Vaccines and the Eye: Current Understanding of the Molecular and Immunological Effects of Vaccination on the Eye


Abstract
:1. Introduction
2. Methods of Literature Search
3. Molecular and Immunological Effects
3.1. Direct Inflammation Due to Live Attenuated Vaccines
3.2. Autoimmune/Inflammatory Syndrome Induced by Adjuvants (ASIA)
3.2.1. Inflammation Caused by Adjuvant and Adaptive Immune Component Interactions
3.2.2. Aluminum Particles That Delay Vaccine Dissolution Are Captured by Immune Cells
3.2.3. Aluminum-Related Dorsal Root Ganglion Injury
3.2.4. AS01B-Induced Ocular Inflammation
3.3. Type III Hypersensitivity/Immune Complex Disease and Type IV Hypersensitivity/Delayed-Type Hypersensitivity (DTH)
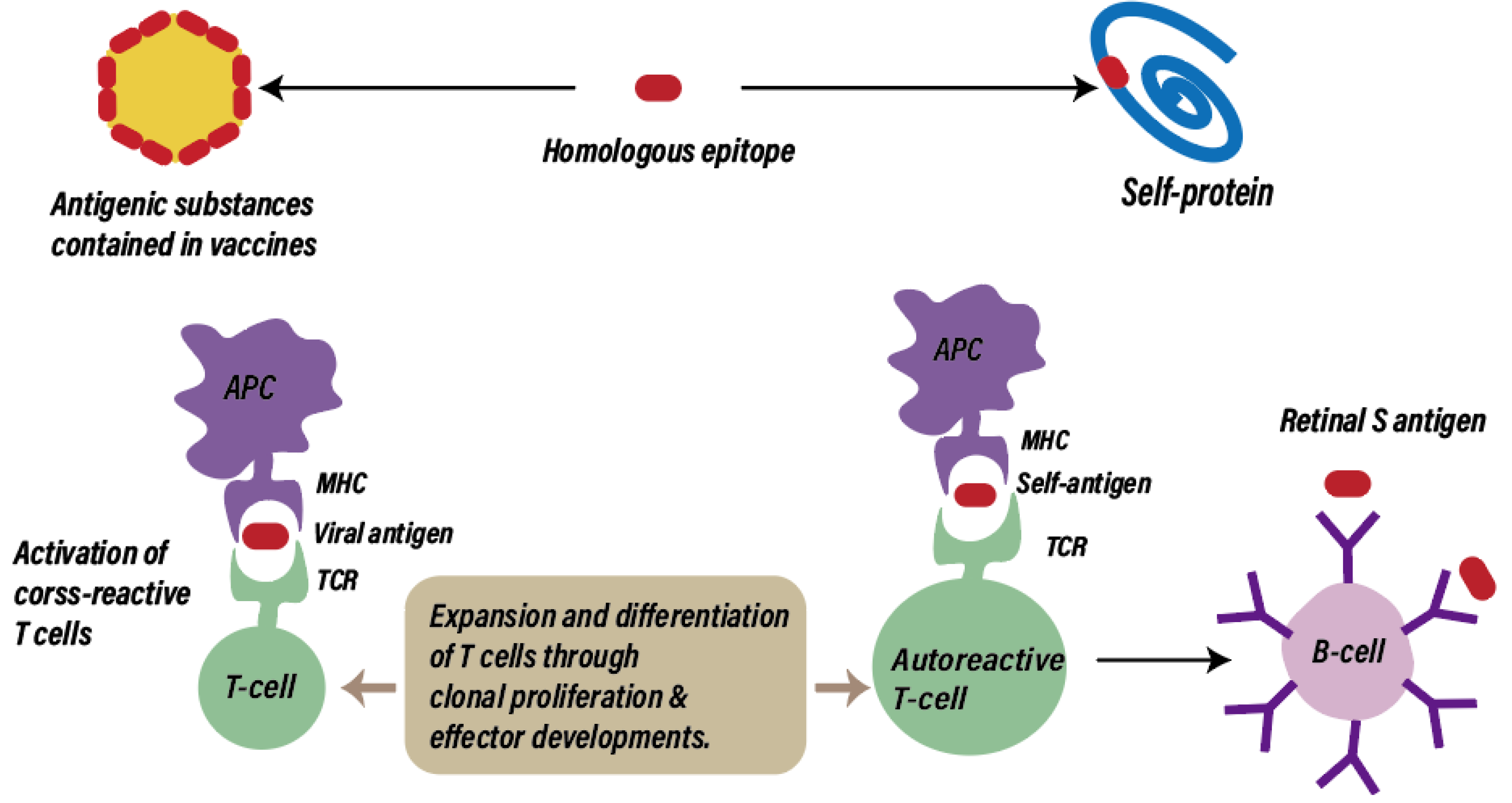
3.4. Molecular Mimicry
3.4.1. Acute Posterior Multifocal Lamellar Pigment (APMPPE) and Molecular Mimicry
3.4.2. Tubulointerstitial Nephritis and Uveitis (TINU) Syndrome and Molecular Mimicry
3.5. PEG-Induced Allergic Reactions
3.6. Type 1 IFN Activation and the Inflammatory Response of Retinal Pigment Epithelial (RPE) Cells
3.7. Free Extracellular RNA
3.8. Specific Components
Thimerosal
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Watad, A.; Bragazzi, N.L.; McGonagle, D.; Adawi, M.; Bridgewood, C.; Damiani, G.; Alijotas-Reig, J.; Esteve-Valverde, E.; Quaresma, M.; Amital, H.; et al. Autoimmune/inflammatory syndrome induced by adjuvants (ASIA) demonstrates distinct autoimmune and autoinflammatory disease associations according to the adjuvant subtype: Insights from an analysis of 500 cases. Clin. Immunol. 2019, 203, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tervaert, J.W.C.; Martinez-Lavin, M.; Jara, L.J.; Halpert, G.; Watad, A.; Amital, H.; Shoenfeld, Y. Autoimmune/inflammatory syndrome induced by adjuvants (ASIA) in 2023. Autoimmun. Rev. 2023, 22, 103287. [Google Scholar] [CrossRef] [PubMed]
- Vera-Lastra, O.; Medina, G.; Cruz-Dominguez, M.d.P.; Ramirez, P.; Gayosso-Rivera, J.A.; Anduaga-Dominguez, H.; Lievana-Torres, C.; Jara, L.J. Human adjuvant disease induced by foreign substances: A new model of ASIA (Shoenfeld’s syndrome). Lupus 2012, 21, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Bragazzi, N.L.; Watad, A.; Amital, H.; Shoenfeld, Y. Debate on vaccines and autoimmunity: Do not attack the author, yet discuss it methodologically. Vaccine 2017, 35, 5522–5526. [Google Scholar] [CrossRef] [PubMed]
- Elisha, E.; Guetzkow, J.; Shir-Raz, Y.; Ronel, N. Suppressing Scientific Discourse on Vaccines? Self-perceptions of researchers and practitioners. HEC Forum 2024, 36, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Salmon, D.A.; Proschan, M.; Forshee, R.; Gargiullo, P.; Bleser, W.; Burwen, D.R.; Cunningham, F.; Garman, P.; Greene, S.K.; Lee, G.M.; et al. Association between Guillain-Barré syndrome and influenza A (H1N1) 2009 monovalent inactivated vaccines in the USA: A meta-analysis. Lancet 2013, 381, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Miravalle, A.; Biller, J.; Schnitzler, E.; Bonwit, A. Neurological complications following vaccinations. Neurol. Res. 2010, 32, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Cecinati, V.; Principi, N.; Brescia, L.; Giordano, P.; Esposito, S. Vaccine administration and the development of immune thrombocytopenic purpura in children. Hum. Vaccines Immunother. 2013, 9, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Benage, M.; Fraunfelder, F.W. Vaccine-Associated Uveitis. Mo. Med. 2016, 113, 48–52. [Google Scholar] [PubMed]
- Agarwal, M.; Dutta Majumder, P.; Babu, K.; Konana, V.K.; Goyal, M.; Touhami, S.; Stanescu-Segall, D.; Bodaghi, B. Drug-induced uveitis: A review. Indian J. Ophthalmol. 2020, 68, 1799–1807. [Google Scholar] [CrossRef]
- Cheng, J.Y.; Margo, C.E. Ocular adverse events following vaccination: Overview and update. Surv. Ophthalmol. 2022, 67, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Kamoi, K.; Zong, Y.; Zhang, J.; Yang, M.; Ohno-Matsui, K. Ocular Inflammation Post-Vaccination. Vaccines 2023, 11, 1626. [Google Scholar] [CrossRef] [PubMed]
- Minor, P.D. Live attenuated vaccines: Historical successes and current challenges. Virology 2015, 479, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Oxman, M.N.; Levin, M.J.; Johnson, G.; Schmader, K.; Straus, S.; Gelb, L.; Arbeit, R.; Simberkoff, M.; Gershon, A.; Davis, L. A vaccine to prevent herpes zoster and postherpetic neuralgia in older adults. N. Engl. J. Med. 2005, 352, 2271–2284. [Google Scholar] [CrossRef] [PubMed]
- Grillo, A.; Fraunfelder, F.W. Keratitis in association with herpes zoster and varicella vaccines. Drugs Today 2017, 53, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Heath, G.; Depledge, D.P.; Brown, J.R.; Hale, A.D.; Tutil, H.; Williams, R.; Breuer, J. Acute Retinal Necrosis Caused by the Zoster Vaccine Virus. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 65, 2122–2125. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.A.; Levison, A.L.; Stewart, J.M.; Acharya, N.R.; Margolis, T.P. Retinal necrosis following varicella-zoster vaccination. Arch. Ophthalmol. 2012, 130, 1355–1356. [Google Scholar] [CrossRef] [PubMed]
