

Berberis vulgaris L. Root Extract as a Multi-Target Chemopreventive Agent against Colon Cancer Causing Apoptosis in Human Colon Adenocarcinoma Cell Lines
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Biological Activity of BVR
2.1.1. Antiradical and Antilipooxigenase Activity of BVR
2.1.2. Influence of BVR and Its Main Constituents on the Viability of CCD841 CoN, LS180, and HT-29 Cells Using the MTT Assay
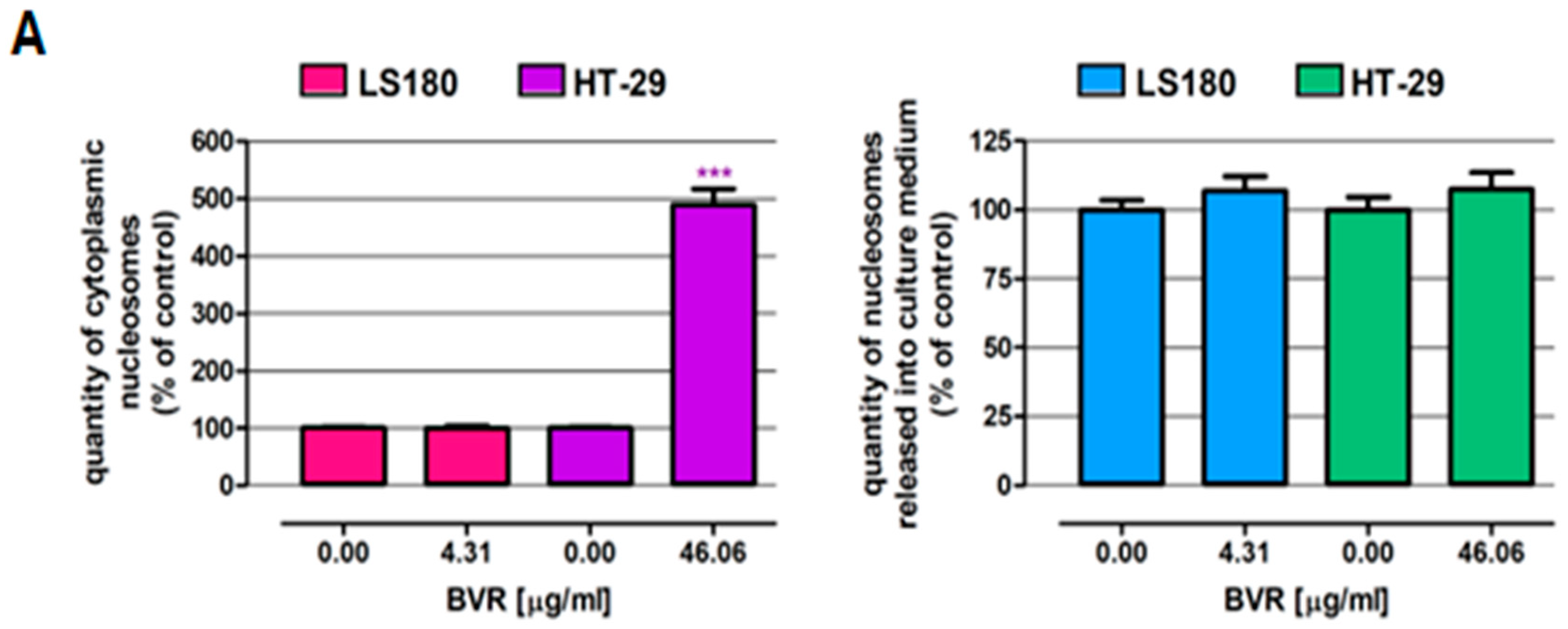
2.1.3. PCR Analysis
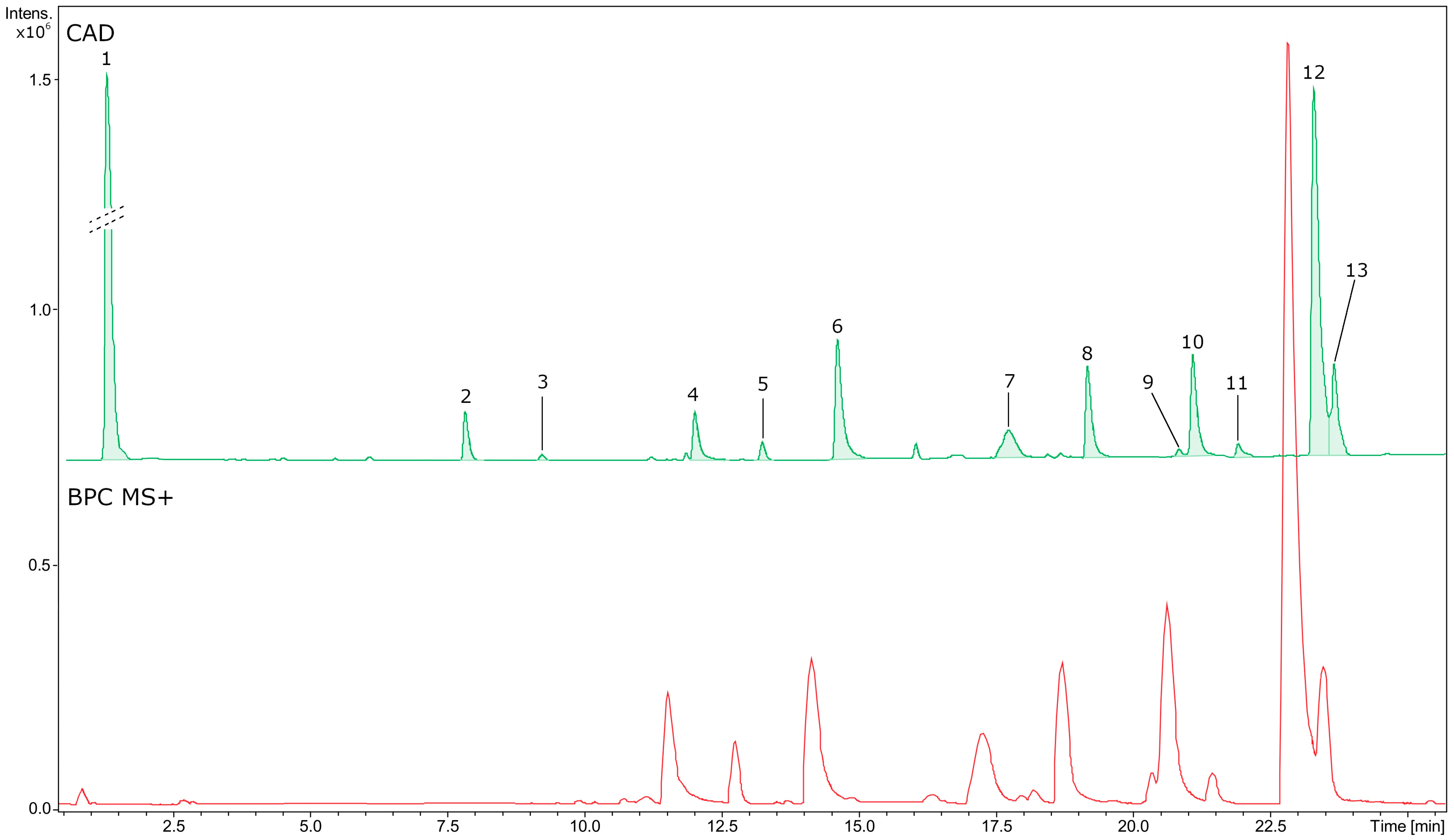
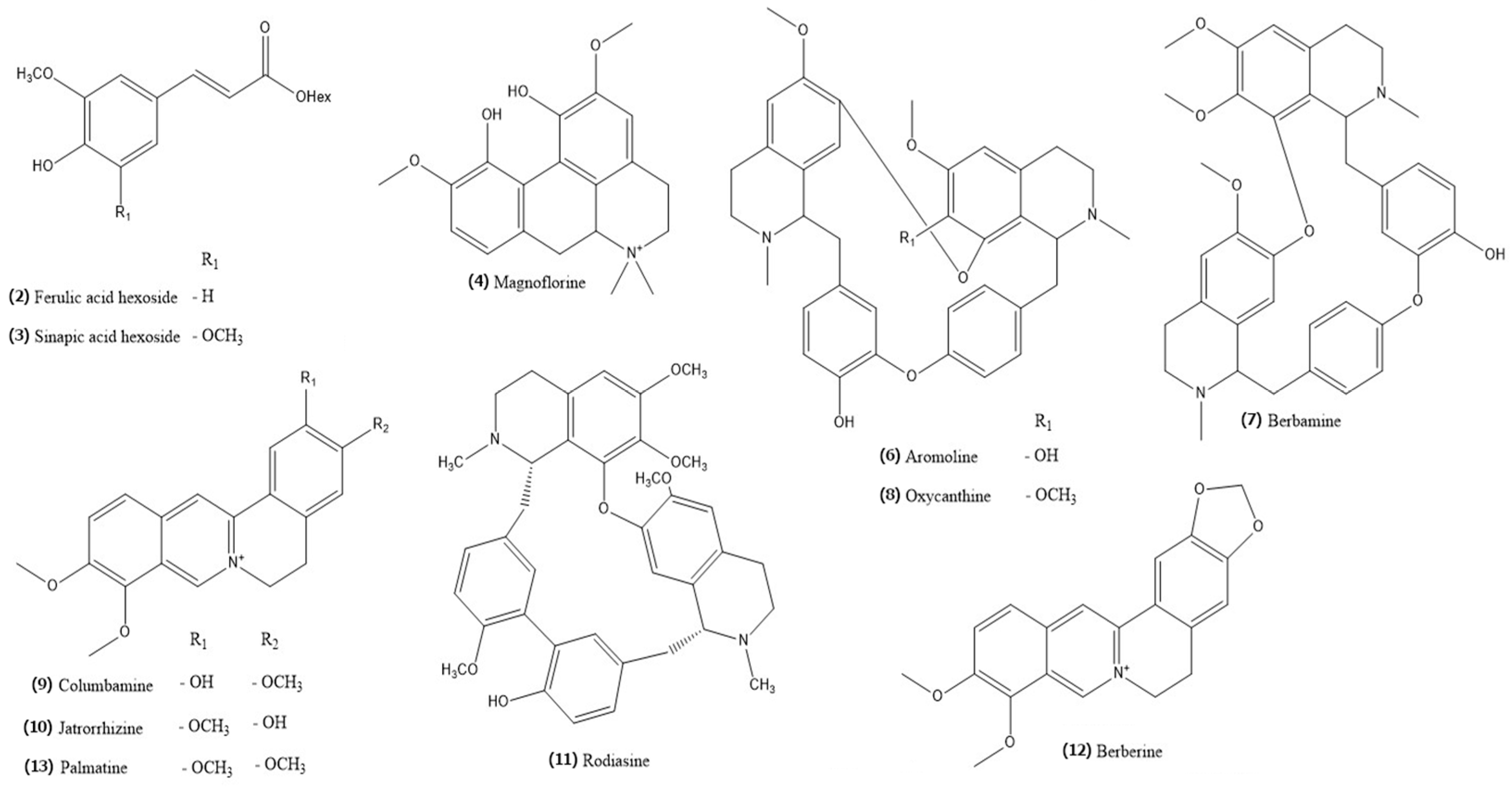
2.2. Phytochemical Profiling and Quantification of Major Specialized Metabolites
3. Discussion
4. Materials and Methods
4.1. Plant Material and Preparation of Extract
4.2. Chemicals and Reagents
4.3. Antiradical Activity Analyses
4.3.1. Determination of Antiradical Potential with the DPPH• Assay
4.3.2. Determination of Antiradical Capacity with the ABTS•+ Assay
4.3.3. Oxygen Radical Absorbance Capacity (ORAC) Assay
4.4. Lipoxygenase (LOX) Inhibitor Screening Assay
4.5. Cell Cultures
4.6. Cell Viability Assessment—MTT Assay
4.7. Cell Death Assessment—ELISA
4.8. Cell Death Detection—Nuclear Double Staining
4.9. Gene Expression Analysis
4.10. Phytochemical Profiling and Quantification of Major Specialized Metabolites Using LC-UV-MS/MS Technique
