
The Neurotranscriptome of Monochamus alternatus
 ,
,
Abstract
:1. Introduction
2. Results
2.1. The Analysis of the Spatiotemporal Expression Levels of Hte CSP Genes of M. alternatus
2.2. The Analysis of the Spatiotemporal Expression Levels of the SNMP Genes of M. alternatus
2.3. The Analysis of the Spatiotemporal Expression Levels of the OBP Genes of M. alternatus
2.4. The Analysis of the Spatiotemporal Expression Levels of the OR Genes of M. alternatus
2.5. The Analysis of the Spatiotemporal Expression Levels of the GR Genes of M. alternatus
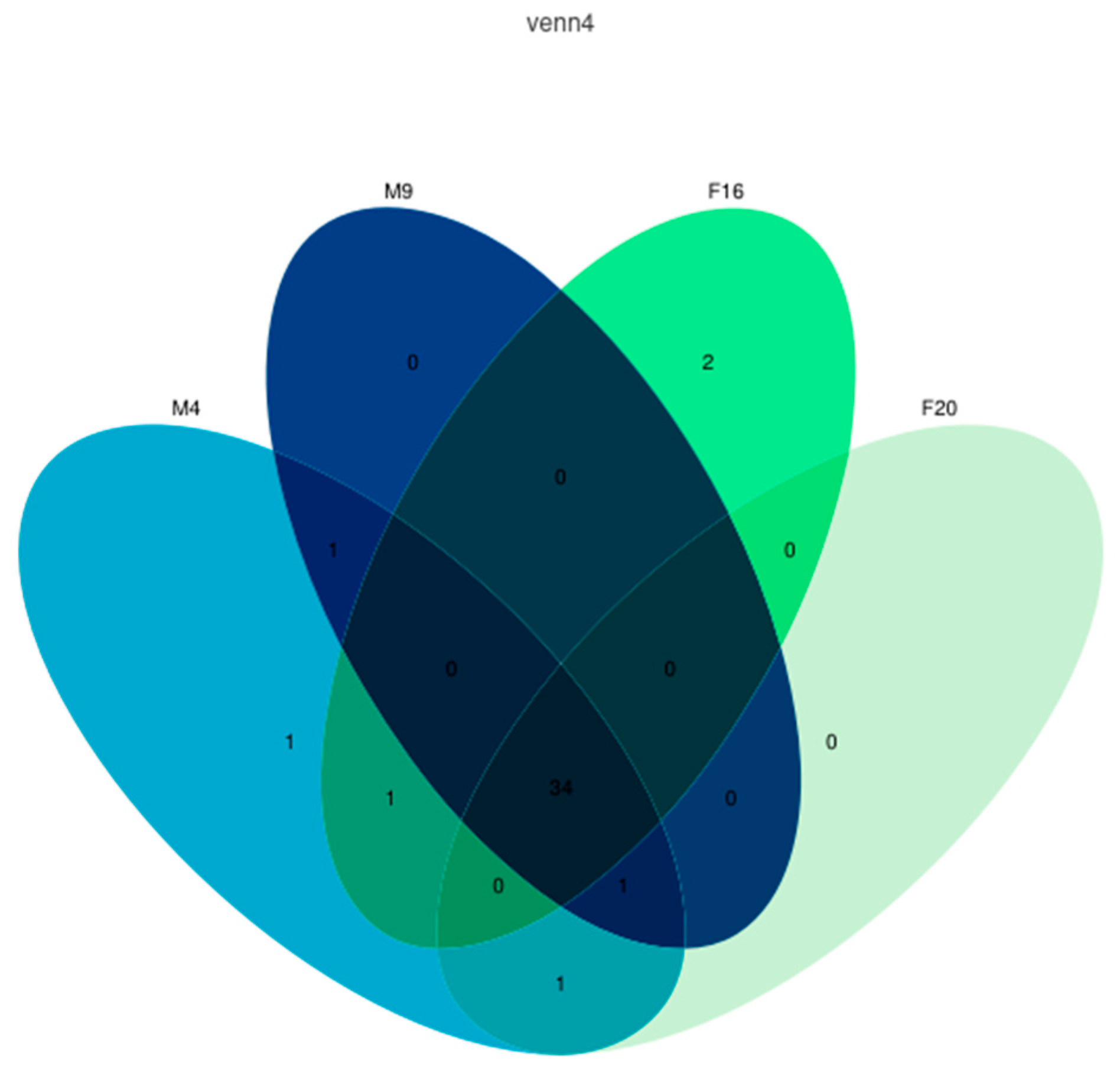
2.6. The Analysis of the Coexpression of the IR Genes of M. alternatus
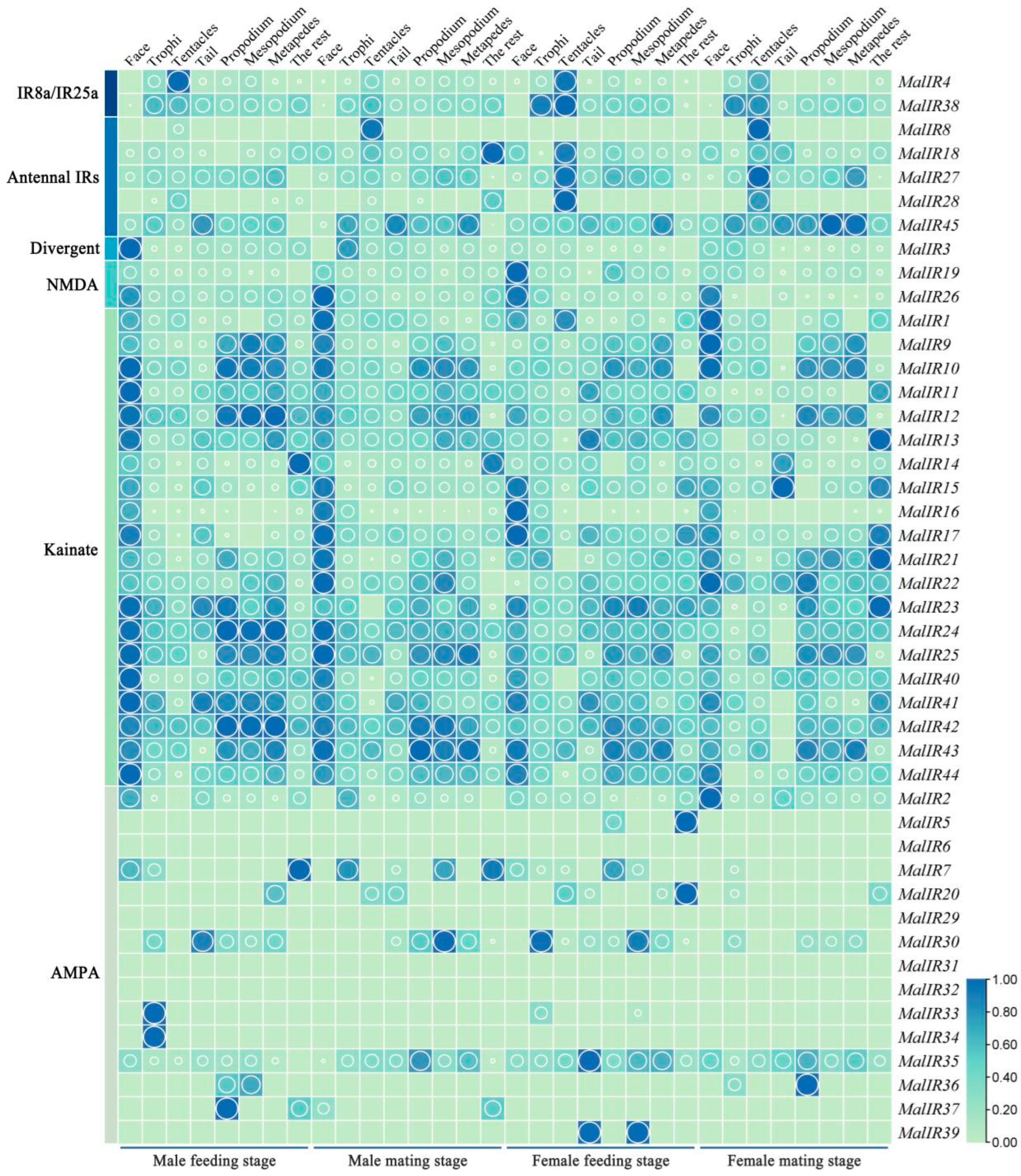
2.7. The Analysis of the Spatiotemporal Expression Levels of the IR Genes of M. alternatus
3. Discussion
4. Materials and Methods
4.1. The Test Insect
4.2. RNA Isolation and Illumina Sequencing
4.3. De Novo Transcriptome Assembly and Annotation
4.4. Differential Expression Analysis of Chemosensory Receptor Protein Genes
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Linit, M.J. Nemtaode-vector relationships in the pine wilt disease system. J. Nematol. 1988, 20, 227. [Google Scholar] [PubMed]
- Togashi, K.; Arakawa, Y. Horizontal transmission of Bursaphelenchus xylophilus between sexes of Monochamus alternatus. J. Nematol. 2003, 35, 7–16. [Google Scholar] [PubMed]
- Han, X.; Li, Y.; Huang, W.; Wang, R.; Hu, X.; Liang, G.; Huang, S.; Lian, C.; Zhang, F.; Wu, S. Landscapes drive the dispersal of Monochamus alternatus, vector of the pinewood nematode, revealed by whole-genome resequencing. For. Ecol. Manag. 2023, 529, 120682. [Google Scholar] [CrossRef]
- Wen, X.; Hong, Y.; Zhong, J.; Li, L.; Ma, Q.; Hu, X.; Han, X.; Guo, W.; Huang, Y.; Zhang, F. Assessing the impact of pine wilt disease on aboveground carbon storage in planted Pinus massoniana Lamb. forests via remote sensing. Sci. Total Environ. 2024, 914, 169906. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhou, T.; Hu, X.; Zhu, Y.; Shi, Z.; Chen, S.; Liu, Y.; Weng, X.; Zhang, F.; Wu, S. Discovery and Characterization of MaK: A Novel Knottin Antimicrobial Peptide from Monochamus alternatus. Int. J. Mol. Sci. 2023, 24, 17565. [Google Scholar] [CrossRef]