- Ferrini, W.; Aubert, V.; Balmer, A.; Munier, F.L.; Abouzeid, H. Anterior uveitis and cataract after rubella vaccination: A case report of a 12-month-old girl. Pediatrics 2013, 132, e1035–e1038. [Google Scholar] [CrossRef] [PubMed]
- Jampol, L.M.; Sieving, P.A.; Pugh, D.; Fishman, G.A.; Gilbert, H. Multiple evanescent white dot syndrome. I. Clinical findings. Arch. Ophthalmol. 1984, 102, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Stangos, A.; Zaninetti, M.; Petropoulos, I.; Baglivo, E.; Pournaras, C. Multiple evanescent white dot syndrome following simultaneous hepatitis-A and yellow fever vaccination. Ocul. Immunol. Inflamm. 2006, 14, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Fine, L.; Fine, A.; Cunningham, E.T., Jr. Multiple evanescent white dot syndrome following hepatitis a vaccination. Arch. Ophthalmol. 2001, 119, 1856–1858. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Nazarian, S.M.; Thayi, D.R.; Hammond, F.; Petrovic, V. Multiple evanescent white dot syndrome following recent influenza vaccination. Can. J. Ophthalmol. 2013, 48, e115–e116. [Google Scholar] [CrossRef] [PubMed]
- Watad, A.; Quaresma, M.; Brown, S.; Cohen Tervaert, J.W.; Rodríguez-Pint, I.; Cervera, R.; Perricone, C.; Shoenfeld, Y. Autoimmune/inflammatory syndrome induced by adjuvants (Shoenfeld’s syndrome)—An update. Lupus 2017, 26, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.B.; O’Keefe, G.; Bui, D.; Jain, N. Vogt-Koyanagi-Harada Disease Associated with Hepatitis B Vaccination. Ocul. Immunol. Inflamm. 2019, 27, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Watad, A.; Quaresma, M.; Bragazzi, N.L.; Cervera, R.; Tervaert, J.W.C.; Amital, H.; Shoenfeld, Y. The autoimmune/inflammatory syndrome induced by adjuvants (ASIA)/Shoenfeld’s syndrome: Descriptive analysis of 300 patients from the international ASIA syndrome registry. Clin. Rheumatol. 2018, 37, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Awate, S.; Babiuk, L.A.; Mutwiri, G. Mechanisms of action of adjuvants. Front. Immunol. 2013, 4, 114. [Google Scholar] [CrossRef] [PubMed]
- Marciani, D.J. Vaccine adjuvants: Role and mechanisms of action in vaccine immunogenicity. Drug Discov. Today 2003, 8, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Koenig, H.C.; Sutherland, A.; Izurieta, H.S.; McGonagle, D. Application of the immunological disease continuum to study autoimmune and other inflammatory events after vaccination. Vaccine 2011, 29, 913–919. [Google Scholar] [CrossRef] [PubMed]
- McGonagle, D.; McDermott, M.F. A proposed classification of the immunological diseases. PLoS Med. 2006, 3, e297. [Google Scholar] [CrossRef] [PubMed]
- Fraunfelder, F.W.; Suhler, E.B.; Fraunfelder, F.T. Hepatitis B vaccine and uveitis: An emerging hypothesis suggested by review of 32 case reports. Cutan. Ocul. Toxicol. 2010, 29, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.M.; El-Sheikh, H.F.; Tabbara, K.F. Anterior uveitis following combined vaccination for measles, mumps and rubella (MMR): A report of two cases. Acta Ophthalmol. Scand. 2000, 78, 590–592. [Google Scholar] [CrossRef] [PubMed]
- Menge, T.; Cree, B.; Saleh, A.; Waterboer, T.; Berthele, A.; Kalluri, S.R.; Hemmer, B.; Aktas, O.; Hartung, H.-P.; Methner, A. Neuromyelitis optica following human papillomavirus vaccination. Neurology 2012, 79, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Lee, H.L.; Yeo, M.; Kim, J.S.; Shin, D.I.; Lee, S.S.; Lee, S.H. Recurrent optic neuritis and neuromyelitis optica-IgG following first and second human papillomavirus vaccinations. Clin. Neurol. Neurosurg. 2016, 144, 126–128. [Google Scholar] [CrossRef] [PubMed]
- Heydari-Kamjani, M.; Vante, I.; Uppal, P.; Demory Beckler, M.; Kesselman, M.M. Uveitis Sarcoidosis Presumably Initiated after Administration of Shing rix Vaccine. Cureus 2019, 11, e4920. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, S.; Shekhawat, N.S.; Chen, A.; Woreta, F.A. Presumed Herpes Zoster Ophthalmicus Reactivation Following Recombinant Zoster Vaccination. Cornea 2021, 40, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.J.; Wingelaar, M.J.; Armbrust, K.R.; Kopplin, L.J. Uveitis reactivation following recombinant zoster vaccination. Am. J. Ophthalmol. Case Rep. 2021, 23, 101115. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.I.; Deaner, J.D.; Srivastava, S.K.; Lowder, C.Y. Acute retinal necrosis following recombinant subunit varicella-zoster virus vaccine. Am. J. Ophthalmol. Case Rep. 2020, 20, 100962. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, A.; Matoba, A. Reactivation of Herpes Zoster Stromal Keratitis After HZ/su Adjuvanted Herpes Zoster Subunit Vaccine. Ophthalmology 2018, 125, 1682. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Sethi, H.S.; Rai, H.K.; Menon, V. Bilateral neuro-retinitis following chick embryo cell anti-rabies vaccination—A case report. BMC Ophthalmol. 2005, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, V.L.; Acheson, J.F.; Ball, J.; Plant, G.T. Optic neuritis following measles/rubella vaccination in two 13-year-old children. Br. J. Ophthalmol. 1996, 80, 1110–1111. [Google Scholar] [CrossRef] [PubMed]