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Cancer Research Fund International. Available online: https://www.wcrf-uk.org/ (accessed on 12 March 2023).
- Labianca, R.; Nordlinger, B.; Beretta, G.D.; Mosconi, S.; Mandalà, M.; Cervantes, A.; Arnold, D. Early colon cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 24, 64–72. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer. (accessed on 12 March 2023).
- Simon, K. Colorectal cancer development and advances in screening. Clin. Interv. Aging 2016, 11, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Bardelčíková, A.; Šoltys, J.; Mojžiš, J. Oxidative Stress, Inflammation and Colorectal Cancer: An Overview. Antioxidants 2023, 12, 901. [Google Scholar] [CrossRef] [PubMed]
- George, B.P.; Chandran, R.; Abrahamse, H. Role of phytochemicals in cancer chemoprevention: Insights. Antioxidants 2021, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Maresso, K.C.; Tsai, K.Y.; Brown, P.H.; Szabo, E.; Lippman, S.; Hawk, E.T. Molecular cancer prevention: Current status and future directions. CA A Cancer J. Clin. 2015, 65, 345–383. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, Y.; Yong, P.E.; Qian, Z.; Xiao, P. New collection of crude drugs in Chinese pharmacopoeia 2010 II. Sankezhen (Berberis spp.). Chin. Herb. Med. 2011, 3, 272–288. [Google Scholar]
- Henriette’s Herbal Homepage. Available online: https://www.henriettes-herb.com (accessed on 26 July 2020).
