
Systems Modeling Reveals Shared Metabolic Dysregulation and Potential Treatments in ME/CFS and Long COVID

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
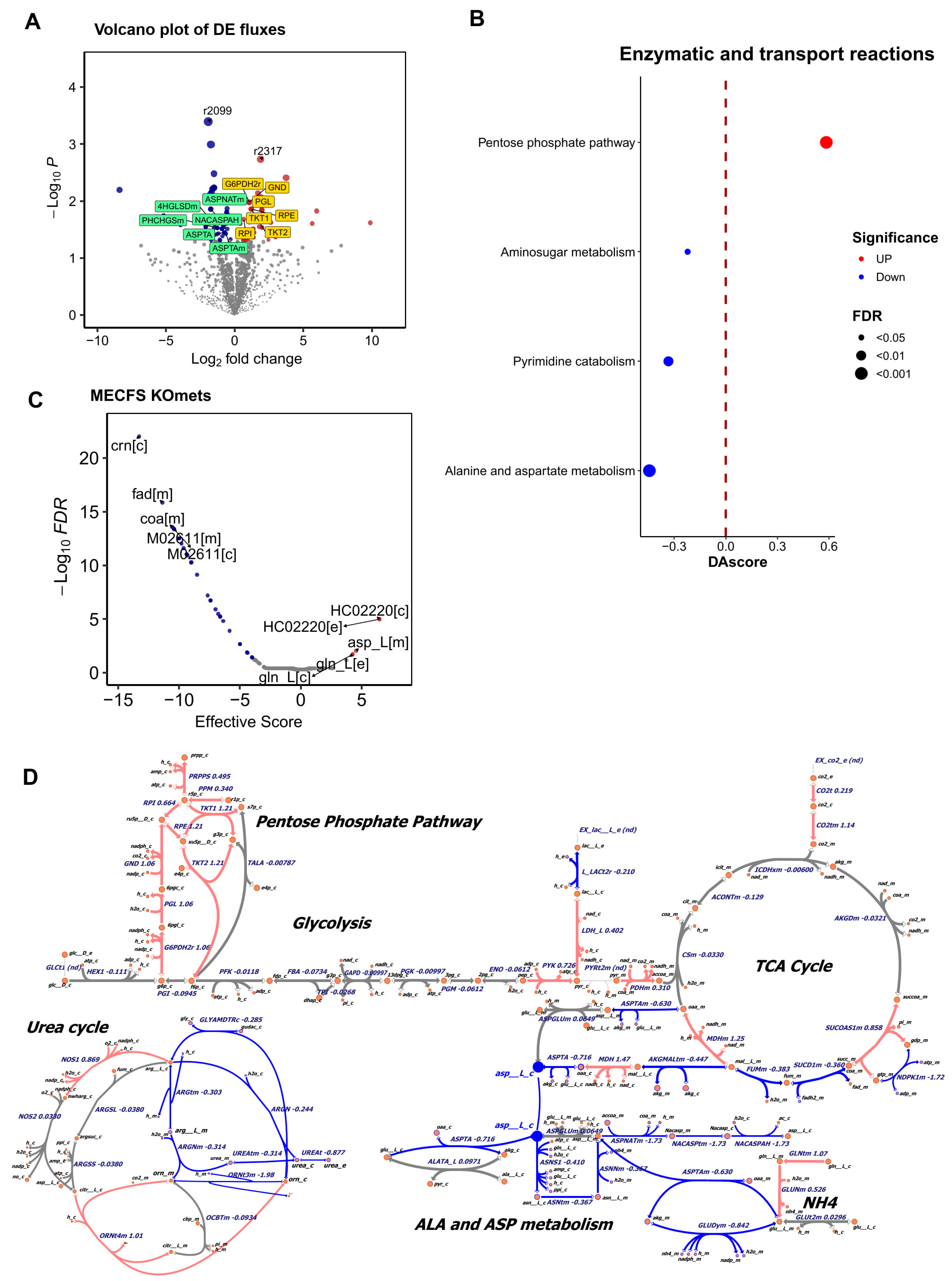
2.1. Metabolic Modeling Reveals Altered Metabolism in the Muscle of ME/CFS Patients
2.2. Metabolomics Measurements of Long COVID Reveals Down-Regulated Asparagine (ASN) During PEM in Muscle and the Blood
3. Discussion
4. Materials and Methods
4.1. Dataset Collection
4.2. Genome-Wide Precision Modeling of Metabolic Fluxes in the Muscle of Patients and Controls
4.2.1. Metabolic Modeling
4.2.2. Identification of Significantly Changed Fluxes
4.2.3. Identification of Significantly Changed Metabolic Pathways
4.2.4. All-Against-All Knockout Analysis
4.3. Metabolomics Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Board on the Health of Select Populations, Committee on the Diagnostic Criteria for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Beyond Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: Redefining an Illness; National Academies Press: Washington, DC, USA, 2015. [Google Scholar] [CrossRef]
- Cortes Rivera, M.; Mastronardi, C.; Silva-Aldana, C.T.; Arcos-Burgos, M.; Lidbury, B.A. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: A Comprehensive Review. Diagnostics 2019, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Valdez, A.R.; Hancock, E.E.; Adebayo, S.; Kiernicki, D.J.; Proskauer, D.; Attewell, J.R.; Bateman, L.; DeMaria, A., Jr.; Lapp, C.W.; Rowe, P.C.; et al. Estimating Prevalence, Demographics, and Costs of ME/CFS Using Large Scale Medical Claims Data and Machine Learning. Front. Pediatr. 2018, 6, 412. [Google Scholar] [CrossRef]
- Ladds, E.; Darbyshire, J.L.; Bakerly, N.D.; Falope, Z.; Tucker-Bell, I. Cognitive dysfunction after covid-19. BMJ 2024, 384, e075387. [Google Scholar] [CrossRef] [PubMed]
- Thaweethai, T.; Jolley, S.E.; Karlson, E.W.; Levitan, E.B.; Levy, B.; McComsey, G.A.; McCorkell, L.; Nadkarni, G.N.; Parthasarathy, S.; Singh, U.; et al. Development of a Definition of Postacute Sequelae of SARS-CoV-2 Infection. JAMA 2023, 329, 1934–1946. [Google Scholar] [CrossRef]
