
Loss of CHOP Prevents Joint Degeneration and Pain in a Mouse Model of Pseudoachondroplasia

 , , , , and
, , , , and
Abstract
:
1. Introduction
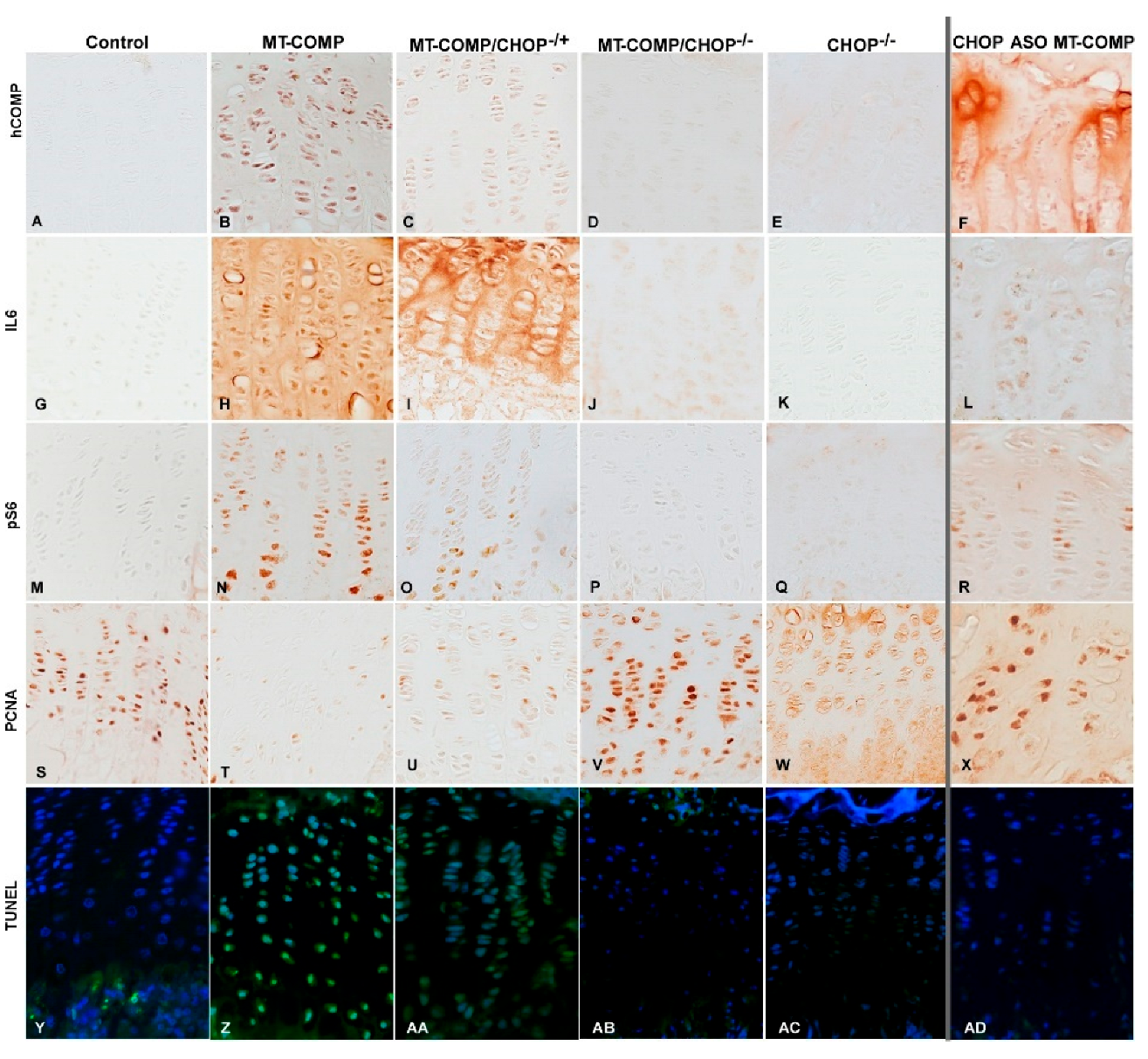
2. Results
2.1. Loss of CHOP in MT-COMP Mice Does Not Improve Femur Length
2.2. Reduction of CHOP Partially Normalizes COMP Localization in the Growth Plate
2.3. Joint Health Is Preserved with Loss of CHOP
2.4. Pain Mitigation Is Associated with the Absence or Reduction of CHOP in MT-COMP Mice
2.5. Articular Chondrocyte Death and Markers of Pathologic Processes That Lead to Cell Death Are Reduced in the Absence or Reduction of CHOP
2.6. Loss or Reduction of CHOP in MT-COMP Lessens Inflammation and Downstream Molecules in Articular Chondrocytes
3. Discussion
4. Materials and Methods
4.1. Bigenic Mice
4.2. Mutant COMP Induction
4.3. CHOP ASO Treatment
4.4. Immunohistochemistry
4.5. Limb Length Measurements
4.6. Grooming Assessments
4.7. Joint Degeneration Scoring
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halasz, K.; Kassner, A.; Morgelin, M.; Heinegard, D. COMP acts as a catalyst in collagen fibrillogenesis. J. Biol. Chem. 2007, 282, 31166–31173. [Google Scholar] [CrossRef] [PubMed]
- Merritt, T.M.; Bick, R.; Poindexter, B.J.; Alcorn, J.L.; Hecht, J.T. Unique matrix structure in the rough endoplasmic reticulum cisternae of pseudoachondroplasia chondrocytes. Am. J. Pathol. 2007, 170, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Thur, J.; Rosenberg, K.; Nitsche, D.P.; Pihlajamaa, T.; Ala-Kokko, L.; Heinegard, D.; Paulsson, M.; Maurer, P. Mutations in cartilage oligomeric matrix protein causing pseudoachondroplasia and multiple epiphyseal dysplasia affect binding of calcium and collagen I, II, and IX. J. Biol. Chem. 2001, 276, 6083–6092. [Google Scholar] [CrossRef] [PubMed]
- Bleasel, J.F.; Poole, A.R.; Heinegard, D.; Saxne, T.; Holderbaum, D.; Ionescu, M.; Jones, P.; Moskowitz, R.W. Changes in serum cartilage marker levels indicate altered cartilage metabolism in families with the osteoarthritis-related type II collagen gene COL2A1 mutation. Arthritis Rheum. 1999, 42, 39–45. [Google Scholar] [CrossRef]
- Lohmander, L.S.; Saxne, T.; Heinegard, D.K. Release of cartilage oligomeric matrix protein (COMP) into joint fluid after knee injury and in osteoarthritis. Ann. Rheum. Dis. 1994, 53, 8–13. [Google Scholar] [CrossRef]
- Mann, H.H.; Ozbek, S.; Engel, J.; Paulsson, M.; Wagener, R. Interactions between the cartilage oligomeric matrix protein and matrilins. Implications for matrix assembly and the pathogenesis of chondrodysplasias. J. Biol. Chem. 2004, 279, 25294–25298. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, Y.; Ilalov, K.; Carlson, C.S.; Feng, J.Q.; Di Cesare, P.E.; Liu, C.J. Cartilage oligomeric matrix protein associates with granulin-epithelin precursor (GEP) and potentiates GEP-stimulated chondrocyte proliferation. J. Biol. Chem. 2007, 282, 11347–11355. [Google Scholar] [CrossRef]
