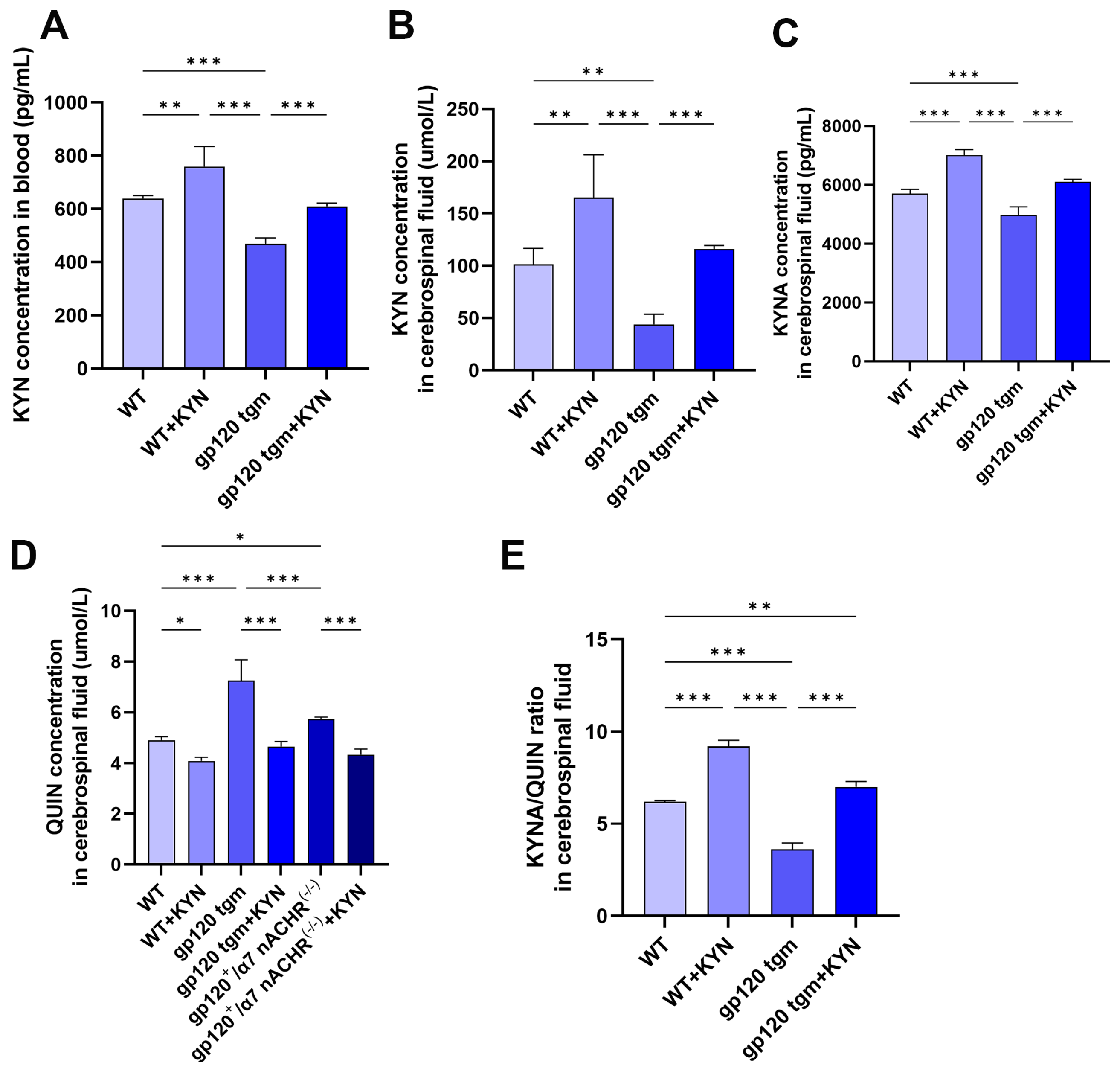
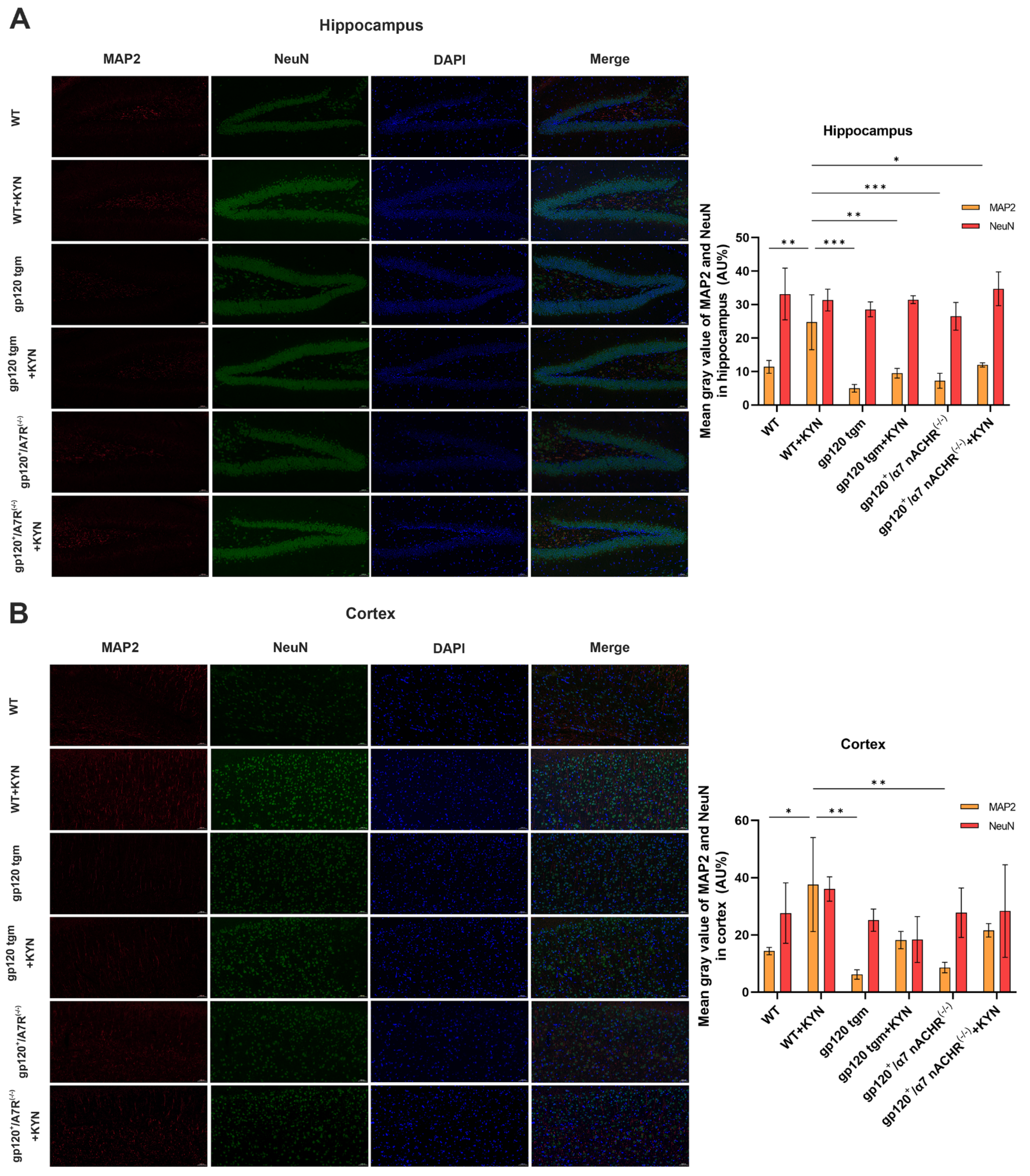
2.5. Intraperitoneal Injection of KYN and PROB Effectively Increased KYNA Content in the Brain of gp120 tgm Mice and Reduced Neuronal Damage
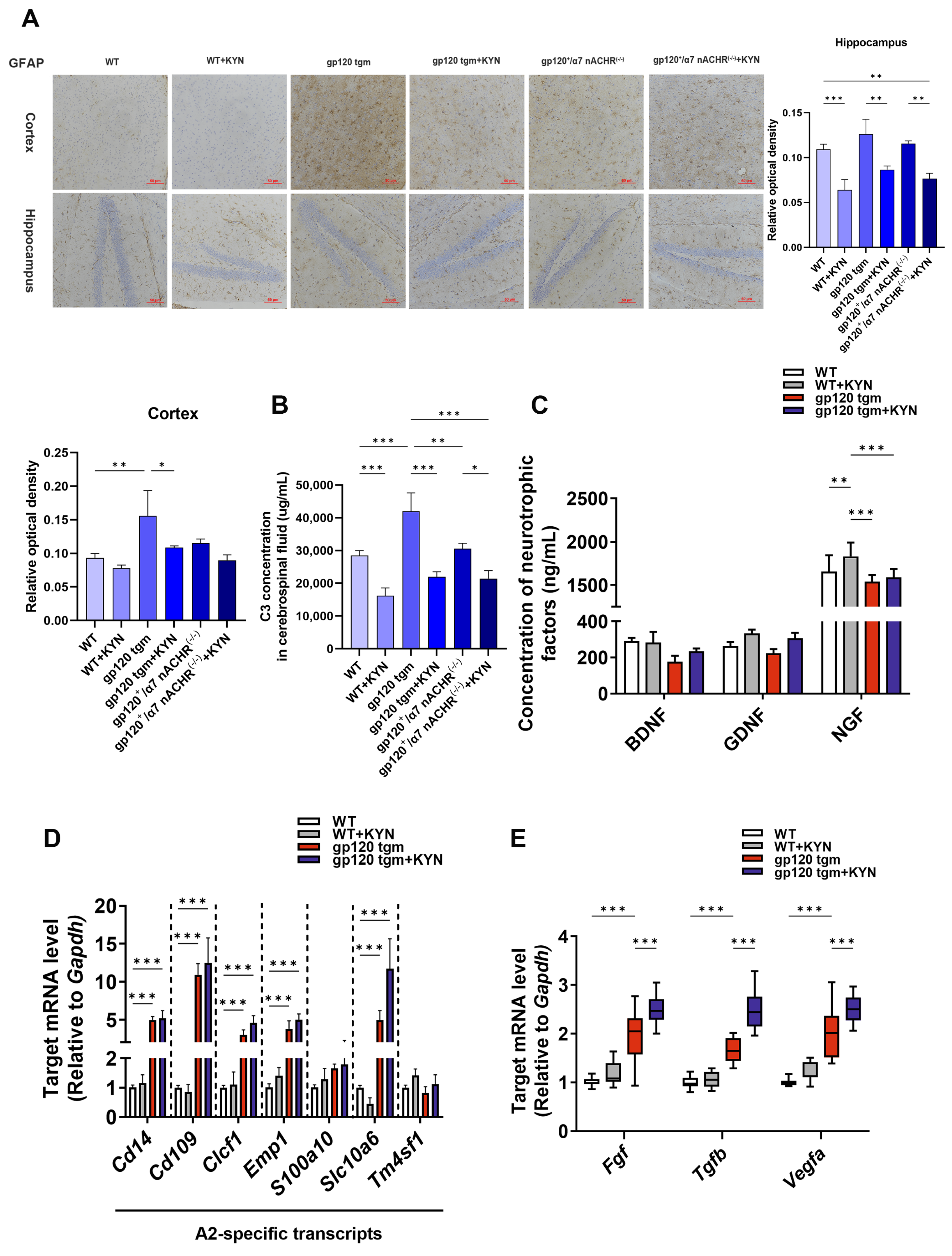