- Komaroff, A.L.; Lipkin, W.I. ME/CFS and Long COVID share similar symptoms and biological abnormalities: Road map to the literature. Front. Med. 2023, 10, 1187163. [Google Scholar] [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine; Health and Medicine Division; Board on Health Sciences Policy; Board on Global Health; Forum on Neuroscience and Nervous System Disorders; Forum on Microbial Threats. Toward a Common Research Agenda in Infection-Associated Chronic Illnesses: Proceedings of a Workshop; Snair, M., Liao, J., Ashby, E., Biffl, C., Eds.; The National Academies Collection: Washington, DC, USA, 2024. [Google Scholar]
- Saito, S.; Shahbaz, S.; Luo, X.; Osman, M.; Redmond, D.; Cohen Tervaert, J.W.; Li, L.; Elahi, S. Metabolomic and immune alterations in long COVID patients with chronic fatigue syndrome. Front. Immunol. 2024, 15, 1341843. [Google Scholar] [CrossRef] [PubMed]
- Domingo, J.C.; Battistini, F.; Cordobilla, B.; Zaragoza, M.C.; Sanmartin-Sentanes, R.; Alegre-Martin, J.; Cambras, T.; Castro-Marrero, J. Association of circulating biomarkers with illness severity measures differentiates myalgic encephalomyelitis/chronic fatigue syndrome and post-COVID-19 condition: A prospective pilot cohort study. J. Transl. Med. 2024, 22, 343. [Google Scholar] [CrossRef]
- Naviaux, R.K.; Naviaux, J.C.; Li, K.; Bright, A.T.; Alaynick, W.A.; Wang, L.; Baxter, A.; Nathan, N.; Anderson, W.; Gordon, E. Metabolic features of chronic fatigue syndrome. Proc. Natl. Acad. Sci. USA 2016, 113, E5472–E5480. [Google Scholar] [CrossRef]
- Rutherford, G.; Manning, P.; Newton, J.L. Understanding Muscle Dysfunction in Chronic Fatigue Syndrome. J. Aging. Res. 2016, 2016, 2497348. [Google Scholar] [CrossRef]
- Huth, T.K.; Eaton-Fitch, N.; Staines, D.; Marshall-Gradisnik, S. A systematic review of metabolomic dysregulation in Chronic Fatigue Syndrome/Myalgic Encephalomyelitis/Systemic Exertion Intolerance Disease (CFS/ME/SEID). J. Transl. Med. 2020, 18, 198. [Google Scholar] [CrossRef]
- Hoel, F.; Hoel, A.; Pettersen, I.K.; Rekeland, I.G.; Risa, K.; Alme, K.; Sorland, K.; Fossa, A.; Lien, K.; Herder, I.; et al. A map of metabolic phenotypes in patients with myalgic encephalomyelitis/chronic fatigue syndrome. JCI Insight 2021, 6, e149217. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Zhang, J.; Heller, M.; Later, W.; Heymsfield, S.B.; Muller, M.J. Evaluation of specific metabolic rates of major organs and tissues: Comparison between men and women. Am. J. Hum. Biol. 2011, 23, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Walitt, B.; Singh, K.; LaMunion, S.R.; Hallett, M.; Jacobson, S.; Chen, K.; Enose-Akahata, Y.; Apps, R.; Barb, J.J.; Bedard, P.; et al. Deep phenotyping of post-infectious myalgic encephalomyelitis/chronic fatigue syndrome. Nat. Commun. 2024, 15, 907. [Google Scholar] [CrossRef] [PubMed]