- Chen, F.H.; Thomas, A.O.; Hecht, J.T.; Goldring, M.B.; Lawler, J. Cartilage oligomeric matrix protein/thrombospondin 5 supports chondrocyte attachment through interaction with integrins. J. Biol. Chem. 2005, 280, 32655–32661. [Google Scholar] [CrossRef]
- Unger, S.; Hecht, J.T. Pseudoachondroplasia and multiple epiphyseal dysplasia: New etiologic developments. Am. J. Med. Genet. 2001, 106, 244–250. [Google Scholar] [CrossRef]
- Posey, K.L.; Hayes, E.; Haynes, R.; Hecht, J.T. Role of TSP-5/COMP in pseudoachondroplasia. Int. J. Biochem. Cell Biol. 2004, 36, 1005–1012. [Google Scholar] [CrossRef]
- Posey, K.L.; Hecht, J.T. The role of cartilage oligomeric matrix protein (COMP) in skeletal disease. Curr. Drug Targets 2008, 9, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Posey, K.L.; Coustry, F.; Hecht, J.T. Cartilage oligomeric matrix protein: COMPopathies and beyond. Matrix Biol. 2018, 71–72, 161–173. [Google Scholar] [CrossRef] [PubMed]
- McKeand, J.; Rotta, J.; Hecht, J.T. Natural history study of pseudoachondroplasia. Am. J. Med. Genet. 1996, 63, 406–410. [Google Scholar] [CrossRef]
- Deere, M.; Sanford, T.; Ferguson, H.L.; Daniels, K.; Hecht, J.T. Identification of twelve mutations in cartilage oligomeric matrix protein (COMP) in patients with pseudoachondroplasia. Am. J. Med. Genet. 1998, 80, 510–513. [Google Scholar] [CrossRef]
- Deere, M.; Sanford, T.; Francomano, C.A.; Daniels, K.; Hecht, J.T. Identification of nine novel mutations in cartilage oligomeric matrix protein in patients with pseudoachondroplasia and multiple epiphyseal dysplasia. Am. J. Med. Genet. 1999, 85, 486–490. [Google Scholar] [CrossRef]
- Hecht, J.T.; Veerisetty, A.C.; Hossain, M.G.; Patra, D.; Chiu, F.; Coustry, F.; Posey, K.L. Joint Degeneration in a Mouse Model of Pseudoachondroplasia: ER Stress, Inflammation, and Block of Autophagy. Int. J. Mol. Sci. 2021, 22, 9239. [Google Scholar] [CrossRef]
- Posey, K.L.; Coustry, F.; Veerisetty, A.C.; Hossain, M.; Alcorn, J.L.; Hecht, J.T. Antioxidant and anti-inflammatory agents mitigate pathology in a mouse model of pseudoachondroplasia. Hum. Mol. Genet. 2015, 24, 3918–3928. [Google Scholar] [CrossRef]
- Posey, K.L.; Veerisetty, A.C.; Liu, P.; Wang, H.R.; Poindexter, B.J.; Bick, R.; Alcorn, J.L.; Hecht, J.T. An inducible cartilage oligomeric matrix protein mouse model recapitulates human pseudoachondroplasia phenotype. Am. J. Pathol. 2009, 175, 1555–1563. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef]
- Gotoh, T.; Endo, M.; Oike, Y. Endoplasmic reticulum stress-related inflammation and cardiovascular diseases. Int. J. Inflam. 2011, 2011, 259462. [Google Scholar] [CrossRef]