- Imanshahidi, M.; Hosseinzadeh, H. Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine. Phytother. Res. 2008, 22, 999–1012. [Google Scholar] [CrossRef]
- Li, Y.; Wang, P.; Chai, M.J.; Yang, F.; Li, H.S.; Zhao, J.; Wang, H.; Lu, D.D. Effects of berberine on serum inflammatory factors and carotid atherosclerotic plaques in patients with acute cerebral ischemic stroke. China J. Chin. Mater. Medica 2016, 41, 4066–4071. [Google Scholar] [CrossRef]
- Meng, S.; Wang, L.S.; Huang, Z.Q.; Zhou, Q.; Sun, Y.G.; Cao, J.T.; Li, Y.G.; Wang, C.Q. Berberine ameliorates inflammation in patients with acute coronary syndrome following percutaneous coronary intervention. Clin. Exp. Pharmacol. Physiol. 2012, 39, 406–411. [Google Scholar] [CrossRef]
- Kiasalari, Z.; Khalili, M.; Ahmadi, P. Effect of alcoholic extract of Berberis vulgaris fruit on acute and chronic inflammation in male rats. J. Babol Univ. Med. Sci. 2011, 13, 28–35. [Google Scholar]
- Minaiyan, M.; Ghannadi, A.; Mahzouni, P.; Jaffari-Shirazi, E. Comparative study of Berberis vulgaris fruit extract and berberine chloride effects on acetic acid-induced colitis in rats. Iran. J. Pharm. Res. 2011, 10, 97–104. [Google Scholar] [PubMed]
- Majeed, W.; Aslam, B.; Javed, I.; Khaliq, T.; Muhammad, F.; Ali, A.; Raza, A. Histopathological evaluation of gastro protective effect of Berberis vulgaris (Zereshk) seeds against aspirin induced ulcer in albino mice. Pak. J. Pharm. Sci. 2015, 28, 1953–1958. [Google Scholar] [PubMed]
- Mohebali, S.; Nasri, S.; Kamalinejhad, M.; Noori, A.S. Antinociceptive & anti-inflammatory effects of Berberis vulgaris L. root’s hydroalcoholic extract and determination of it’s possible antinociceptive mechanism in male mice. J. Paramed. Sci. (JPS) 2011, 2, 12–18. [Google Scholar]
- Ivanovska, N.; Philipov, S. Study on the anti-inflammatory action of Berberis vulgaris root extract, alkaloid fractions and pure alkaloids. Int. J. Immunopharmacol. 1996, 18, 553–561. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ (accessed on 26 April 2023).
- Abd El-Wahab, A.E.; Ghareeb, D.A.; Sarhan, E.E.M.; Abu-Serie, M.M.; el Demellawy, M.A. In vitro biological assessment of Berberis vulgaris and its active constituent, berberine: Antioxidants, anti-acetylcholinesterase, anti-diabetic and anticancer effects. BMC Complement. Altern. Med. 2013, 13, 218. [Google Scholar] [CrossRef]
- Gird, C.E.; Ligiaelena, D.U.; Costea, T.; Nencu, I.; Popescu, M.; Balaci, T.; Olaru, O. Research regarding obtaining herbal extracts with antitumor activity. note ii. phytochemical analysis, antioxidant activity and cytotoxic effects of Chelidonium majus L., Medicago sativa L. and Berberis vulgaris L. dry extracts. Medicago sativa L. and Berberis vulgaris L. dry extracts. Farmacia 2017, 65, 703–708. [Google Scholar]
- Ghafourian, E.; Sadeghifard, N.; Pakzad, I.; Valizadeh, N.; Maleki, A.; Jafari, F.; Ghiasvand, N.; Abdi, J.; Shokoohinia, Y.; Ghafourian, S. Ethanolic extract of Berberis vulgaris fruits inhibits the proliferation of MCF-7 breast cancer cell line through induction of apoptosis. Former. Curr. Drug Targets-Infect. Disord. 2017, 17, 192–198. [Google Scholar] [CrossRef] [PubMed]
- El Khalki, L.; Tilaoui, M.; Jaafari, A.; Ait Mouse, H.; Zyad, A. Studies on the Dual Cytotoxicity and Antioxidant Properties of Berberis vulgaris Extracts and Its Main Constituent Berberine. Adv. Pharmacol. Sci. 2018, 2018, 3018498. [Google Scholar] [CrossRef]
- Lin, C.C.; Ng, L.T.; Hsu, F.F.; Shieh, D.E.; Chiang, L.C. Cytotoxic effects of Coptis chinensis and Epimedium sagittatum extracts and their major constituents (berberine, coptisine and icariin) on hepatoma and leukaemia cell growth. Clin. Exp. Pharmacol. Physiol. 2004, 31, 65–69. [Google Scholar] [CrossRef]
- Och, A.; Olech, M.; Bąk, K.; Kanak, S.; Cwener, A.; Cieśla, M.; Nowak, R. Evaluation of the antioxidant and anti-lipoxygenase activity of Berberis vulgaris L. leaves, fruits, and stem and their LC MS/MS polyphenolic profile. Antioxidants 2023, 12, 1467. [Google Scholar] [CrossRef]
- Gorizpa, M.; Bahmanyar, F.; Mirmoghtadaie, L.; Shafaei, F. Evaluation of Antioxidant and Antimicrobial Properties of Root and Stem Bark Extracts of Three Species of Barberry in Bread. Res. Innov. Food Sci. Technol. 2022, 10, 413–426. [Google Scholar] [CrossRef]
- Luo, A.; Fan, Y. Antioxidant activities of berberine hydrochloride. J. Med. Plants Res. 2011, 5, 3702–3707. [Google Scholar]
- Chaves, S.K.; Afzal, M.I.; Islam, M.T.; Hameed, A.; Da Mata, A.M.; da Silva Araújo, L.; Ali, S.W.; Rolim, H.M.; De Medeiros, M.D.; Costa, E.V.; et al. Palmatine antioxidant and anti-acetylcholinesterase activities: A pre-clinical assessment. Cell. Mol. Biol. 2020, 66, 54–59. [Google Scholar] [CrossRef]
- Villinski, J.R.; Dumas, E.R.; Chai, H.B.; Pezzuto, J.M.; Angerhofer, C.K.; Gafner, S. Antibacterial activity and alkaloid content of Berberis thunbergii, Berberis vulgaris and Hydrastis canadensis. Pharm. Biol. 2003, 41, 551–557. [Google Scholar] [CrossRef]
- Xu, T.; Kuang, T.; Du, H.; Li, Q.; Feng, T.; Zhang, Y.; Fan, G. Magnoflorine: A review of its pharmacology, pharmacokinetics and toxicity. Pharmacol. Res. 2020, 152, 104632. [Google Scholar] [CrossRef]