- Fried, M.; Conen, D.; Conzelmann, M.; Steinemann, E. Uveitis after hepatitis B vaccination. Lancet 1987, 2, 631–632. [Google Scholar] [CrossRef]
- Sawai, T.; Shimizu, M.; Sakai, T.; Yachie, A. Tubulointerstitial Nephritis and Uveitis Syndrome Associated with Human Papillomavirus Vaccine. J. Pediatr. Ophthalmol. Strabismus 2016, 53, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Bandyopadhyay, S.; Bapuraj, J.R.; Gupta, A. Bilateral Optic Neuritis Complicating Rabies Vaccination. Retina 2004, 24, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Chen, C.L.; Hu, Y.Z.; Zeng, R. Multiple evanescent white dot syndrome following rabies vaccination: A case report. BMC Ophthalmol. 2018, 18, 312. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Chang, Y.-H.; Lee, Y.-C. Panuveitis following administration of quadrivalent human papillomavirus vaccine. Tzu Chi Med. J. 2014, 26, 44–46. [Google Scholar] [CrossRef]
- Khalifa, Y.M.; Monahan, P.M.; Acharya, N.R. Ampiginous choroiditis following quadrivalent human papilloma virus vaccine. Br. J. Ophthalmol. 2010, 94, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, L.S.; Montgomery, J.R.; Baker, K.M.; Colyer, M.H. Acute posterior multifocal placoid pigment epitheliopathy after immunization with multiple vaccines. Retin. Cases Brief Rep. 2022, 16, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-H.; Stone, C.A.; Jakubovic, B.; Phillips, E.J.; Sussman, G.; Park, J.; Hoang, U.; Kirshner, S.L.; Levin, R.; Kozlowski, S. Anti-PEG IgE in anaphylaxis associated with polyethylene glycol. J. Allergy Clin. Immunol. Pract. 2021, 9, 1731–1733.e1733. [Google Scholar] [CrossRef]
- Yasuda, E.; Matsumiya, W.; Maeda, Y.; Kusuhara, S.; Nguyen, Q.D.; Nakamura, M.; Hara, R. Multiple evanescent white dot syndrome following BNT162b2 mRNA COVID-19 vaccination. Am. J. Ophthalmol. Case Rep. 2022, 26, 101532. [Google Scholar] [CrossRef]
- Fowler, N.; Mendez Martinez, N.R.; Pallares, B.V.; Maldonado, R.S. Acute-onset central serous retinopathy after immunization with COVID-19 mRNA vaccine. Am. J. Ophthalmol. Case Rep. 2021, 23, 101136. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, A.; Beiran, I.; Krasnitz, I.; Miller, B. Bilateral optic neuritis after inactivated influenza vaccination. Isr. Med. Assoc. J. IMAJ 2012, 14, 705–707. [Google Scholar] [PubMed]
- Gherardi, R.K.; Crépeaux, G.; Authier, F.-J. Myalgia and chronic fatigue syndrome following immunization: Macrophag ic myofasciitis and animal studies support linkage to aluminum adjuvan t persistency and diffusion in the immune system. Autoimmun. Rev. 2019, 18, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Verdier, F.; Burnett, R.; Michelet-Habchi, C.; Moretto, P.; Fievet-Groyne, F.; Sauzeat, E. Aluminium assay and evaluation of the local reaction at several time points after intramuscular administration of aluminium containing vaccines in the Cynomolgus monkey. Vaccine 2005, 23, 1359–1367. [Google Scholar] [CrossRef]
- Authier, F.-J.; Sauvat, S.; Christov, C.; Chariot, P.; Raisbeck, G.; Poron, M.-F.; Yiou, F.; Gherardi, R. Al(OH)3-adjuvanted vaccine-induced macrophagic myofasciitis in rats is influenced by the genetic background. Neuromuscul. Disord. 2006, 16, 347–352. [Google Scholar] [CrossRef]
- Priest, N.; Newton, D.; Day, J.; Talbot, R.; Warner, A. Human metabolism of aluminium-26 and gallium-67 injected as citrates. Hum. Exp. Toxicol. 1995, 14, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Flarend, R.E.; Hem, S.L.; White, J.L.; Elmore, D.; Suckow, M.A.; Rudy, A.C.; Dandashli, E.A. In vivo absorption of aluminium-containing vaccine adjuvants using 26Al. Vaccine 1997, 15, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Banstola, A.; Reynolds, J.N.J. The Sheep as a Large Animal Model for the Investigation and Treatment of Human Disorders. Biology 2022, 11, 1251. [Google Scholar] [CrossRef]
- González, J.M.; Figueras, L.; Ortega, M.E.; Lozano, M.; de Arcaute, M.R.; Royo, R.; Cebrián, L.M.; Ferrer, L.M.; Fariñas, F.; de Jalón, J.A.; et al. Possible adverse reactions in sheep after vaccination with inactivated BTV vaccines. Vet. Rec. 2010, 166, 757–758. [Google Scholar] [CrossRef] [PubMed]
- Gunasekaran, M.; Chatterjee, P.K.; Shih, A.; Imperato, G.H.; Addorisio, M.; Kumar, G.; Lee, A.; Graf, J.F.; Meyer, D.; Marino, M. Immunization elicits antigen-specific antibody sequestration in dorsal root ganglia sensory neurons. Front. Immunol. 2018, 9, 638. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.R.; Harding, B.L.; Hoffman, P.N.; Griffin, J.W.; Price, D.L.; Troncoso, J.C. Aluminum-induced neurofilamentous changes in cultured rat dorsal root ganglia explants. J. Neurosci. 1992, 12, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, Y.M.; Jacoby, R.M.; Margolis, T.P. Exacerbation of zoster interstitial keratitis after zoster vaccination in an adult. Arch. Ophthalmol. 2010, 128, 1079–1080. [Google Scholar] [CrossRef] [PubMed]