- Salminen, A.; Kauppinen, A.; Suuronen, T.; Kaarniranta, K.; Ojala, J. ER stress in Alzheimer’s disease: A novel neuronal trigger for inflammation and Alzheimer’s pathology. J. Neuroinflamm. 2009, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, S.B.; Diehl, J.A. Coordination of ER and oxidative stress signaling: The PERK/Nrf2 signaling pathway. Int. J. Biochem. Cell Biol. 2006, 38, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [PubMed]
- Posey, K.L.; Coustry, F.; Veerisetty, A.C.; Hossain, M.G.; Gambello, M.J.; Hecht, J.T. Novel mTORC1 Mechanism Suggests Therapeutic Targets for COMPopathies. Am. J. Pathol. 2019, 189, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.T.; Coustry, F.; Veerisetty, A.C.; Hossain, M.G.; Posey, K.L. Resveratrol Reduces COMPopathy in Mice Through Activation of Autophagy. J. Bone Miner. Res. Plus 2021, 5, e10456. [Google Scholar] [CrossRef]
- Hecht, J.T.; Makitie, O.; Hayes, E.; Haynes, R.; Susic, M.; Montufar-Solis, D.; Duke, P.J.; Cole, W.G. Chondrocyte cell death and intracellular distribution of COMP and type IX collagen in the pseudoachondroplasia growth plate. J. Orthop. Res. 2004, 22, 759–767. [Google Scholar] [CrossRef]
- Hecht, J.T.; Montufar-Solis, D.; Decker, G.; Lawler, J.; Daniels, K.; Duke, P.J. Retention of cartilage oligomeric matrix protein (COMP) and cell death in redifferentiated pseudoachondroplasia chondrocytes. Matrix Biol. 1998, 17, 625–633. [Google Scholar] [CrossRef]
- Merritt, T.M.; Alcorn, J.L.; Haynes, R.; Hecht, J.T. Expression of mutant cartilage oligomeric matrix protein in human chondrocytes induces the pseudoachondroplasia phenotype. J. Orthop. Res. 2006, 24, 700–707. [Google Scholar] [CrossRef]
- Cooper, R.R.; Ponseti, I.V.; Maynard, J.A. Pseudoachondroplasia dwarfism. A rough-surfaced endoplasmic reticulum disorder. J. Bone Jt. Surg. Am. 1973, 55A, 475–484. [Google Scholar] [CrossRef]
- Posey, K.L.; Alcorn, J.L.; Hecht, J.T. Pseudoachondroplasia/COMP-translating from the bench to the bedside. Matrix Biol. 2014, 37, 167–173. [Google Scholar] [CrossRef]
- Posey, K.L.; Coustry, F.; Veerisetty, A.C.; Liu, P.; Alcorn, J.L.; Hecht, J.T. Chop (Ddit3) is essential for D469del-COMP retention and cell death in chondrocytes in an inducible transgenic mouse model of pseudoachondroplasia. Am. J. Pathol. 2012, 180, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Posey, K.L.; Coustry, F.; Veerisetty, A.C.; Liu, P.; Alcorn, J.L.; Hecht, J.T. Chondrocyte-specific pathology during skeletal growth and therapeutics in a murine model of pseudoachondroplasia. J. Bone Miner. Res. 2014, 29, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Posey, K.L.; Hecht, J.T. Novel therapeutic interventions for pseudoachondroplasia. Bone 2017, 102, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, R.J. Stress signaling from the lumen of the endoplasmic reticulum: Coordination of gene transcriptional and translational controls. Genes Dev. 1999, 13, 1211–1233. [Google Scholar] [CrossRef] [PubMed]