- Kalmarzi, R.N.; Naleini, S.N.; Ashtary-Larky, D.; Peluso, I.; Jouybari, L.; Rafi, A.; Ghorat, F.; Heidari, N.; Sharifian, F.; Mardaneh, J.; et al. Anti-inflammatory and immunomodulatory effects of barberry (Berberis vulgaris) and its main compounds. Oxidative Med. Cell. Longev. 2019, 2019, 6183965. [Google Scholar] [CrossRef] [PubMed]
- Sarraf, M.; Beig Babaei, A.; Naji-Tabasi, S. Investigating functional properties of barberry species: An overview. J. Sci. Food Agric. 2019, 99, 5255–5269. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Jiang, K.; Wu, H.; Yang, C.; Zhao, G.; Deng, G. Magnoflorine ameliorates lipopolysaccharide-induced acute lung injury via suppressing NF-κB and MAPK activation. Front. Pharmacol. 2018, 9, 393441. [Google Scholar] [CrossRef]
- Mokhber-Dezfuli, N.; Saeidnia, S.; Gohari, A.; Kurepaz-Mahmoodabadi, M. Phytochemistry and pharmacology of berberis species. Pharmacogn. Rev. 2014, 8, 8–15. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Kaushik, N. Phytochemical and pharmacological studies in genus Berberis. Phytochem. Rev. 2012, 11, 523–542. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, H.; Chen, W.; Huang, P. Berberine induces apoptosis and arrests the cell cycle in multiple cancer cell lines. Arch. Med. Sci. AMS 2023, 19, 1530–1537. [Google Scholar] [CrossRef]
- Och, A.; Zalewski, D.; Komsta, Ł.; Kołodziej, P.; Kocki, J.; Bogucka-Kocka, A. Cytotoxic and proapoptotic activity of sanguinarine, berberine, and extracts of chelidonium majus L. and berberis thunbergii DC. Toward hematopoietic cancer cell lines. Toxins 2019, 11, 485. [Google Scholar] [CrossRef] [PubMed]
- Bala, M.; Pratap, K.; Verma, P.K.; Singh, B.; Padwad, Y. Validation of ethnomedicinal potential of Tinospora cordifolia for anticancer and immunomodulatory activities and quantification of bioactive molecules by HPTLC. J. Ethnopharmacol. 2015, 175, 131–137. [Google Scholar] [CrossRef]
- Mou, L.; Liang, B.; Liu, G.; Jiang, J.; Liu, J.; Zhou, B.; Huang, J.; Zang, N.; Liao, Y.; Ye, L.; et al. Berbamine exerts anticancer effects on human colon cancer cells via induction of autophagy and apoptosis.; inhibition of cell migration and MEK/ERK signalling pathway. J. BUON 2019, 24, 1870–1875. [Google Scholar] [PubMed]
- Zhang, L.L.; Li-Na, M.A.; Dan, Y.A.; Zhang, C.E.; Dan, G.A.; Xiong, Y.; Sheng, F.Y.; Xiao-Ping, D.O.; Xiao-He, X.I. Dynamic monitoring of the cytotoxic effects of protoberberine alkaloids from Rhizoma Coptidis on HepG2 cells using the xCELLigence system. Chin. J. Nat. Med. 2014, 12, 428–435. [Google Scholar] [CrossRef]
- Kalaiarasi, A.; Anusha, C.; Sankar, R.; Rajasekaran, S.; John Marshal, J.; Muthusamy, K.; Ravikumar, V. Plant isoquinoline alkaloid berberine exhibits chromatin remodeling by modulation of histone deacetylase to induce growth arrest and apoptosis in the A549 cell line. J. Agric. Food Chem. 2016, 64, 9542–9550. [Google Scholar] [CrossRef]
- Su, Y.H.; Tang, W.C.; Cheng, Y.W.; Sia, P.; Huang, C.C.; Lee, Y.C.; Jiang, H.Y.; Wu, M.H.; Lai, I.L.; Lee, J.W.; et al. Targeting of multiple oncogenic signaling pathways by Hsp90 inhibitor alone or in combination with berberine for treatment of colorectal cancer. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2015, 1853, 2261–2272. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Yang, Q.; Mu, Y.; Zhou, L.; Liu, Y.; Zhou, Q.; He, B. Berberine inhibits the proliferation of colon cancer cells by inactivating Wnt/β-catenin signaling. Int. J. Oncol. 2012, 41, 292–298. [Google Scholar] [CrossRef]
- Chidambara Murthy, K.N.; Jayaprakasha, G.K.; Patil, B.S. The natural alkaloid berberine targets multiple pathways to induce cell death in cultured human colon cancer cells. Eur. J. Pharmacol. 2012, 688, 14–21. [Google Scholar] [CrossRef]
- Dai, W.; Mu, L.; Cui, Y.; Li, Y.; Chen, P.; Xie, H.; Wang, X. Berberine promotes apoptosis of colorectal cancer via regulation of the long non-coding RNA (lncRNA) cancer susceptibility candidate 2 (CASC2)/AU-binding factor 1 (AUF1)/B-cell CLL/lymphoma 2 (Bcl-2) axis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 730–738. [Google Scholar] [CrossRef]
- Vuddanda, P.R.; Chakraborty, S.; Singh, S. Berberine: A potential phytochemical with multispectrum therapeutic activities. Expert Opin. Investig. Drugs 2010, 19, 1297–1307. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, X.; Wu, P.; Wink, M.; Li, J.; Dian, L.; Liang, Z. Jatrorrhizine inhibits mammary carcinoma cells by targeting TNIK mediated Wnt/β-catenin signalling and epithelial-mesenchymal transition (EMT). Phytomedicine 2019, 63, 153015. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Gao, X.Y.; Yang, S.Q.; Sun, Z.X.; Dian, L.L.; Qasim, M.; Phyo, A.T.; Liang, Z.S.; Sun, Y.F. Jatrorrhizine inhibits colorectal carcinoma proliferation and metastasis through Wnt/β-catenin signaling pathway and epithelial–mesenchymal transition. Drug Des. Dev. Ther. 2019, 13, 2235–2247. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Zhang, H.; Zhang, W.W.; Huang, J.T.; Song, E.L.; Chen, S.G.; He, F.; Xu, J.; Wang, H.Q. Neuroprotective effect of Jatrorrhizine on hydrogen peroxide-induced cell injury and its potential mechanisms in PC12 cells. Neurosci. Lett. 2011, 498, 227–231. [Google Scholar] [CrossRef]