2.6. Intraperitoneal Injection of KYN and PROB Ameliorates Glutamate Toxicity in gp120 tgm Mice by Promoting A2 Astrocyte Formation
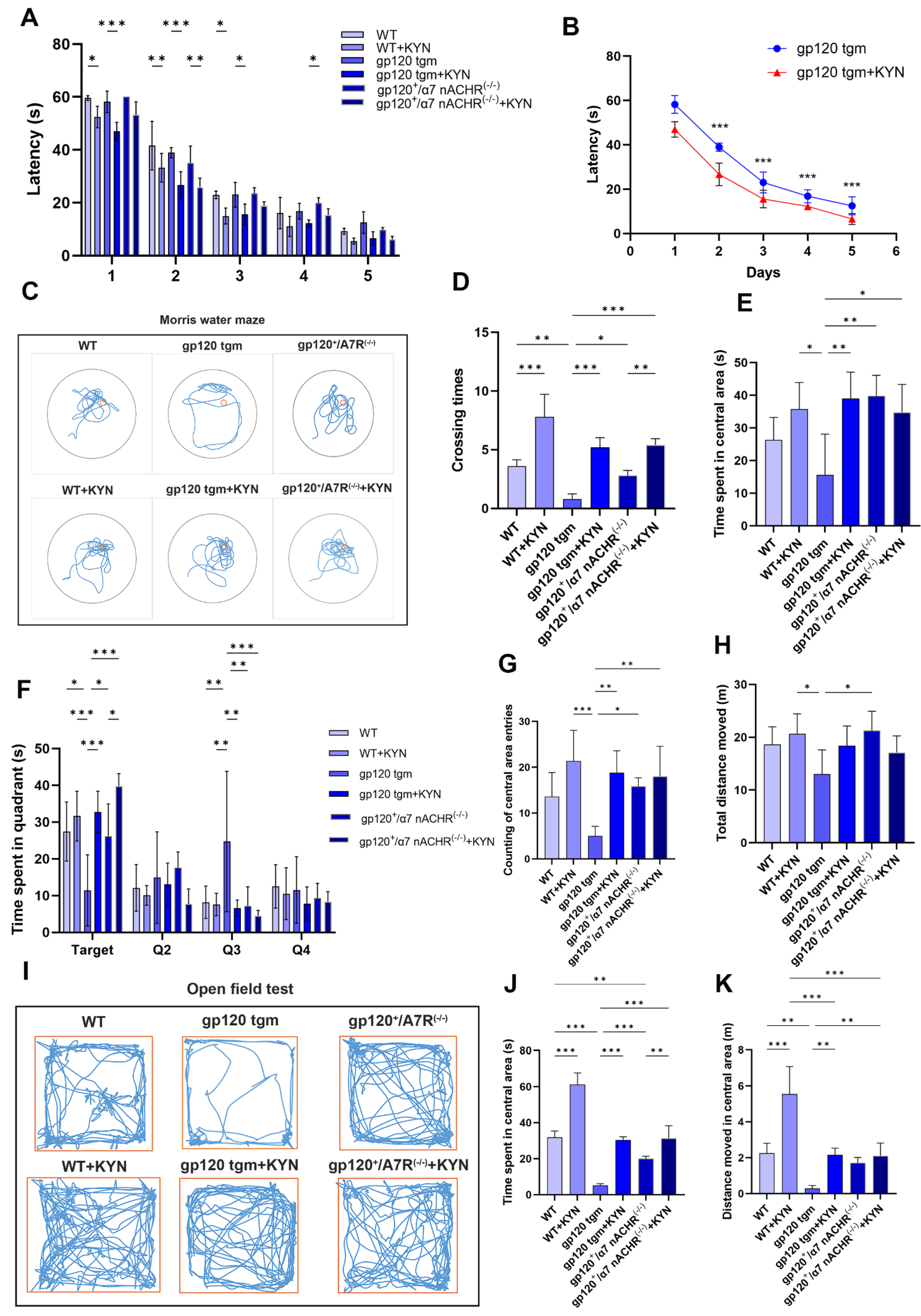
2.7. Intraperitoneal Injection of KYN and PROB Improves Cognitive Function in gp120 tgm Mice
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of Primary Astrocytes and Drug Treatment
4.2. Quantitative Reverse Transcription PCR (RT-qPCR) Analysis
4.3. Western Blotting
4.4. Glutamate Uptake Assay
4.5. Immunofluorescence
4.6. Animal Models and Treatments
4.7. Behavioral Testing
4.8. Immunohistochemistry and Immunofluorescence
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- The Joint United Nations Programme on HIV/AIDS (UNAIDS). Global HIV & AIDS Statistics—Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 31 August 2023).