- Coustry, F.; Posey, K.L.; Maerz, T.; Baker, K.; Abraham, A.M.; Ambrose, C.G.; Nobakhti, S.; Shefelbine, S.J.; Bi, X.; Newton, M.; et al. Mutant cartilage oligomeric matrix protein (COMP) compromises bone integrity, joint function and the balance between adipogenesis and osteogenesis. Matrix Biol. 2018, 67, 75–89. [Google Scholar] [CrossRef]
- Yong, J.; Parekh, V.S.; Reilly, S.M.; Nayak, J.; Chen, Z.; Lebeaupin, C.; Jang, I.; Zhang, J.; Prakash, T.P.; Sun, H.; et al. Chop/Ddit3 depletion in beta cells alleviates ER stress and corrects hepatic steatosis in mice. Sci. Transl. Med. 2021, 13, eaba9796. [Google Scholar] [CrossRef]
- Glasson, S.S.; Chambers, M.G.; Van Den Berg, W.B.; Little, C.B. The OARSI histopathology initiative-recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18, S17–S23. [Google Scholar] [CrossRef]
- Hecht, J.T.; Veerisetty, A.C.; Patra, D.; Hossain, M.G.; Chiu, F.; Mobed, C.; Gannon, F.H.; Posey, K.L. Early Resveratrol Treatment Mitigates Joint Degeneration and Dampens Pain in a Mouse Model of Pseudoachondroplasia (PSACH). Biomolecules 2023, 13, 1553. [Google Scholar] [CrossRef]
- Husa, M.; Petursson, F.; Lotz, M.; Terkeltaub, R.; Liu-Bryan, R. C/EBP homologous protein drives pro-catabolic responses in chondrocytes. Arthritis Res. Ther. 2013, 15, R218. [Google Scholar] [CrossRef]
- Hu, H.; Tian, M.; Ding, C.; Yu, S. The C/EBP Homologous Protein (CHOP) Transcription Factor Functions in Endoplasmic Reticulum Stress-Induced Apoptosis and Microbial Infection. Front. Immunol. 2018, 9, 3083. [Google Scholar] [CrossRef]
- Yang, C.; Dong, W.; Wang, Y.; Dong, X.; Xu, X.; Yu, X.; Wang, J. DDIT3 aggravates TMJOA cartilage degradation via Nrf2/HO-1/NLRP3-mediated autophagy. Osteoarthr. Cartil. 2024, 32, 921–937. [Google Scholar] [CrossRef] [PubMed]
- Uehara, Y.; Hirose, J.; Yamabe, S.; Okamoto, N.; Okada, T.; Oyadomari, S.; Mizuta, H. Endoplasmic reticulum stress-induced apoptosis contributes to articular cartilage degeneration via C/EBP homologous protein. Osteoarthr. Cartil. 2014, 22, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.T.; Veerisetty, A.C.; Hossain, M.G.; Chiu, F.; Posey, K.L. CurQ+, a Next-Generation Formulation of Curcumin, Ameliorates Growth Plate Chondrocyte Stress and Increases Limb Growth in a Mouse Model of Pseudoachondroplasia. Int. J. Mol. Sci. 2023, 24, 3845. [Google Scholar] [CrossRef] [PubMed]
- Coustry, F.; Posey, K.L.; Liu, P.; Alcorn, J.L.; Hecht, J.T. D469del-COMP retention in chondrocytes stimulates caspase-independent necroptosis. Am. J. Pathol. 2012, 180, 738–748. [Google Scholar] [CrossRef]