- Kumari, S.; Kaladhar, D.S.; Solmon, K.S.; Malla, R.R.; Kishore, G. Anti-proliferative and metastatic protease inhibitory activities of protoberberines: An in silico and in vitro approaches. Process Biochem. 2013, 48, 1565–1571. [Google Scholar] [CrossRef]
- Rolle, J.; Asante, D.O.; Kok-Fong, L.L.; Boucetta, H.; Seidu, T.A.; Tai, L.L.K.; Alolga, R.N. Jatrorrhizine: A review of its pharmacological effects. J. Pharm. Pharmacol. 2021, 73, 709–719. [Google Scholar] [CrossRef]
- Ali, D.; Ali, H. Assessment of DNA damage and cytotoxicity of palmatine on human skin epithelial carcinoma cells. Toxicol. Environ. Chem. 2014, 96, 941–950. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Zhang, X.F.; Tang, Y.L.; Xiang, J.F.; Tian, M.Y. Studies of the interactions between three protoberberine alkaloids and Bcl-2 by fluorescence spectroscopy. Acta Chim. Sin. 2011, 69, 247. [Google Scholar]
- Wu, J.; Xiao, O.; Zhang, N.; Xue, C.; Leung, A.W.; Zhang, H.; Xu, C.; Tang, Q. Photodynamic action of palmatine hydrochlo-ride on colon adenocarcinoma HT-29 cells. Photodiagnosis Photodyn. Ther. 2016, 15, 53–58. [Google Scholar] [CrossRef]
- Inbaraj, J.J.; Kukielczak, B.M.; Bilski, P.; He, Y.Y.; Sik, R.H.; Chignell, C.F. Photochemistry and photocytotoxicity of alkaloids from Goldenseal (Hydrastis canadensis L.). 2. Palmatine, hydrastine, canadine, and hydrastinine. Chem. Res. Toxicol. 2006, 19, 739–744. [Google Scholar] [CrossRef]
- Hirakawa, K.; Kawanishi, S.; Hirano, T. The mechanism of guanine specific photooxidation in the presence of berberine and palmatine: Activation of photosensitized singlet oxygen generation through DNA-binding interaction. Chem. Res. Toxicol. 2005, 18, 1545–1552. [Google Scholar] [CrossRef]
- Ma, W.K.; Li, H.; Dong, C.L.; He, X.; Guo, C.R.; Zhang, C.F.; Yu, C.H.; Wang, C.Z.; Yuan, C.S. Palmatine from Mahonia bealei attenuates gut tumorigenesis in ApcMin/+ mice via inhibition of inflammatory cytokines. Mol. Med. Rep. 2016, 14, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Yao, Y.; Shen, B.; Liu, J.; Pan, Q.; Liu, N.; Li, L.; Huang, J.; Long, Z.; Shao, L. Columbamine suppresses the proliferation and malignization of colon cancer cells via abolishing Wnt/β-catenin signaling pathway. Cancer Manag. Res. 2019, 23, 8635–8645. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Zhang, X.W.; Zhai, H.J.; Zhang, D.; Ma, S.Y. Magnoflorine inhibits human gastric cancer progression by inducing autophagy, apoptosis and cell cycle arrest by JNK activation regulated by ROS. Biomed. Pharmacother. 2020, 125, 109118. [Google Scholar] [CrossRef]
- Mohamed, S.M.; Hassan, E.M.; Ibrahim, N.A. Cytotoxic and antiviral activities of aporphine alkaloids of Magnolia grandiflora L. Nat. Prod. Res. 2010, 24, 1395–1402. [Google Scholar] [CrossRef]
- Bai, L.; Zhang, H.; Liu, Q.; Zhao, Y.; Cui, X.; Guo, S.; Zhang, L.; Ho, C.T.; Bai, N. Chemical characterization of the main bioactive constituents from fruits of Ziziphus jujuba. Food Funct. 2016, 7, 2870–2877. [Google Scholar] [CrossRef]
- Koutova, D.; Kulhava, M.; Havelek, R.; Majorosova, M.; Královec, K.; Habartova, K.; Hošťálková, A.; Opletal, L.; Cahlikova, L.; Řezáčová, M. Bersavine: A Novel Bisbenzylisoquinoline Alkaloid with Cytotoxic, Antiproliferative and Apoptosis-Inducing Effects on Human Leukemic Cells. Molecules 2020, 25, 964. [Google Scholar] [CrossRef]
- Habartová, K.; Havelek, R.; Seifrtova, M.; Hostalkova, A.; Cahlikova, L.; Rezacova, M. A new isoquinoline alkaloid bersavine as a possible anticancer agent. Ann. Oncol. 2017, 28, v21. [Google Scholar] [CrossRef]
- Hošt’álková, A.; Novák, Z.; Pour, M.; Jirošová, A.; Opletal, L.; Kuneš, J.; Cahlíková, L. Berbanine: A new isoquinoline-isoquinolone alkaloid from Berberis vulgaris (Berberidaceae). Nat. Prod. Commun. 2013, 8, 1934578X1300800407. [Google Scholar] [CrossRef]
- Liang, Y.; Qiu, X.; Xu, R.Z.; Zhao, X.Y. Berbamine inhibits proliferation and induces apoptosis of KU812 cells by increasing Smad3 activity. J. Zhejiang Univ. Sci. B 2011, 12, 568–574. [Google Scholar] [CrossRef]
- Liang, Y.; Xu, R.Z.; Zhang, L.; Zhao, X.Y. Berbamine, a novel nuclear factor κB inhibitor, inhibits growth and induces apoptosis in human myeloma cells. Acta Pharmacol. Sin. 2009, 30, 1659–1665. [Google Scholar] [CrossRef]
- Hussain, S.F. Secobisbenzylisoquinoline alkaloids-chemistry and pharmacology: A review. Baqai J. Health Sci. 2018, 21, 2. [Google Scholar]
- Saedi, T.A.; Ghafourian, S.; Jafarlou, M.; Sabariah, M.N.; Ismail, P.; Eusni, R.M.; Othman, F. Berberis vulgaris fruit crude extract as a novel anti-leukaemic agent. J. Biol. Regul. Homeost. Agents 2015, 29, 395–399. [Google Scholar] [PubMed]