- Abbate, M.T.A.; Ramöller, I.K.; Sabri, A.H.; Paredes, A.J.; Hutton, A.J.; McKenna, P.E.; Peng, K.; Hollett, J.A.; McCarthy, H.O.; Donnelly, R.F. Formulation of antiretroviral nanocrystals and development into a microneedle delivery system for potential treatment of HIV-associated neurocognitive disorder (HAND). Int. J. Pharmaceut. 2023, 640, 123005. [Google Scholar] [CrossRef] [PubMed]
- Chen, B. Molecular Mechanism of HIV-1 Entry. Trends Microbiol. 2019, 27, 878–891. [Google Scholar] [CrossRef] [PubMed]
- He, X.L.; Yang, W.J.; Zeng, Z.J.; Wei, Y.; Gao, J.; Zhang, B.; Li, L.; Liu, L.Q.; Wan, Y.; Zeng, Q.; et al. NLRP3-dependent pyroptosis is required for HIV-1 gp120-induced neuropathology. Cell Mol. Immunol. 2020, 17, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.Z.; Qu, Z.; Hashimoto, H.; Gradinaru, V.; Kawaguchi, R.; Khakh, B.S. Molecular basis of astrocyte diversity and morphology across the CNS in health and disease. Science 2022, 378, eadc9020. [Google Scholar] [CrossRef] [PubMed]
- Lun, J.X.; Li, Y.B.; Gao, X.F.; Gong, Z.L.; Chen, X.L.; Zou, J.H.; Zhou, C.X.; Huang, Y.Y.; Zhou, B.L.; Huang, P.W.; et al. Kynurenic acid blunts A1 astrocyte activation against neurodegeneration in HIV-associated neurocognitive disorders. J. Neuroinflamm. 2023, 20, 87. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef]
- Gautam, D.; Naik, U.P.; Naik, M.U.; Yadav, S.K.; Chaurasia, R.N.; Dash, D. Glutamate Receptor Dysregulation and Platelet Glutamate Dynamics in Alzheimer’s and Parkinson’s Diseases: Insights into Current Medications. Biomolecules 2023, 13, 1609. [Google Scholar] [CrossRef]
- Potter, M.C.; Figuera-Losada, M.; Rojas, C.; Slusher, B.S. Targeting the Glutamatergic System for the Treatment of HIV-Associated Neurocognitive Disorders. J. Neuroimmune Pharm. 2013, 8, 594–607. [Google Scholar] [CrossRef]
- Guillemin, G.J.; Kerr, S.J.; Smythe, G.A.; Smith, D.G.; Kapoor, V.; Armati, P.J.; Croitoru, J.; Brew, B.J. Kynurenine pathway metabolism in human astrocytes: A paradox for neuronal protection. J. Neurochem. 2001, 78, 842–853. [Google Scholar] [CrossRef]
- Ostapiuk, A.; Urbanska, E.M. Kynurenic acid in neurodegenerative disorders-unique neuroprotection or double-edged sword? CNS Neurosci. Ther. 2022, 28, 19–35. [Google Scholar] [CrossRef]
- Moroni, F.; Cozzi, A.; Sili, M.; Mannaioni, G. Kynurenic acid: A metabolite with multiple actions and multiple targets in brain and periphery. J. Neural Transm. 2012, 119, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Majláth, Z.; Török, N.; Toldi, J.; Vécsei, L. Memantine and Kynurenic Acid: Current Neuropharmacological Aspects. Curr. Neuropharmacol. 2016, 14, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Arnaiz-Cot, J.J.; González, J.C.; Sobrado, M.; Baldelli, P.; Carbone, E.; Gandia, L.; García, A.G.; Hernández-Guijo, J.M. Allosteric modulation of α7 nicotinic receptors selectively depolarizes hippocampal interneurons, enhancing spontaneous GABAergic transmission. Eur. J. Neurosci. 2008, 27, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.W. Kynurenic acid blocks nicotinic synaptic transmission to hippocampal interneurons in young rats. Eur. J. Neurosci. 2007, 25, 2656–2665. [Google Scholar] [CrossRef]
- Fan, Y.Y.; Huo, J. A1/A2 astrocytes in central nervous system injuries and diseases: Angels or devils? Neurochem. Int. 2021, 148, 105080. [Google Scholar] [CrossRef]