- Posey, K.L.; Coustry, F.; Veerisetty, A.C.; Hossain, M.; Gattis, D.; Booten, S.; Alcorn, J.L.; Seth, P.P.; Hecht, J.T. Antisense Reduction of Mutant COMP Reduces Growth Plate Chondrocyte Pathology. Mol. Ther. 2017, 25, 705–714. [Google Scholar] [CrossRef]
- Perez-Arancibia, R.; Rivas, A.; Hetz, C. (off)Targeting UPR signaling: The race toward intervening ER proteostasis. Expert Opin. Ther. Targets 2018, 22, 97–100. [Google Scholar] [CrossRef]
- Linnane, E.; Davey, P.; Zhang, P.; Puri, S.; Edbrooke, M.; Chiarparin, E.; Revenko, A.S.; Macleod, A.R.; Norman, J.C.; Ross, S.J. Differential uptake, kinetics and mechanisms of intracellular trafficking of next-generation antisense oligonucleotides across human cancer cell lines. Nucleic Acids Res. 2019, 47, 4375–4392. [Google Scholar] [CrossRef]
- Crooke, S.T.; Liang, X.H.; Baker, B.F.; Crooke, R.M. Antisense technology: A review. J. Biol. Chem. 2021, 296, 100416. [Google Scholar] [CrossRef]
- Doxtader Lacy, K.A.; Liang, X.H.; Zhang, L.; Crooke, S.T. RNA modifications can affect RNase H1-mediated PS-ASO activity. Mol. Ther. Nucleic Acids 2022, 28, 814–828. [Google Scholar] [CrossRef]
- Hunziker, E.B. Growth plate structure and function. Pathol. Immunopathol. Res. 1988, 7, 9–13. [Google Scholar] [CrossRef]
- Gamble, C.; Nguyen, J.; Hashmi, S.S.; Hecht, J.T. Pseudoachondroplasia and painful sequelae. Am. J. Med. Genet. A 2015, 167, 2618–2622. [Google Scholar] [CrossRef] [PubMed]
- Deuis, J.R.; Dvorakova, L.S.; Vetter, I. Methods Used to Evaluate Pain Behaviors in Rodents. Front. Mol. Neurosci. 2017, 10, 284. [Google Scholar] [CrossRef] [PubMed]
- Eddy, N.B.; Leimbach, D. Synthetic analgesics. II. Dithienylbutenyl- and dithienylbutylamines. J. Pharmacol. Exp. Ther. 1953, 107, 385–393. [Google Scholar] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Dvir-Ginzberg, M.; Mobasheri, A.; Kumar, A. The Role of Sirtuins in Cartilage Homeostasis and Osteoarthritis. Curr. Rheumatol. Rep. 2016, 18, 43. [Google Scholar] [CrossRef]
- Kim, K.H.; Jo, J.H.; Cho, H.J.; Park, T.S.; Kim, T.M. Therapeutic potential of stem cell-derived extracellular vesicles in osteoarthritis: Preclinical study findings. Lab. Anim. Res. 2020, 36, 10. [Google Scholar] [CrossRef]
- Hamamura, K.; Nishimura, A.; Iino, T.; Takigawa, S.; Sudo, A.; Yokota, H. Chondroprotective effects of Salubrinal in a mouse model of osteoarthritis. Bone Jt. Res. 2015, 4, 84–92. [Google Scholar] [CrossRef]
- Mehana, E.E.; Khafaga, A.F.; El-Blehi, S.S. The role of matrix metalloproteinases in osteoarthritis pathogenesis: An updated review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef]
- Yu, M.; Yi, S.Q.; Wu, Y.R.; Sun, H.L.; Song, F.F.; Wang, J.W. Ddit3 suppresses the differentiation of mouse chondroprogenitor cells. Int. J. Biochem. Cell Biol. 2016, 81, 156–163. [Google Scholar] [CrossRef]