- Liu, L.; Sun, L.; Zheng, J.; Cui, L. Berberine modulates Keratin 17 to inhibit cervical cancer cell viability and metastasis. J. Recept. Signal Transduct. 2021, 41, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xie, N.; Chai, Y.; Nie, Y.; Liu, K.; Liu, Y.; Yang, Y.; Su, J.; Zhang, C. Apoptosis induction, a sharp edge of berberine to exert anti-cancer effects, focus on breast, lung, and liver cancer. Front. Pharmacol. 2022, 13, 803717. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Feng, Y.; Zhu, M.; Tsang, C.M.; Man, K.; Tong, Y.; Tsao, S.W. Berberine induces autophagic cell death and mitochondrial apoptosis in liver cancer cells: The cellular mechanism. J. Cell. Biochem. 2010, 111, 1426–1436. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, Q.; Lin, Z.; Yang, P.; Dou, K.; Zhang, R. Berberine inhibits growth of liver cancer cells by suppressing glutamine uptake. OncoTargets Ther. 2019, 31, 11751–11763. [Google Scholar] [CrossRef]
- Liu, Y.; Nwafor, E.O.; Li, Z.; Wang, J.; Feng, X.; Li, H.; Jia, B.; Ma, H.; He, J.; Pi, J.; et al. Effects of Berberine on Liver Cancer. Nat. Prod. Commun. 2022, 17, 1934578X221102032. [Google Scholar] [CrossRef]
- Hanachi, P.; Othman, F.; Motaleb, G.R. Effect of Berberis vulgaris aqueous extract on the apoptosis, sodium and potassium in hepatocarcinogenic rats. Iran. J. Basic Med. Sci. 2008, 11, 62–69. [Google Scholar]
- Wu, J.; Xiao, Q.; Zhang, N.; Xue, C.; Leung, A.W.; Zhang, H.; Tang, Q.J.; Xu, C. Palmatine hydrochloride mediated photodynamic inactivation of breast cancer MCF-7 cells: Effectiveness and mechanism of action. Photodiagnosis Photodyn. Ther. 2016, 15, 133–138. [Google Scholar] [CrossRef]
- Kaboli, P.J.; Rahmat, A.; Ismail, P.; Ling, K.H. Targets and mechanisms of berberine, a natural drug with potential to treat cancer with special focus on breast cancer. Eur. J. Pharmacol. 2014, 740, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Tomosaka, H.; Chin, Y.W.; Salim, A.A.; Keller, W.J.; Chai, H.; Kinghorn, A.D. Antioxidant and cytoprotective compounds from Berberis vulgaris (barberry). Phytother. Res. 2008, 22, 979–981. [Google Scholar] [CrossRef] [PubMed]
- Hoshyar, R.; Mahboob, Z.; Zarban, A. The antioxidant and chemical properties of Berberis vulgaris and its cytotoxic effect on human breast carcinoma cells. Cytotechnology 2016, 68, 1207–1213. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Shi, Y.; Cao, H.; Chaturvedi, R.; Calcutt, M.W. Berberine induces caspase-independent cell death in colon tumor cells through activation of apoptosis-inducing factor. PLoS ONE 2012, 7, e36418. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.S.; Liu, K.; Wang, J.; Shi, Y.M. Anticancer Potential of Nature-Derived Isoquinoline Alkaloids (A Review). Russ. J. Gen. Chem. 2023, 93, 1294–1310. [Google Scholar] [CrossRef]
- Liu, R.; Cao, Z.; Pan, Y.; Zhang, G.; Yang, P.; Guo, P.; Zhou, Q. Jatrorrhizine hydrochloride inhibits the proliferation and neovascularization of C8161 metastatic melanoma cells. Anti-Cancer Drugs 2013, 24, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Sindhoor, S.M.; Naveen, N.R.; Rao, G.K.; Gopan, G.; Chopra, H.; Park, M.N.; Alshahrani, M.M.; Jose, J.; Emran, T.B.; Kim, B. A spotlight on alkaloid nanoformulations for the treatment of lung cancer. Front. Oncol. 2022, 12, 994155. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Abbas, K.; Raza, M.T.; Yahya, H.; Saifi, M.F.; Kamal, S. Evaluation of Berberis vulgaris Phytochemicals for Targeting PIM1 Kinase in Prostate Cancer: An In silico Approach. Int. Res. J. Oncol. 2024, 7, 60–72. [Google Scholar]
- Hostalkova, A.; Marikova, J.; Opletal, L.; Korabecny, J.; Hulcova, D.; Kunes, J.; Novakova, L.; Perez, D.I.; Jun, D.; Kucera, T. Isoquinoline Alkaloids from Berberis vulgaris as Potential Lead Compounds for the Treatment of Alzheimer’s Disease. J. Nat. Prod. 2019, 82, 239–248. [Google Scholar] [CrossRef]
- Hošťálková, A.; Novák, Z.; Hrabinová, M.; Štěpánková, Š.; Peréz, D.; Kuneš, J.; Cahlíková, L.; Opletal, L. Alkaloids from Berberis vulgaris and their biological activity connected to Alzheimer’s disease. Planta Medica 2015, 81, PW_126. [Google Scholar] [CrossRef]
- Novák, Z.; Hošt’álková, A.; Opletal, L.; Nováková, L.; Hrabinová, M.; Kuneš, J.; Cahliková, L. (+)-chenabinol (revised NMR data) and two new alkaloids from Berberis vulgaris and their biological activity. Nat. Prod. Commun. 2015, 10, 1934578X1501001015. [Google Scholar] [CrossRef]
- Zeng, X.H.; Zeng, X.J.; Li, Y.Y. Efficacy and safety of berberine for congestive heart failure secondary to ischemic or idiopathic dilated cardiomyopathy. Am. J. Cardiol. 2003, 92, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Barrios, V.; Escobar, C.; Cicero, A.F.G.; Burke, D.; Fasching, P.; Banach, M.; Bruckert, E. A nutraceutical approach (Armolipid Plus) to reduce total and LDL cholesterol in individuals with mild to moderate dyslipidemia: Review of the clinical evidence. Atheroscler. Suppl. 2017, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Zhao, Z.X.; Huang, M.; Feng, R.; He, C.Y.; Ma, C.; Luo, S.-H.; Fu, J.; Wen, B.Y.; Ren, L.; et al. Effect of Berberine on promoting the excretion of cholesterol in high-fat diet-induced hyperlipidemic hamsters. J. Transl. Med. 2015, 13, 278. [Google Scholar] [CrossRef]
- Brusq, J.M.; Ancellin, N.; Grondin, P.; Guillard, R.; Martin, S.; Saintillan, Y.; Issandou, M. Inhibition of lipid synthesis through activation of AMP kinase: An additional mechanism for the hypolipidemic effects of berberine. J. Lipid Res. 2006, 47, 1281–1288. [Google Scholar] [CrossRef]