- Jastrzebski, A.; Brownstein, S.; Ziai, S.; Saleh, S.; Lam, K.; Jackson, W.B. Reactivation of herpes zoster keratitis with corneal perforation after zoster vaccination. Cornea 2017, 36, 740–742. [Google Scholar] [CrossRef]
- Lal, H.; Cunningham, A.L.; Godeaux, O.; Chlibek, R.; Diez-Domingo, J.; Hwang, S.J.; Levin, M.J.; McElhaney, J.E.; Poder, A.; Puig-Barberà, J.; et al. Efficacy of an adjuvanted herpes zoster subunit vaccine in older adults. N. Engl. J. Med. 2015, 372, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, T.; Ming, D.; Breuer, J. A critical appraisal of ‘Shingrix’, a novel herpes zoster subunit vaccine (HZ/Su or GSK1437173A) for varicella zoster virus. Hum. Vaccines Immunother. 2017, 13, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Tilton, R.G.; Chang, K.; Corbett, J.A.; Misko, T.P.; Currie, M.G.; Bora, N.S.; Kaplan, H.J.; Williamson, J.R. Endotoxin-induced uveitis in the rat is attenuated by inhibition of nitric oxide production. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3278–3288. [Google Scholar]
- Newman, M.J.; Wu, J.Y.; Gardner, B.H.; Anderson, C.A.; Kensil, C.R.; Recchia, J.; Coughlin, R.T.; Powell, M.F. Induction of cross-reactive cytotoxic T-lymphocyte responses specific for HIV-1 gp120 using saponin adjuvant (QS-21) supplemented subunit vaccine formulations. Vaccine 1997, 15, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Guy, B. The perfect mix: Recent progress in adjuvant research. Nat. Rev. Microbiol. 2007, 5, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Chlibek, R.; Bayas, J.M.; Collins, H.; de la Pinta, M.L.; Ledent, E.; Mols, J.F.; Heineman, T.C. Safety and immunogenicity of an AS01-adjuvanted varicella-zoster virus subunit candidate vaccine against herpes zoster in adults ≥ 50 years of age. J. Infect. Dis. 2013, 208, 1953–1961. [Google Scholar] [CrossRef]
- Chlibek, R.; Smetana, J.; Pauksens, K.; Rombo, L.; Van den Hoek, J.A.; Richardus, J.H.; Plassmann, G.; Schwarz, T.F.; Ledent, E.; Heineman, T.C. Safety and immunogenicity of three different formulations of an adjuvanted varicella-zoster virus subunit candidate vaccine in older adults: A phase II, randomized, controlled study. Vaccine 2014, 32, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Landrigan, P.J. Neurologic Disorders Following Live Measles-Virus Vaccination. JAMA 1973, 223, 1459. [Google Scholar] [CrossRef] [PubMed]
- Dyck, P.J.; Daube, J.; O’Brien, P.; Pineda, A.; Low, P.A.; Windebank, A.J.; Swanson, C. Plasma exchange in chronic inflammatory demyelinating polyradiculoneur opathy. N. Engl. J. Med. 1986, 314, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Behan, P.O.; Moore, M.J.; Lamarche, J.B. Acute Necrotizing Hemorrhagic Encephalopathy. Postgrad. Med. 1973, 54, 154–160. [Google Scholar] [CrossRef]
- Poser, C.M.; Alter, M.; Currier, R.D.; Hunter, S.E. Common Demyelinating and Degenerative Diseases and Extrapyramidal Disorders—Panel 4. Arch. Neurol. 1979, 36, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Reik, L. Disseminated vasculomyelinopathy: An immune complex disease. Ann. Neurol. 1980, 7, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Drachman, D.A.; Paterson, P.Y.; Berlin, B.S.; Roguska, J. Immunosuppression and the Guillain-Barré Syndrome. Arch. Neurol. 1970, 23, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Goust, J.M.; Chenais, F.O.; Carnes, J.E.; Hames, C.G.; Fudenberg, H.H.; Hogan, E.L. Abnormal T cell subpopulations and circulating immune complexes in the Guillain-Barré syndrome and multiple sclerosis. Neurology 1978, 28, 421. [Google Scholar] [CrossRef] [PubMed]
- Hodson, A.K.; Doughty, R.A.; Norman, M.E. Acute Encephalopathy, Streptococcal Infection, and Cryoglobulinemia. Arch. Neurol. 1978, 35, 43–44. [Google Scholar] [CrossRef] [PubMed]
- Sergent, J.S.; Lockshin, M.D.; Christian, C.L.; Gocke, D.J. Vasculitis with hepatitis B antigenemia: Long-term observation in nine patients. Medicine 1976, 55, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, A. Neurologic illness following post-exposure prophylaxis with purifiled chick embryo cell antirabies vaccine. J. Assoc. Physicians India 2001, 49, 927–928. [Google Scholar] [PubMed]
- Waisbren, B.A. Acquired autoimmunity after viral vaccination is caused by molecular mimicry and antigen complimentarity in the presence of an immunologic adjuvant and specific HLA patterns. Med. Hypotheses 2008, 70, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Fujinami, R.S.; Oldstone, M.B.A. Amino Acid Homology Between the Encephalitogenic Site of Myelin Basic Protein and Virus: Mechanism for Autoimmunity. Science 1985, 230, 1043–1045. [Google Scholar] [CrossRef] [PubMed]