- Tavares, R.G.; Tasca, C.I.; Santos, C.E.; Alves, L.B.; Porciúncula, L.O.; Emanuelli, T.; Souza, D.O. Quinolinic acid stimulates synaptosomal glutamate release and inhibits glutamate uptake into astrocytes. Neurochem. Int. 2002, 40, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Connick, J.H.; Stone, T.W. Quinolinic acid effects on amino acid release from the rat cerebral cortex in vitro and in vivo. Br. J. Pharmacol. 1988, 93, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.Q.; Huang, Y.L.; Braun, A.L.; Tong, Z.H.; Zhao, R.Z.; Li, Y.J.; Liu, F.; Zheng, J.L.C. Glutaminase-containing microvesicles from HIV-1-infected macrophages and immune-activated microglia induce neurotoxicity. Mol. Neurodegener. 2015, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.V.; Schousboe, A.; Verkhratsky, A. Astrocyte energy and neurotransmitter metabolism in Alzheimer’s disease: Integration of the glutamate/GABA-glutamine cycle. Prog. Neurobiol. 2022, 217, 102331. [Google Scholar] [CrossRef]
- Vohra, M.; Lemieux, G.A.; Lin, L.; Ashrafi, K. Kynurenic acid accumulation underlies learning and memory impairment associated with aging. Gene Dev. 2018, 32, 14–19. [Google Scholar] [CrossRef]
- Plitman, E.; Iwata, Y.; Caravaggio, F.; Nakajima, S.; Chung, J.K.; Gerretsen, P.; Kim, J.; Takeuchi, H.; Chakravarty, M.M.; Remington, G.; et al. Kynurenic Acid in Schizophrenia: A Systematic Review and Meta-analysis. Schizophr. Bull. 2017, 43, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Pocivavsek, A.; Wu, H.Q.; Potter, M.C.; Elmer, G.I.; Pellicciari, R.; Schwarcz, R. Fluctuations in Endogenous Kynurenic Acid Control Hippocampal Glutamate and Memory. Neuropsychopharmacology 2011, 36, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Kozak, R.; Campbell, B.M.; Strick, C.A.; Horner, W.; Hoffmann, W.E.; Kiss, T.; Chapin, D.S.; McGinnis, D.; Abbott, A.L.; Roberts, B.M.; et al. Reduction of Brain Kynurenic Acid Improves Cognitive Function. J. Neurosci. 2014, 34, 10592–10602. [Google Scholar] [CrossRef] [PubMed]
- Hilmas, C.; Pereira, E.F.; Alkondon, M.; Rassoulpour, A.; Schwarcz, R.; Albuquerque, E.X. The brain metabolite kynurenic acid inhibits alpha7 nicotinic receptor activity and increases non-alpha7 nicotinic receptor expression: Physiopathological implications. J. Neurosci. 2001, 21, 7463–7473. [Google Scholar] [CrossRef] [PubMed]
- Pizzi, M.; Spano, P. Distinct roles of diverse nuclear factor-κB complexes in neuropathological mechanisms. Eur. J. Pharmacol. 2006, 545, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, H.Y.; Chen, Y.C.; Chen, H.M.; Tu, P.H.; Chern, Y.J. A critical role of astrocyte-mediated nuclear factor-κB-dependent inflammation in Huntington’s disease. Hum. Mol. Genet. 2013, 22, 1826–1842. [Google Scholar] [CrossRef] [PubMed]
- Chi, F.; Wang, L.; Zheng, X.Y.; Wu, C.H.; Jong, A.; Sheard, M.A.; Shi, W.; Huang, S.H. Meningitic K1 Penetration and Neutrophil Transmigration Across the Blood-Brain Barrier are Modulated by Alpha7 Nicotinic Receptor. PLoS ONE 2011, 6, e25016. [Google Scholar] [CrossRef]
- Wang, S.F.; Peng, L.; Gai, Z.T.; Zhang, L.H.; Jong, A.; Cao, H.; Huang, S.H. Pathogenic Triad in Bacterial Meningitis: Pathogen Invasion, NF-κB Activation, and Leukocyte Transmigration that Occur at the Blood-Brain Barrier. Front. Microbiol. 2016, 7, 148. [Google Scholar] [CrossRef]
- Sitcheran, R.; Gupta, P.; Fisher, P.B.; Baldwin, A.S. Positive and negative regulation of EAAT2 by NF-κB:: A role for N-myc in TNFα-controlled repression. Embo J. 2005, 24, 510–520. [Google Scholar] [CrossRef]
- Ghosh, M.; Yang, Y.J.; Rothstein, J.D.; Robinson, M.B. Nuclear Factor-κB Contributes to Neuron-Dependent Induction of Glutamate Transporter-1 Expression in Astrocytes. J. Neurosci. 2011, 31, 9159–9169. [Google Scholar] [CrossRef]