- Wang, L.; Peng, L.; Wei, G.; Ge, H. Therapeutic Effects of Berberine Capsule on Patients with Mild Hyperlipidemia. Zhongguo Zhong Xi Yi Jie He Za Zhi Zhongguo Zhongxiyi Jiehe Zazhi Chin. J. Integr. Tradit. West. Med. 2016, 36, 681–684. [Google Scholar]
- Luo, T.; Shen, X.Y.; Li, S. The protective effect of jatrorrhizine against oxidative stress in primary rat cortical neurons. CNS Neurol. Disord. Drug Targets 2017, 16, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Och, A.; Podgórski, R.; Nowak, R. Biological activity of berberine—A summary update. Toxins 2020, 12, 713. [Google Scholar] [CrossRef] [PubMed]
- Nor, N.M.; Noor, S.M.; Tohit, E.M.; Othman, F. Anti-inflammatory effect of Berberis vulgaris extract in vivo. Atherosclerosis 2020, 315, e117. [Google Scholar] [CrossRef]
- Och, A.; Och, M.; Nowak, R.; Podgórska, D.; Podgórski, R. Berberine, a herbal metabolite in the metabolic syndrome: The risk factors, course, and consequences of the disease. Molecules 2022, 27, 1351. [Google Scholar] [CrossRef]
- Tabeshpour, J.; Imenshahidi, M.; Hosseinzadeh, H. A review of the effects of Berberis vulgaris and its major component, berberine, in metabolic syndrome. Iran. J. Basic Med. Sci. 2017, 20, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Och, A.; Nowak, R. Barberry (Berberis vulgaris)—Traditional and Contemporary Use. In Medicinal Plants: Domestication, Biotechnology and Regional Importance; Springer: Berlin/Heidelberg, Germany, 2021; pp. 797–825. [Google Scholar]
- Zilaee, M.; Kermany, T.; Tavalaee, S.; Salehi, M.; Ghayour-Mobarhan, M.; Ferns, G.A.A. Barberry treatment reduces serum antiheat shock protein 27 and 60 antibody titres and high-sensitivity C-reactive protein in patients with metabolic syndrome: A double-blind, randomized placebo-controlled trial. Phytother. Res. 2014, 28, 1211–1215. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Sahebkar, A.; Kermani, T.; Zhilaee, M.; Tavallaie, S.; Ghayour Mobarhan, M. Barberry administration and pro-oxidant–antioxidant balance in patients with metabolic syndrome. Iran. Red Crescent Med. J. 2014, 16, e16786. [Google Scholar] [CrossRef]
- Ebrahimi-Mamaghani, M.; Arefhosseini, S.R.; Golzarand, M.; Aliasgarzadeh, A. Long-term effects of processed Berberis vulgaris on some metabolic syndrome components. Iran. J. Endocrinol. Metab. 2009, 11, Pe41–Pe47. [Google Scholar]
- Meliani, N.; Dib, M.A.; Allali, H.; Tabti, B. Hypoglycaemic effect of Berberis vulgaris L. in normal and streptozotocin-induced diabetic rats. Asian Pac. J. Trop. Biomed. 2011, 1, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Safari, Z.; Farrokhzad, A.; Ghavami, A.; Fadel, A.; Hadi, A.; Rafiee, S.; Mokari-Yamchi, A.; Askari, G. The Effect of Barberry (Berberis vulgaris L.) on Glycemic Indices: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Complement. Ther. Med. 2020, 51, 102414. [Google Scholar] [CrossRef] [PubMed]
- Kukula-Koch, W.; Koch, W.; Angelis, A.; Halabalaki, M.; Aligiannis, N. Application of pH-zone refining hydrostatic countercurrent chromatography (hCCC) for the recovery of antioxidant phenolics and the isolation of alkaloids from Siberian barberry herb. Food Chem. 2016, 203, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Olech, M.; Łyko, L. Nowak R, Influence of Accelerated Solvent Extraction Conditions on the LC-ESI-MS/MS Polyphenolic Profile, Triterpenoid Content, and Antioxidant and Anti-lipoxygenase Activity of Rhododendron luteum Sweet Leaves. Antioxidants 2020, 9, 822. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Pieczykolan, A.; Pietrzak, W.; Nowak, R.; Pielczyk, J.; Łamacz, K. Optimization of Extraction Conditions for Determination of Tiliroside in Tilia L, Flowers Using an LC-ESI-MS/MS Method. J. Anal. Methods Chem. 2019, 2019, 9052425. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-Throughput Assay of Oxygen Radical Absorbance Capacity (ORAC) Using a Multichannel Liquid Handling System Coupled with a Microplate Fluorescence Reader in 96-Well Format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Baraniak, B.; Szymanowska, U. Lipooxygenase in food of plant origin. Żywność Nauka Technol. Jakość 2006, 2, 29–45. [Google Scholar]
- Podgórski, R.; Cieśla, M.; Podgórska, D.; Bajorek, W.; Płonka, A.; Czarny, W.; Trybulski, R.; Król, P. Plasma microRNA-320a as a Potential Biomarker of Physiological Changes during Training in Professional Volleyball Players. J. Clin. Med. 2022, 11, 263. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ABTS•+ [mgTE/g] | ORAC [mgTE/g] | DPPH• [mgTE/g] | LOX Inhibition [%] |
|---|---|---|---|
| 122.92 ± 0.01 | 220.29 ± 0.02 | 63.93 ± 0.01 | 62.60 ± 0.87 |
| CCD841 CoN | LS180 | HT-29 | |
|---|---|---|---|
| µg/mL | µg/mL | µg/mL | |
| BVR | 50.21 ± 1.22 | 4.31 ± 1.19 | 46.06 ± 1.11 |
| Berberine | 254.8 ± 2.05 | 0.45 ± 1.25 | 15.92 ± 1.08 |
| Palmatine | 179.8 ± 2.24 | 12.92 ± 1.10 | 29.34 ± 1.08 |
| Berbamine | 13.63 ± 1.02 | 37.62 ± 1.02 | 8.77 ± 1.07 |
| Gene | LS180 | HT29 | ||||
|---|---|---|---|---|---|---|
| Control | Exposed | Change of Expression (%) | Control | Exposed | Change of Expression (%) | |
| CASP3 | 0.46 [0.38–0.51] | 1.14 [1.09–1.17] | 247.83↑ | 0.57 ± 0.1 | 1.01 ± 0.11 | 177.19↑ |