- Wucherpfennig, K.W.; Strominger, J.L. Molecular mimicry in T cell-mediated autoimmunity: Viral peptides activate human T cell clones specific for myelin basic protein. Cell 1995, 80, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.C.; Root-Bernstein, R.S. An explanation of prevention and suppression of experimental allergic encephalomyelitis. Mol. Immunol. 1983, 20, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Yamakawa, T.; Sugita, M.; Kijima, M.; Ishioka, M.; Tanaka, S.; Ohno, S. The role of tumor necrosis factor-alpha in the induction of experimental autoimmune uveoretinitis in mice. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3884–3889. [Google Scholar]
- Yokochi, T.; Fujii, Y.; Nakashima, I.; Asai, J.; Kiuchi, M.; Kojima, K.; Kato, N. A murine model of experimental autoimmune lens-induced uveitis using Klebsiella O3 lipopolysaccharide as a potent immunological adjuvant. Int. J. Exp. Pathol. 1993, 74, 573–582. [Google Scholar] [PubMed]
- de Vos, A.F.; Klaren, V.N.; Kijlstra, A. Expression of multiple cytokines and IL-1RA in the uvea and retina during endotoxin-induced uveitis in the rat. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3873–3883. [Google Scholar]
- Charteris, D.G.; Lightman, S.L. In vivo lymphokine production in experimental autoimmune uveoretinitis. Immunology 1993, 78, 387–392. [Google Scholar] [PubMed]
- Merino, G.; Fujino, Y.; Hanashiro, R.K. Lipoteichoic acid as an inducer of acute uveitis in the rat. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1251–1256. [Google Scholar]
- Natale, C.; Giannini, T.; Lucchese, A.; Kanduc, D. Computer-assisted analysis of molecular mimicry between human papillomavirus 16 E7 oncoprotein and human protein sequences. Immunol. Cell Biol. 2000, 78, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Garip, A.; Diedrichs-Möhring, M.; Thurau, S.R.; Deeg, C.A.; Wildner, G. Uveitis in a patient treated with Bacille-Calmette-Guérin: Possible antigenic mimicry of mycobacterial and retinal antigens. Ophthalmology 2009, 116, 2457–2462.e2451–e2452. [Google Scholar] [CrossRef] [PubMed]
- Gass, J.D.M. Acute posterior multifocal placoid pigment epitheliopathy. Arch. Ophthalmol. 1968, 80, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Reichhart, M.D.; Patterson, M.C.; Borruat, F.-X. Chapter 37—Acute Posterior Multifocal Placoid Pigment Epitheliopathy. In Uncommon Causes of Stroke; Cambridge University Press: Cambridge, UK, 2018; pp. 275–289. [Google Scholar] [CrossRef]
- Escott, S.; Tarabishy, A.B.; Davidorf, F.H. Multifocal choroiditis following simultaneous hepatitis A, typhoid, and yellow fever vaccination. Clin. Ophthalmol. 2013, 7, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.D.; Folk, J.C.; Panknen, C.A.; Goeken, N.E. HLA-B7 and HLA-DR2 antigens and acute posterior multifocal placoid pigment epitheliopathy. Arch. Ophthalmol. 1990, 108, 698–700. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Garner, L.I.; Zvyagin, I.V.; Paley, M.A.; Komech, E.A.; Jude, K.M.; Zhao, X.; Fernandes, R.A.; Hassman, L.M.; Paley, G.L.; et al. Autoimmunity-associated T cell receptors recognize HLA-B*27-bound pept ides. Nature 2022, 612, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Baxter, K.R.; Opremcak, E.M. Panretinal acute multifocal placoid pigment epitheliopathy: A novel posterior uveitis syndrome with HLA-A3 and HLA-C7 association. J. Ophthalmic Inflamm. Infect. 2013, 3, 29. [Google Scholar] [CrossRef]
- Stepanov, A.; Feuermannová, A.; Studnička, J.; Hejsek, L.; Burova, M.; Jirásková, N.; Rozsíval, P. Acute posterior multifocal placoid pigment epitheliopathy-case report. Ceska A Slov. Oftalmol. Cas. Ceske Oftalmol. Spol. A Slov. Oftalmol. Spol. 2014, 70, 72–76. [Google Scholar]
- Amaro, D.; Carreño, E.; Steeples, L.R.; Oliveira-Ramos, F.; Marques-Neves, C.; Leal, I. Tubulointerstitial nephritis and uveitis (TINU) syndrome: A review. Br. J. Ophthalmol. 2020, 104, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Mackensen, F.; David, F.; Schwenger, V.; Smith, L.K.; Rajalingam, R.; Levinson, R.D.; Austin, C.R.; Houghton, D.; Martin, T.M.; Rosenbaum, J.T. HLA-DRB1* 0102 is associated with TINU syndrome and bilateral, sudden-onset anterior uveitis but not with interstitial nephritis alone. Br. J. Ophthalmol. 2011, 95, 971–975. [Google Scholar] [CrossRef]
- Reddy, A.K.; Hwang, Y.-S.; Mandelcorn, E.D.; Davis, J.L. HLA-DR, DQ class II DNA typing in pediatric panuveitis and tubulointer stitial nephritis and uveitis. Am. J. Ophthalmol. 2013, 157, 678–686.e2. [Google Scholar] [CrossRef] [PubMed]