- Wang, C.; Tan, Z.; Niu, B.; Tsang, K.Y.; Tai, A.; Chan, W.C.W.; Lo, R.L.K.; Leung, K.K.H.; Dung, N.W.F.; Itoh, N.; et al. Inhibiting the integrated stress response pathway prevents aberrant chondrocyte differentiation thereby alleviating chondrodysplasia. Elife 2018, 7, e37673. [Google Scholar] [CrossRef]
- Posey, K.L.; Yang, Y.; Veerisetty, A.C.; Sharan, S.K.; Hecht, J.T. Model systems for studying skeletal dysplasias caused by TSP-5/COMP mutations. Cell. Mol. Life Sci. 2008, 65, 687–699. [Google Scholar] [CrossRef]
- Hecht, J.T.; Veerisetty, A.C.; Wu, J.; Coustry, F.; Hossain, M.G.; Chiu, F.; Gannon, F.H.; Posey, K.L. Primary Osteoarthritis Early Joint Degeneration Induced by Endoplasmic Reticulum Stress Is Mitigated by Resveratrol. Am. J. Pathol. 2021, 191, 1624–1637. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | p Value at Time Points in Weeks (wks) | ||||||
|---|---|---|---|---|---|---|---|
| 8 wk | 12 wk | 16 wk | 20 wk | 24 wk | 30 wk | 36 wk | |
| Control vs. MT-COMP | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Control vs. MT-COMP/CHOP−/− | 0.292 | 0.716 | 0.408 | 0.056 | 0.598 | 1.000 | 0.814 |
| Control vs. MT-COMP/CHOP−/+ | 0.079 | 0.999 | 0.111 | 0.294 | - | - | - |
| MT-COMP vs. Control | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| MT-COMP vs. MT-COMP/CHOP−/− | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.003 |
| MT-COMP vs. MT-COMP/CHOP−/+ | <0.001 | <0.001 | <0.001 | 0.007 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hecht, J.T.; Veerisetty, A.C.; Hossain, M.G.; Patra, D.; Carrer, M.; Chiu, F.; Relic, D.; Jafar-nejad, P.; Posey, K.L. Loss of CHOP Prevents Joint Degeneration and Pain in a Mouse Model of Pseudoachondroplasia. Int. J. Mol. Sci. 2025, 26, 16. https://doi.org/10.3390/ijms26010016
Hecht JT, Veerisetty AC, Hossain MG, Patra D, Carrer M, Chiu F, Relic D, Jafar-nejad P, Posey KL. Loss of CHOP Prevents Joint Degeneration and Pain in a Mouse Model of Pseudoachondroplasia. International Journal of Molecular Sciences. 2025; 26(1):16. https://doi.org/10.3390/ijms26010016
Chicago/Turabian StyleHecht, Jacqueline T., Alka C. Veerisetty, Mohammad G. Hossain, Debabrata Patra, Michele Carrer, Frankie Chiu, Dorde Relic, Paymaan Jafar-nejad, and Karen L. Posey. 2025. "Loss of CHOP Prevents Joint Degeneration and Pain in a Mouse Model of Pseudoachondroplasia" International Journal of Molecular Sciences 26, no. 1: 16. https://doi.org/10.3390/ijms26010016
APA StyleHecht, J. T., Veerisetty, A. C., Hossain, M. G., Patra, D., Carrer, M., Chiu, F., Relic, D., Jafar-nejad, P., & Posey, K. L. (2025). Loss of CHOP Prevents Joint Degeneration and Pain in a Mouse Model of Pseudoachondroplasia. International Journal of Molecular Sciences, 26(1), 16. https://doi.org/10.3390/ijms26010016