- Gao, R.; Liu, L.; Zhao, L.; Cui, S. Potentially suitable geographical area for Monochamus alternatus under current and future climatic scenarios based on optimized maxent model. Insects 2023, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Dwinell, L. First report of pinewood nematode (Bursaphelenchus xylophilus) in Mexico. Plant Dis. 1993, 77, 846. [Google Scholar] [CrossRef]
- Hunt, D. Pine wilt disease: A worldwide threat to forest ecosystems. Nematology 2009, 11, 315–316. [Google Scholar] [CrossRef]
- Li, M.; Li, H.; Ding, X.; Wang, L.; Wang, X.; Chen, F. The detection of pine wilt disease: A literature review. Int. J. Mol. Sci. 2022, 23, 10797. [Google Scholar] [CrossRef]
- Maehara, N.; Kanzaki, N.; Aikawa, T.; Nakamura, K. Potential vector switching in the evolution of Bursaphelenchus xylophilus group nematodes (Nematoda: Aphelenchoididae). Ecol. Evol. 2020, 10, 14320–14329. [Google Scholar] [CrossRef]
- Shimazu, M. Use of microbes for control of Monochamus alternatus, vector of the invasive pinewood nematode. In Use of Microbes for Control Eradication of Invasive Arthropods; Springer: Berlin/Heidelberg, Germany, 2009; Volume 6, pp. 141–157. [Google Scholar] [CrossRef]
- Zhao, B.G. Pine wilt disease in China. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 18–25. [Google Scholar]
- Huiling, H. Application of insect chemical ecology on the prevention and control of public health pests. Chin. J. Vector Biol. Control 2018, 29, 313–316. [Google Scholar]
- Tao, M.A. Monitoring Monochamus alternatus population dynamics with APF-I attractant in yunnan. For. Pest Dis. 2016, 35, 21–23. [Google Scholar]
- Jianting, F.; Juncuo, M.; Baode, W.; Lilin, Z.; Jianghua, S. Field trapping the japanese pine sawyer Monochamus alternatus hope (Coleoptera: Cerambycidae) using an aggregation pheromone and host volatiles. Chin. J. Appl. Entomol. 2013, 50, 1274–1279. [Google Scholar]
- Dejun, H.; Binqi, F.; Jingen, T.; Yan, W.; Fenglin, M. Screening of attractants for Monochamus alternatus and its attraction effects. J. Northeast For. Univ. 2009, 37, 86–87. [Google Scholar]