- Clive, D.M.; Vanguri, V.K. The Syndrome of Tubulointerstitial Nephritis with Uveitis (TINU). Am. J. Kidney Dis. 2017, 72, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Suzuki, K.; Nakahata, T.; Tateyama, T.; Waga, S.; Ito, E. Repeat renal biopsy in a girl with tubulointerstitial nephritis and uveitis syndrome. Pediatr. Nephrol. 2001, 16, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Dummer, C.D.; Carpio, V.N.; Gonçalves, L.F.S.; Manfro, R.C.; Veronese, F.V. FOXP3+ regulatory T cells: From suppression of rejection to induction of renal allograft tolerance. Transpl. Immunol. 2012, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rytkönen, S.H.; Kulmala, P.; Autio-Harmainen, H.; Arikoski, P.; Endén, K.; Kataja, J.; Karttunen, T.; Nuutinen, M.; Jahnukainen, T. FOXP3+ T cells are present in kidney biopsy samples in children with tubulointerstitial nephritis and uveitis syndrome. Pediatr. Nephrol. 2018, 33, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.; Jirawuthiworavong, G.V.; Nguyen, E.V.; Awazu, M.; Levinson, R.D.; Gordon, L.K. Tubulointerstitial Nephritis and Uveitis Syndrome: A Case with an Autoimmune Reactivity Against Retinal and Renal Antigens. Ocul. Immunol. Inflamm. 2008, 16, 51–53. [Google Scholar] [CrossRef]
- Tan, Y.; Yu, F.; Qu, Z.; Su, T.; Xing, G.-Q.; Wu, L.-H.; Wang, F.-M.; Liu, G.; Yang, L.; Zhao, M.-H. Modified C-reactive protein might be a target autoantigen of TINU synd rome. Clin. J. Am. Soc. Nephrol. 2011, 6, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Su, T.; Chu, R.; Li, X.; Yang, L. Tubulointerstitial nephritis with uveitis in Chinese adults. Clin. J. Am. Soc. Nephrol. 2014, 9, 21–28. [Google Scholar] [CrossRef]
- Ljutić, D.; Glavina, M. Tubulointerstitial Nephritis with Uveitis Syndrome following Varicella Zoster Reactivation. Nephron 1995, 71, 485–486. [Google Scholar] [CrossRef] [PubMed]
- Mackensen, F.; Smith, J.R.; Rosenbaum, J.T. Enhanced recognition, treatment, and prognosis of tubulointerstitial n ephritis and uveitis syndrome. Ophthalmology 2007, 114, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, C.; Xu, X.-F.; Xu, W.; Liu, S.-W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Book, B.A.J.; Schmidt, B.; Foerster, A.M.H. Bilateral Acute Macular Neuroretinopathy after Vaccination against SARS-CoV-2. JAMA Ophthalmol. 2021, 139, e212471. [Google Scholar] [CrossRef] [PubMed]
- Forchette, L.; Sebastian, W.; Liu, T. A Comprehensive Review of COVID-19 Virology, Vaccines, Variants, and Therapeutics. Curr. Med. Sci. 2021, 41, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Ichhpujani, P.; Parmar, U.P.S.; Duggal, S.; Kumar, S. COVID-19 Vaccine-Associated Ocular Adverse Effects: An Overview. Vaccines 2022, 10, 1879. [Google Scholar] [CrossRef] [PubMed]
- WHO. Coronavirus (COVID-19) Dashboard WHO Coronavirus (COVID-19) Dashboard with Vaccination Data. Available online: https://covid19.who.int/ (accessed on 26 February 2024).
- Ng, X.L.; Betzler, B.K.; Testi, I.; Ho, S.L.; Tien, M.; Ngo, W.K.; Zierhut, M.; Chee, S.P.; Gupta, V.; Pavesio, C.E.; et al. Ocular Adverse Events After COVID-19 Vaccination. Ocul. Immunol. Inflamm. 2021, 29, 1216–1224. [Google Scholar] [CrossRef] [PubMed]
- Eleiwa, T.K.; Gaier, E.D.; Haseeb, A.; ElSheikh, R.H.; Sallam, A.B.; Elhusseiny, A.M. Adverse Ocular Events following COVID-19 Vaccination. Inflamm. Res. 2021, 70, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Bøhler, A.D.; Strøm, M.E.; Sandvig, K.U.; Moe, M.C.; Jørstad, Ø.K. Acute macular neuroretinopathy following COVID-19 vaccination. Eye 2022, 36, 644–645. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, T.; Ben-Arie-Weintrob, Y.; Hareuveni-Blum, T.; Shaer, B.; Vishnevskia-Dai, V.; Shulman, S.; Newman, H.; Biadsy, M.; Masarwa, D.; Fischer, N.; et al. UVEITIS AFTER THE BNT162b2 mRNA VACCINATION AGAINST SARS-CoV-2 INFECTION: A Possible Association. RETINA 2021, 41, 2462–2471. [Google Scholar] [CrossRef] [PubMed]
- Inagawa, S.; Onda, M.; Miyase, T.; Murase, S.; Murase, H.; Mochizuki, K.; Sakaguchi, H. Multiple evanescent white dot syndrome following vaccination for COVID-19: A case report. Medicine 2022, 101, e28582. [Google Scholar] [CrossRef]
- Goyal, M.; Murthy, S.I.; Annum, S. Bilateral Multifocal Choroiditis following COVID-19 Vaccination. Ocul. Immunol. Inflamm. 2021, 29, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Kamoi, K.; Ohno-Matsui, K. Long Vax in the Eye: Long Post-COVID Vaccination Syndrome Presenting with Frosted Branch Angiitis. Diseases 2024, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Hamad, I.; Hunter, A.C.; Szebeni, J.; Moghimi, S.M. Poly(ethylene glycol)s generate complement activation products in human serum through increased alternative pathway turnover and a MASP-2-dependent process. Mol. Immunol. 2008, 46, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Ramadan, E.; Elsadek, N.E.; Emam, S.E.; Shimizu, T.; Ando, H.; Ishima, Y.; Elgarhy, O.H.; Sarhan, H.A.; Hussein, A.K.; et al. Polyethylene glycol (PEG): The nature, immunogenicity, and role in the hypersensitivity of PEGylated products. J. Control. Release 2022, 351, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.M.; Fries, L.F. The role of complement in inflammation and phagocytosis. Immunol. Today 1991, 12, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Noris, M.; Remuzzi, G. Overview of Complement Activation and Regulation. Semin. Nephrol. 2013, 33, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Lai, S.K. Anti-PEG immunity: Emergence, characteristics, and unaddressed questions. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 655–677. [Google Scholar] [CrossRef]