- Swann, J.R.; Spitzer, S.O.; Diaz Heijtz, R. Developmental Signatures of Microbiota-Derived Metabolites in the Mouse Brain. Metabolites 2020, 10, 172. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Mao, Q.; Xie, J.; Wang, Y.; Shao, W.H.; Zhong, Q.; Chen, J.J. Multi-omics data reveals the disturbance of glycerophospholipid metabolism caused by disordered gut microbiota in depressed mice. J. Adv. Res. 2022, 39, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Santamaría, A.; Ríos, C.; Solís-Hernández, F.; Ordaz-Moreno, J.; González-Reynoso, L.; Altagracia, M.; Kravzov, J. Systemic DL-kynurenine and probenecid pretreatment attenuates quinolinic acid-induced neurotoxicity in rats. Neuropharmacology 1996, 35, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Clifford, D.B.; Ances, B.M. HIV-associated neurocognitive disorder. Lancet Infect. Dis. 2013, 13, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Martin-Blondel, G.; Delobel, P.; Blancher, A.; Massip, P.; Marchou, B.; Liblau, R.S.; Mars, L.T. Pathogenesis of the immune reconstitution inflammatory syndrome affecting the central nervous system in patients infected with HIV. Brain 2011, 134, 928–946. [Google Scholar] [CrossRef] [PubMed]
- Paul, R. Neurocognitive Phenotyping of HIV in the Era of Antiretroviral Therapy. Curr. Hiv-Aids Rep. 2019, 16, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Keegan, M.R.; Chittiprol, S.; Letendre, S.L.; Winston, A.; Fuchs, D.; Boasso, A.; Iudicello, J.; Ellis, R.J. Tryptophan Metabolism and Its Relationship with Depression and Cognitive Impairment Among HIV-infected Individuals. Int. J. Tryptophan Res. 2016, 9, 79–88. [Google Scholar] [CrossRef]
- Qi, Q.; Hua, S.; Clish, C.B.; Scott, J.M.; Hanna, D.B.; Wang, T.; Haberlen, S.A.; Shah, S.J.; Glesby, M.J.; Lazar, J.M.; et al. Plasma Tryptophan-Kynurenine Metabolites Are Altered in Human Immunodeficiency Virus Infection and Associated with Progression of Carotid Artery Atherosclerosis. Clin. Infect. Dis. 2018, 67, 235–242. [Google Scholar] [CrossRef]
- Rivero-Buceta, E.; Doyagüez, E.G.; Colomer, I.; Quesada, E.; Mathys, L.; Noppen, S.; Liekens, S.; Camarasa, M.J.; Pérez-Pérez, M.J.; Balzarini, J.; et al. Tryptophan dendrimers that inhibit HIV replication, prevent virus entry and bind to the HIV envelope glycoproteins gp120 and gp41. Eur. J. Med. Chem. 2015, 106, 34–43. [Google Scholar] [CrossRef]
- Tóth, F.; Cseh, E.K.; Vécsei, L. Natural Molecules and Neuroprotection: Kynurenic Acid, Pantethine and α-Lipoic Acid. Int. J. Mol. Sci. 2021, 22, 403. [Google Scholar] [CrossRef]
- Martos, D.; Tuka, B.; Tanaka, M.; Vécsei, L.; Telegdy, G. Memory Enhancement with Kynurenic Acid and Its Mechanisms in Neurotransmission. Biomedicines 2022, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Kerr, S.J.; Armati, P.J.; Pemberton, L.A.; Smythe, G.; Tattam, B.; Brew, B.J. Kynurenine pathway inhibition reduces neurotoxicity of HIV-1-infected macrophages. Neurology 1997, 49, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, S.; Olsson, S.K.; Engberg, G. Pharmacological manipulation of kynurenic acid: Potential in the treatment of psychiatric disorders. CNS Drugs 2009, 23, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Okuno, A.; Fukuwatari, T.; Shibata, K. High tryptophan diet reduces extracellular dopamine release via kynurenic acid production in rat striatum. J. Neurochem. 2011, 118, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Fukui, S.; Schwarcz, R.; Rapoport, S.I.; Takada, Y.; Smith, Q.R. Blood-brain barrier transport of kynurenines: Implications for brain synthesis and metabolism. J. Neurochem. 1991, 56, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Turska, M.; Paluszkiewicz, P.; Turski, W.A.; Parada-Turska, J. A Review of the Health Benefits of Food Enriched with Kynurenic Acid. Nutrients 2022, 14, 4182. [Google Scholar] [CrossRef] [PubMed]