- Long, C.; Qiang, L.; Junnan, L.; Baode, W.; Feiping, Z. Comparison of efficacies for Monochamus alternatus adult caught by panel traps baited with APF-I chemical attractant placed at different field positions. J. Fujian Coll. For. 2014, 34, 11–14. [Google Scholar]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Zhang, M.; Üçpunar, H.K.; Svensson, T.; Quillery, E.; Gompel, N.; Ignell, R.; Kadow, I.C.G. Ionotropic chemosensory receptors mediate the taste and smell of polyamines. PLoS Biol. 2016, 14, e1002454. [Google Scholar] [CrossRef]
- Koh, T.W.; He, Z.; Gorur-Shandilya, S.; Menuz, K.; Larter, N.K.; Stewart, S.; Carlson, J.R. The Drosophila IR20a clade of ionotropic receptors are candidate taste and pheromone receptors. Neuron 2014, 83, 850–865. [Google Scholar] [CrossRef] [PubMed]
- Herre, M.; Goldman, O.V.; Lu, T.C.; Caballero-Vidal, G.; Qi, Y.; Gilbert, Z.N.; Gong, Z.; Morita, T.; Rahiel, S.; Ghaninia, M.; et al. Non-canonical odor coding in the mosquito. Cell 2022, 185, 3104–3123.e28. [Google Scholar] [CrossRef]
- Iovinella, I.; Bozza, F.; Caputo, B.; Della Torre, A.; Pelosi, P. Ligand-binding study of Anopheles gambiae chemosensory proteins. Chem. Senses 2013, 38, 409–419. [Google Scholar] [CrossRef]
- Ban, L.; Zhang, L.; Yan, Y.; Pelosi, P. Binding properties of a locust’s chemosensory protein. Biochem. Biophys. Res. Commun. 2002, 293, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; He, Y.Y.; Zhang, Y.J.; Guo, Z.J.; Xie, W.; Wu, Q.-J.; Wang, S.-J. Characterization of the chemosensory protein EforCSP3 and its potential involvement in host location by Encarsia formosa. J. Integr. Agric. 2023, 22, 514–525. [Google Scholar]
- Ishimaru, Y.; Inada, H.; Kubota, M.; Zhuang, H.; Tominaga, M.; Matsunami, H. Transient receptor potential family members PKD1L3 and PKD2L1 form a candidate sour taste receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 12569–12574. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; Zhang, S.; Luo, J.Y.; Zhu, J.; Cui, J.J.; Dong, S.L. Expression analysis and binding assays in the chemosensory protein gene family indicate multiple roles in Helicoverpa armigera. J. Chem. Ecol. 2015, 41, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Sugaya, S.; Yasukawa, J.; Aigaki, T.; Fuyama, Y. Odorant-binding proteins OBP57d and OBP57e affect taste perception and host-plant preference in Drosophila sechellia. PLoS Biol. 2007, 5, e118. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Sun, X.; Wang, M.Q. Expression pattern and ligand-binding properties of odorant-binding protein 13 from Monochamus alternatus hope. J. Appl. Entomol. 2017, 141, 751–757. [Google Scholar] [CrossRef]
- Zhu, J.; Iovinella, I.; Dani, F.R.; Liu, Y.L.; Huang, L.Q.; Liu, Y.; Wang, C.Z.; Pelosi, P.; Wang, G. Conserved chemosensory proteins in the proboscis and eyes of Lepidoptera. Int. J. Biol. Sci. 2016, 12, 1394–1404. [Google Scholar] [CrossRef]
- Chang, X.; Bi, Y.; Chi, H.; Fang, Q.; Lu, Z.; Wang, F.; Ye, G. Identification and expressionanalysis of odorant-binding and chemosensory protein genes in virus vector Nephotettix cincticeps. Insects 2022, 13, 1024. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Guo, H.; Huang, L.Q.; Pelosi, P.; Wang, C.Z. Unique function of a chemosensory protein in the proboscis of two Helicoverpa species. J. Exp. Biol. 2014, 217, 1821–1826. [Google Scholar] [CrossRef]
- Liu, Y.P.; Cui, Z.Y.; Si, P.F.; Liu, Y.; Zhou, Q.; Wang, G. Characterization of a specific odorant receptor for linalool in the Chinese citrus fly Bactrocera minax (Diptera: Tephritidae). Insect Biochem. 2020, 122, 103389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Anderson, A.R.; Trowell, S.C.; Luo, A.R.; Xiang, Z.H.; Xia, Q.Y. Topological and functional characterization of an insect gustatory receptor. PLoS ONE 2011, 6, e24111. [Google Scholar]
- Lee, Y.S.; Moon, S.J.; Montell, C. Multiple gustatory receptors required for the caffeine response in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 4495–4500. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Hall, J. Conditioned courtship in Drosophila and its mediation by association of chemical cues. Proc. Natl. Acad. Sci. USA 1979, 76, 3430–3434. [Google Scholar] [CrossRef] [PubMed]