- Hsieh, Y.-C.; Wang, H.-E.; Lin, W.-W.; Roffler, S.R.; Cheng, T.-C.; Su, Y.-C.; Li, J.-J.; Chen, C.-C.; Huang, C.-H.; Chen, B.-M.; et al. Pre-existing anti-polyethylene glycol antibody reduces the therapeutic efficacy and pharmacokinetics of PEGylated liposomes. Theranostics 2018, 8, 3164–3175. [Google Scholar] [CrossRef]
- Ganson, N.J.; Povsic, T.J.; Sullenger, B.A.; Alexander, J.H.; Zelenkofske, S.L.; Sailstad, J.M.; Rusconi, C.P.; Hershfield, M.S. Pre-existing anti–polyethylene glycol antibody linked to first-exposure allergic reactions to pegnivacogin, a PEGylated RNA aptamer. J. Allergy Clin. Immunol. 2016, 137, 1610–1613.e1617. [Google Scholar] [CrossRef]
- Yang, Q.; Jacobs, T.M.; McCallen, J.D.; Moore, D.T.; Huckaby, J.T.; Edelstein, J.N.; Lai, S.K. Analysis of Pre-existing IgG and IgM Antibodies against Polyethylene Glycol (PEG) in the General Population. Anal. Chem. 2016, 88, 11804–11812. [Google Scholar] [CrossRef]
- Elsadek, N.E.; Lila, A.S.A.; Emam, S.E.; Shimizu, T.; Takata, H.; Ando, H.; Ishima, Y.; Ishida, T. Pegfilgrastim (PEG-G-CSF) induces anti-PEG IgM in a dose dependent manner and causes the accelerated blood clearance (ABC) phenomenon upon repeated administration in mice. Eur. J. Pharm. Biopharm. 2020, 152, 56–62. [Google Scholar] [CrossRef]
- Kozma, G.T.; Mészáros, T.; Vashegyi, I.; Fülöp, T.; Örfi, E.; Dézsi, L.; Rosivall, L.; Bavli, Y.; Urbanics, R.; Mollnes, T.E.; et al. Pseudo-anaphylaxis to Polyethylene Glycol (PEG)-Coated Liposomes: Roles of Anti-PEG IgM and Complement Activation in a Porcine Model of Human Infusion Reactions. ACS Nano 2019, 13, 9315–9324. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.; Abu Lila, A.S.; Shimizu, T.; Alaaeldin, E.; Hussein, A.; Sarhan, H.A.; Szebeni, J.; Ishida, T. PEGylated liposomes: Immunological responses. Sci. Technol. Adv. Mater. 2019, 20, 710–724. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ice, J.A.; Lessard, C.J.; Sivils, K.L. Interferons in Sjögren’s Syndrome: Genes, Mechanisms, and Effects. Front. Immunol. 2013, 4, 62005. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Wang, X.; Mao, X.; Yang, L.; Huang, K.; Zhang, F.; Wang, Y.; Luo, X.; Wang, C.; Peng, J.; et al. Absence of Interferon Regulatory Factor 1 Protects Against Atherosclerosis in Apolipoprotein E-Deficient Mice. Theranostics 2019, 9, 4688–4703. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.M.; Minter, M.R.; Newman, A.G.; Zhang, M.; Adlard, P.A.; Crack, P.J. Type-1 interferon signaling mediates neuro-inflammatory events in models of Alzheimer’s disease. Neurobiol. Aging 2014, 35, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Schustak, J.; Twarog, M.; Wu, X.; Wu, H.Y.; Huang, Q.; Bao, Y. Mechanism of Nucleic Acid Sensing in Retinal Pigment Epithelium (RPE): RIG-I Mediates Type I Interferon Response in Human RPE. J. Immunol. Res. 2021, 2021, 9975628. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.E.; Hughes, J.; Gu, Q.; Behdenna, A.; Singer, J.B.; Dennis, T.; Orton, R.J.; Varela, M.; Gifford, R.J.; Wilson, S.J.; et al. Fundamental properties of the mammalian innate immune system revealed by multispecies comparison of type I interferon responses. PLoS Biol. 2017, 15, e2004086. [Google Scholar] [CrossRef] [PubMed]
- Kerur, N.; Fukuda, S.; Banerjee, D.; Kim, Y.; Fu, D.; Apicella, I.; Varshney, A.; Yasuma, R.; Fowler, B.J.; Baghdasaryan, E.; et al. cGAS drives noncanonical-inflammasome activation in age-related macular degeneration. Nat. Med. 2018, 24, 50–61. [Google Scholar] [CrossRef]
- Orozco, L.D.; Chen, H.-H.; Cox, C.; Katschke, K.J., Jr.; Arceo, R.; Espiritu, C.; Caplazi, P.; Nghiem, S.S.; Chen, Y.-J.; Modrusan, Z.; et al. Integration of eQTL and a Single-Cell Atlas in the Human Eye Identifie s Causal Genes for Age-Related Macular Degeneration. Cell Rep. 2019, 30, 1246–1259.e1246. [Google Scholar] [CrossRef] [PubMed]
- Dhir, A.; Dhir, S.; Borowski, L.S.; Jimenez, L.; Teitell, M.; Rötig, A.; Crow, Y.J.; Rice, G.I.; Duffy, D.; Tamby, C.; et al. Mitochondrial double-stranded RNA triggers antiviral signalling in hum ans. Nature 2018, 560, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Chawla-Sarkar, M.; Lindner, D.J.; Liu, Y.F.; Williams, B.R.; Sen, G.C.; Silverman, R.H.; Borden, E.C. Apoptosis and interferons: Role of interferon-stimulated genes as mediators of apoptosis. Apoptosis 2003, 8, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, X.; Hu, C.; Du, Y.; Guo, C.; Di, W.; Zhao, W.; Huang, G.; Li, C.; Lu, Q.; et al. Type I IFN operates pyroptosis and necroptosis during multidrug-resistant A. baumannii infection. Cell Death Differ. 2018, 25, 1304–1318. [Google Scholar] [CrossRef] [PubMed]
- Stawowczyk, M.; Van Scoy, S.; Kumar, K.P.; Reich, N.C. The interferon stimulated gene 54 promotes apoptosis. J. Biol. Chem. 2011, 286, 7257–7266. [Google Scholar] [CrossRef] [PubMed]