- Németh, H.; Robotka, H.; Kis, Z.; Rózsa, E.; Janáky, T.; Somlai, C.; Marosi, M.; Farkas, T.; Toldi, J.; Vécsei, L. Kynurenine administered together with probenecid markedly inhibits pentylenetetrazol-induced seizures. An electrophysiological and behavioural study. Neuropharmacology 2004, 47, 916–925. [Google Scholar] [CrossRef]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Brandebura, A.N.; Paumier, A.; Onur, T.S.; Allen, N.J. Astrocyte contribution to dysfunction, risk and progression in neurodegenerative disorders. Nat. Rev. Neurosci. 2023, 24, 23–39. [Google Scholar] [CrossRef]
- Zamanian, J.L.; Xu, L.J.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic Analysis of Reactive Astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef]
- Lei, W.; Zhao, C.; Sun, J.; Jin, Y.; Duan, Z. Activation of α7nAChR preserves intestinal barrier integrity by enhancing the HO-1/STAT3 signaling to inhibit NF-κB activation in mice. Biomed. Pharmacother. 2022, 149, 112733. [Google Scholar] [CrossRef] [PubMed]
- Galle-Treger, L.; Suzuki, Y.; Patel, N.; Sankaranarayanan, I.; Aron, J.L.; Maazi, H.; Chen, L.; Akbari, O. Nicotinic acetylcholine receptor agonist attenuates ILC2-dependent airway hyperreactivity. Nat. Commun. 2016, 7, 13202. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Barres, B. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef]
- Carrero, I.; Gonzalo, M.R.; Martin, B.; Sanz-Anquela, J.M.; Arévalo-Serrano, J.; Gonzalo-Ruiz, A. Oligomers of beta-amyloid protein (Aβ1-42) induce the activation of cyclooxygenase-2 in astrocytes via an interaction with interleukin-1beta, tumour necrosis factor-alpha, and a nuclear factor kappa-B mechanism in the rat brain. Exp. Neurol. 2012, 236, 215–227. [Google Scholar] [CrossRef]
- Marszalek-Grabska, M.; Walczak, K.; Gawel, K.; Wicha-Komsta, K.; Wnorowska, S.; Wnorowski, A.; Turski, W.A. Kynurenine emerges from the shadows-Current knowledge on its fate and function. Pharmacol. Therapeut. 2021, 225, 107845. [Google Scholar] [CrossRef]
- Wolf, H.; Brown, R.R.; Arend, R.A. The kynurenine load test, an adjunct to the tryptophan load test. Scand. J. Clin. Lab. Investig. 1980, 40, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Vámos, E.; Párdutz, A.; Varga, H.; Bohár, Z.; Tajti, J.; Fülöp, F.; Toldi, J.; Vécsei, L. l-kynurenine combined with probenecid and the novel synthetic kynurenic acid derivative attenuate nitroglycerin-induced nNOS in the rat caudal trigeminal nucleus. Neuropharmacology 2009, 57, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Silva-Adaya, D.; Pérez-De La Cruz, V.; Villeda-Hernández, J.; Carrillo-Mora, P.; González-Herrera, I.G.; García, E.; Colín-Barenque, L.; Pedraza-Chaverrí, J.; Santamaría, A. Protective effect of L-kynurenine and probenecid on 6-hydroxydopamine-induced striatal toxicity in rats: Implications of modulating kynurenate as a protective strategy. Neurotoxicol. Teratol. 2011, 33, 303–312. [Google Scholar] [CrossRef]
- Mok, M.H.S.; Fricker, A.C.; Weil, A.; Kew, J.N.C. Electrophysiological characterisation of the actions of kynurenic acid at ligand-gated ion channels. Neuropharmacology 2009, 57, 242–249. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Zou, J.; Huang, P.; Gao, X.; Lun, J.; Li, Y.; Gong, Z.; Cao, H. KYNA Ameliorates Glutamate Toxicity of HAND by Enhancing Glutamate Uptake in A2 Astrocytes. Int. J. Mol. Sci. 2024, 25, 4286. https://doi.org/10.3390/ijms25084286
Chen J, Zou J, Huang P, Gao X, Lun J, Li Y, Gong Z, Cao H. KYNA Ameliorates Glutamate Toxicity of HAND by Enhancing Glutamate Uptake in A2 Astrocytes. International Journal of Molecular Sciences. 2024; 25(8):4286. https://doi.org/10.3390/ijms25084286
Chicago/Turabian StyleChen, Jie, Jinhu Zou, Pengwei Huang, Xuefeng Gao, Jingxian Lun, Yubin Li, Zelong Gong, and Hong Cao. 2024. "KYNA Ameliorates Glutamate Toxicity of HAND by Enhancing Glutamate Uptake in A2 Astrocytes" International Journal of Molecular Sciences 25, no. 8: 4286. https://doi.org/10.3390/ijms25084286