| CASP9 | 0.48 [0.44–0.6] | 1.17 [1.01–1.31] | 243.75↑ | 1.69 ± 0.25 | 2.67 ± 0.38 | 157.99↑ |
| BCL2 | 1.23 ± 0.24 | 0.64 ± 0.12 | 52.03↓ | 0.12 [0.12–0.16] | 0.03 [0.02–0.04] | 25↓ |
| BCL2L1 | 0.42 [0.39–0.57] | 1.77 [1.44–1.86] | 421.43↑ | 5.77 ± 1.12 | 7.32 ± 1.32 | 126.86↑ |
| BCL2L2 | 0.55 [0.42–0.58] | 1.32 [1.28–1.43] | 240↑ | 0.93 [0.68–1.15 | 2.66 [2.55–2.8] | 286.02↑ |
| No | RT (min) | Formula | Error (ppm) | Measured m/z | MS/MS Fragments | CAD Area (%) | Identity | Content [mg eq/g of Dry BVR] ± SD |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.81 | - | 0.0 | 341.1089 a | 179.0580, 119.0366 | 54.62 | - | ui c |
| 2 | 7.28 | C16H19O9 | −0.4 | 355.1036 a | 193.0514, 78.0270, 134.0379 | 1.81 | Feruloyl-hexoside | 7.10 d ± 0.03 |
| 3 | 8.67 | C17H22O10 | −1.3 | 385.1145 a | 223.0624, 79.0699, 164.0471 | 0.22 | Sinapoyl-hexoside | 0.66 d ± 0.00 |
| 4 | 11.46 | C20H24NO4 | 1.1 | 342.1695 b | 265.0854, 97.1117, 282.0882, 65.0694 | 1.89 | Magnoflorine * | 11.63 e ± 0.12 |
| 5 | 12.69 | C19H24NO3 | 0.4 | 314.1749 b | 269.1169, 209.0957, 237.0907, 165.0697 | 0.64 | Unidentified alkaloid | <LLOQ |
| 6 | 14.06 | C36H39N2O6 | 0.6 | 595.2799 b | 595.2798, 564.2381, 552.2376, 367.1648 | 6.04 | Aromoline | 7.11 e ± 0.22 |
| 7 | 17.16 | C37H40N2O6 | 0.9 | 609.2954 b | 609.2955, 578.2539, 566.2538, 381.1804 | 2.46 | Berbamine * | 3.19 e ± 0.04 |
| 8 | 18.61 | C37H40N2O6 | 0.4 | 609.2957 b | 609.2955, 381.1807, 174.0913, 578.2532 | 3.85 | Oxycanthine | 5.48 e ± 0.10 |
| 9 | 20.27 | C20H20NO4 | −0.2 | 338.1381 b | 322.1077, 308.0919, 294.1126, 236.0709 | 0.44 | Columbamine | 3.32 e ± 0.08 |
| 10 | 20.53 | C20H20NO4 | −0.5 | 338.1389 b | 322.1079,308.0920, 294.1128, 236.0708 | 4.4 | Jatrorrhizine * | 23.32 e ± 0.11 |
| 11 | 21.35 | C38H43N2O6 | 0.9 | 623.3110 b | 623.3108, 381.1810, 174.0912, 592.2681 | 0.22 | Rodiasine | <LLOQ |
| 12 | 22.73 | C20H18NO4 | 0.3 | 336.1229 b | 320.0916, 292.0966, 278.0809, 306.0760 | 19.34 | Berberine * | 70.27 ± 0.48 |
| 13 | 22.97 | C21H22NO4 | 1.4 | 352.1537 b | 336.1226, 278.0807, 322.1073, 308.1278 | 4.07 | Palmatine * | 7.57 e ± 0.30 |
| Metabolite of the Root of B. vulgaris L. | Biological Activity of BVR Metabolite and Prospective of Further Investigation | References | Literature Confirmation of B. vulgaris L. Extract Activity | References |
|---|---|---|---|---|
| Bersavine Berberine Berbanine Berbamine | Antileukemic | [36,61,62,63,64,65,66] | Yes | [66,67] |
| Bersavine Berbamine Berberine Magnoflorine | Cervix cancer | [59,61,62,68] | No | - |
| Jatrorrhizine Berberine Magnoflorine | Liver cancer | [39,46,59,69,70,71,72] | Yes | [73] |
| Jatrorrhizine Berberine Bersavine Berbamine Magnoflorine Palmatine N-(p-trans-coumaroyl)tyramine cannabisin G (±)-lyoniresinol | Breast cancer | [46,60,61,62,69,74,75,76] | Yes | [21,77] |
| Berberine Jatrorrhizine Palmatine Columbamine Berbamine Bersavine | Colon cancer | [38,41,42,43,44,47,53,53,57,61,62,78] | No | - |
| Magnoflorine Berberine | Gastric cancer | [58,79] | No | - |
| Palmatine Berberine Jatrorrhizine | Skin cancer | [51,54,55,80] | No | - |
| Berberine, Columbamine, Isocorydine, Oxyberberine | Prostate cancer | [81] | No | - |
| Bersavine Berbamine Berberine | Lung cancer | [79,81,82] | No | - |
| (+)-chenabinol Berkristine Verfilline Bersavine Muraricine Berbostrejdine Aromoline Berlambine Obamegine Berberine Jathrrorhizine | Alzheimer disease | [48,83,84,85,86,87,88,89,90,91] | No | - |
| Berberine Magnoflorine Palmatine | Anti-inflammatory | [11,12,56,92] | Yes | [13,15,16,17,24,30,93] |
| Berberine Palmatine Jathrrorhizine | Anti-diabetic and Metabolic syndrome | [50,81,92,94] | Yes | [10,95,96,97,98,99,100,101] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Och, A.; Lemieszek, M.K.; Cieśla, M.; Jedrejek, D.; Kozłowska, A.; Pawelec, S.; Nowak, R. Berberis vulgaris L. Root Extract as a Multi-Target Chemopreventive Agent against Colon Cancer Causing Apoptosis in Human Colon Adenocarcinoma Cell Lines. Int. J. Mol. Sci. 2024, 25, 4786. https://doi.org/10.3390/ijms25094786
Och A, Lemieszek MK, Cieśla M, Jedrejek D, Kozłowska A, Pawelec S, Nowak R. Berberis vulgaris L. Root Extract as a Multi-Target Chemopreventive Agent against Colon Cancer Causing Apoptosis in Human Colon Adenocarcinoma Cell Lines. International Journal of Molecular Sciences. 2024; 25(9):4786. https://doi.org/10.3390/ijms25094786
Chicago/Turabian StyleOch, Anna, Marta Kinga Lemieszek, Marek Cieśla, Dariusz Jedrejek, Aleksandra Kozłowska, Sylwia Pawelec, and Renata Nowak. 2024. "Berberis vulgaris L. Root Extract as a Multi-Target Chemopreventive Agent against Colon Cancer Causing Apoptosis in Human Colon Adenocarcinoma Cell Lines" International Journal of Molecular Sciences 25, no. 9: 4786. https://doi.org/10.3390/ijms25094786
APA StyleOch, A., Lemieszek, M. K., Cieśla, M., Jedrejek, D., Kozłowska, A., Pawelec, S., & Nowak, R. (2024). Berberis vulgaris L. Root Extract as a Multi-Target Chemopreventive Agent against Colon Cancer Causing Apoptosis in Human Colon Adenocarcinoma Cell Lines. International Journal of Molecular Sciences, 25(9), 4786. https://doi.org/10.3390/ijms25094786