- Thapa, R.J.; Nogusa, S.; Chen, P.; Maki, J.L.; Lerro, A.; Andrake, M.; Rall, G.F.; Degterev, A.; Balachandran, S. Interferon-induced RIP1/RIP3-mediated necrosis requires PKR and is lic ensed by FADD and caspases. Proc. Natl. Acad. Sci. USA 2013, 110, E3109–E3118. [Google Scholar] [CrossRef] [PubMed]
- Sarhan, J.; Liu, B.C.; Muendlein, H.I.; Weindel, C.G.; Smirnova, I.; Tang, A.Y.; Ilyukha, V.; Sorokin, M.; Buzdin, A.; Fitzgerald, K.A.; et al. Constitutive interferon signaling maintains critical threshold of MLKL expression to license necroptosis. Cell Death Differ. 2019, 26, 332–347. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Gerriets, T.; Wessels, C.; Walberer, M.; Kostin, S.; Stolz, E.; Zheleva, K.; Hocke, A.; Hippenstiel, S.; Preissner, K.T. Extracellular RNA mediates endothelial-cell permeability via vascular endothelial growth factor. Blood J. Am. Soc. Hematol. 2007, 110, 2457–2465. [Google Scholar] [CrossRef] [PubMed]
- Kannemeier, C.; Shibamiya, A.; Nakazawa, F.; Trusheim, H.; Ruppert, C.; Markart, P.; Song, Y.; Tzima, E.; Kennerknecht, E.; Niepmann, M. Extracellular RNA constitutes a natural procoagulant cofactor in blood coagulation. Proc. Natl. Acad. Sci. USA 2007, 104, 6388–6393. [Google Scholar] [CrossRef] [PubMed]
- Prünte, C.; Flammer, J. Choroidal capillary and venous congestion in central serous chorioretinopathy. Am. J. Ophthalmol. 1996, 121, 26–34. [Google Scholar] [CrossRef]
- Iijima, H.; Iida, T.; Murayama, K.; Imai, M.; Gohdo, T. Plasminogen activator inhibitor 1 in central serous chorioretinopathy. Am. J. Ophthalmol. 1999, 127, 477–478. [Google Scholar] [CrossRef] [PubMed]
- Mutter, J.; Yeter, D. Kawasaki’s disease, acrodynia, and mercury. Curr. Med. Chem. 2008, 15, 3000–3010. [Google Scholar] [CrossRef]
- Stejskal, J.; Stejskal, V.D. The role of metals in autoimmunity and the link to neuroendocrinology. Neuro Endocrinol. Lett. 1999, 20, 351–364. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Underlying Mechanism | Ocular Inflammation | Licensed Vaccine * | Reference | |
|---|---|---|---|---|
| Direct infection | ARN, MEWDS, keratitis, iris heterochromia, uveitis | MMR vaccine (Priorix, strain Wistar RA 27/3; GlaxoSmithKline, Brentford, Middlesex), nasal spray influenza vaccine ^, rotavirus vaccines (RotaTeq® by Merck & Co., Inc., Kenilworth, NJ, USA), yellow fever vaccine ^, live attenuated varicella vaccine (Zostavax® by Merck & Co., Inc., Kenilworth, NJ, USA) | [15,16,17,18,20,21,22,31] | |
| ASIA | Aluminum-related | NMO | HPV vaccine (Gardasil, Merck & Co., Inc., USA) | [32,33] |
| Interactions between adjuvants and adaptive immune components | Uveitis, VKH | HBV vaccine (Recombivax HB and Engerix-B) | [24,30] | |
| AS01B induced | Uveitis, ARN, HZO reactivation, stromal keratitis | VZV vaccine (Shingrix by GlaxoSmithKline, Brentford, UK) | [34,35,36,37,38] | |
| Hypersensitivity | Optic neuritis, NMOSD, MEWDS, ERD | Rabies vaccine (Rabipur® by Hoechst Marion Roussel Ltd, Ankleshwar, Gujarat), influenza vaccines (split, inactivated virions of swine influenza A New Jersey and influenza B Victoria lineages), MMR vaccine ^, HPV vaccine (Gardasil, Merck & Co., Inc., Kenilworth, NJ, USA) | [39,40,41,42,43,44] | |
| Molecular mimicry | Optic neuritis, uveitis, APMPPE, TINU syndrome | HPV vaccine (Gardasil, Merck & Co., Inc., Kenilworth, NJ, USA), purified hepatitis B vaccine ^ | [41,42,45,46,47] | |
| PEG-induced allergic reactions | Optic neuropathy, VKH, MEWDS, AMN | COVID-19 vaccines (BNT163b2 by Pfizer-BioNTech, BioNTech Manufacturing GmbH, Berlin, Germany, and mRNA-1273, by Moderna Biotech, Madrid, Spain) | [48] | |
| Activation of Type 1 IFN and inflammatory response of RPE cells | MEWDS | [49] | ||
| Free extracellular mRNA | CSR | [50] | ||
| Specific components (thimerosal) | Optic neuritis | Influenza (Vaxigrip by Sanofi Pasteur, Lyon, France) | [51] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, Y.; Kamoi, K.; Zong, Y.; Zhang, J.; Yang, M.; Ohno-Matsui, K. Vaccines and the Eye: Current Understanding of the Molecular and Immunological Effects of Vaccination on the Eye. Int. J. Mol. Sci. 2024, 25, 4755. https://doi.org/10.3390/ijms25094755
Zou Y, Kamoi K, Zong Y, Zhang J, Yang M, Ohno-Matsui K. Vaccines and the Eye: Current Understanding of the Molecular and Immunological Effects of Vaccination on the Eye. International Journal of Molecular Sciences. 2024; 25(9):4755. https://doi.org/10.3390/ijms25094755
Chicago/Turabian StyleZou, Yaru, Koju Kamoi, Yuan Zong, Jing Zhang, Mingming Yang, and Kyoko Ohno-Matsui. 2024. "Vaccines and the Eye: Current Understanding of the Molecular and Immunological Effects of Vaccination on the Eye" International Journal of Molecular Sciences 25, no. 9: 4755. https://doi.org/10.3390/ijms25094755
APA StyleZou, Y., Kamoi, K., Zong, Y., Zhang, J., Yang, M., & Ohno-Matsui, K. (2024). Vaccines and the Eye: Current Understanding of the Molecular and Immunological Effects of Vaccination on the Eye. International Journal of Molecular Sciences, 25(9), 4755. https://doi.org/10.3390/ijms25094755