- Abuin, L.; Bargeton, B.; Ulbrich, M.H.; Isacoff, E.Y.; Kellenberger, S.; Benton, R. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 2011, 69, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.Y.; Xu, W.; Dong, S.L.; Zhu, J.Y.; Xu, Y.X.; Anderson, A. Genome-wide analysis of ionotropic receptor gene repertoire in Lepidoptera with an emphasis on its functions of Helicoverpa armigera. Insect Biochem. Mol. Biol. 2018, 99, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Huang, X.Q.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.D.; White, J.; Cheung, F.; Parvizi, B. TIGR gene indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Li, L.T.; Zhu, Y.B.; Ma, J.F.; Li, Z.Y.; Dong, Z.P. An Analysis of the Athetis lepigone transcriptome from four developmental stages. PLoS ONE 2013, 8, e73911. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Pimentel, H. What the FPKM? A review of RNA-Seq expression units. The Farrago. Available online: https://haroldpimentel.wordpress.com/2014/05/08/what-the-fpkm-a-review-rna-seq-expression-units/ (accessed on 7 January 2024).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gender | Instar | Sampling Location |
|---|---|---|
| Male | 4 d post-eclosion (feeding stage) | Face Trophi Tentacles Tail Propodium Mesopodium Metapedes The rest |
| 9 d post-eclosion (mating stage) | ||
| Female | 16 d post-eclosion (feeding stage) | |
| 20 d post-eclosion (mating stage) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Weng, M.; Shi, W.; Wen, Y.; Long, Y.; Hu, X.; Ji, G.; Zhu, Y.; Wen, X.; Zhang, F.; et al. The Neurotranscriptome of Monochamus alternatus. Int. J. Mol. Sci. 2024, 25, 4553. https://doi.org/10.3390/ijms25084553
Han X, Weng M, Shi W, Wen Y, Long Y, Hu X, Ji G, Zhu Y, Wen X, Zhang F, et al. The Neurotranscriptome of Monochamus alternatus. International Journal of Molecular Sciences. 2024; 25(8):4553. https://doi.org/10.3390/ijms25084553
Chicago/Turabian StyleHan, Xiaohong, Mingqing Weng, Wenchao Shi, Yingxin Wen, Yirong Long, Xinran Hu, Guoxi Ji, Yukun Zhu, Xuanye Wen, Feiping Zhang, and et al. 2024. "The Neurotranscriptome of Monochamus alternatus" International Journal of Molecular Sciences 25, no. 8: 4553. https://doi.org/10.3390/ijms25084553
APA StyleHan, X., Weng, M., Shi, W., Wen, Y., Long, Y., Hu, X., Ji, G., Zhu, Y., Wen, X., Zhang, F., & Wu, S. (2024). The Neurotranscriptome of Monochamus alternatus. International Journal of Molecular Sciences, 25(8), 4553. https://doi.org/10.3390/ijms25084553