- Appelman, B.; Charlton, B.T.; Goulding, R.P.; Kerkhoff, T.J.; Breedveld, E.A.; Noort, W.; Offringa, C.; Bloemers, F.W.; van Weeghel, M.; Schomakers, B.V.; et al. Muscle abnormalities worsen after post-exertional malaise in long COVID. Nat. Commun. 2024, 15, 17. [Google Scholar] [CrossRef]
- Orth, J.D.; Thiele, I.; Palsson, B.O. What is flux balance analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef]
- Heirendt, L.; Arreckx, S.; Pfau, T.; Mendoza, S.N.; Richelle, A.; Heinken, A.; Haraldsdóttir, H.S.; Wachowiak, J.; Keating, S.M.; Vlasov, V. Creation and analysis of biochemical constraint-based models using the COBRA Toolbox v. 3.0. Nat. Protoc. 2019, 14, 639–702. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Tseyang, L.T.; Giese, G.E.; Wang, H.; Yao, B.; Zhang, J.; Neve, R.L.; Shank, E.A.; Spinelli, J.B.; et al. A systems-level, semi-quantitative landscape of metabolic flux in C. elegans. Nature 2025, 640, 194–202. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Boren, J.; Smith, U.; Uhlen, M.; Nielsen, J. Systems biology in hepatology: Approaches and applications. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 365–377. [Google Scholar] [CrossRef]
- Li, G.H.; Li, Y.H.; Yu, Q.; Zhou, Q.Q.; Zhang, R.F.; Weng, C.J.; Ge, M.X.; Kong, Q.P. Unraveling the metabolic heterogeneity and commonality in senescent cells using systems modeling. Life Med. 2025, 4, lnaf003. [Google Scholar] [CrossRef]
- Shen, Q.; Yang, H.; Kong, Q.P.; Li, G.H.; Li, L. Metabolic Modeling Identifies a Novel Molecular Type of Glioblastoma Associated with Good Prognosis. Metabolites 2023, 13, 172. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Palsson, B.O. Genome-scale models in human metabologenomics. Nat. Rev. Genet. 2025, 26, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Han, F.; Xiao, F.H.; Gu, K.S.Y.; Shen, Q.; Xu, W.; Li, W.X.; Wang, Y.L.; Liang, B.; Huang, J.F. System-level metabolic modeling facilitates unveiling metabolic signature in exceptional longevity. Aging Cell 2022, 21, e13595. [Google Scholar] [CrossRef] [PubMed]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef]
- Wang, P.Y.; Ma, J.; Kim, Y.C.; Son, A.Y.; Syed, A.M.; Liu, C.; Mori, M.P.; Huffstutler, R.D.; Stolinski, J.L.; Talagala, S.L.; et al. WASF3 disrupts mitochondrial respiration and may mediate exercise intolerance in myalgic encephalomyelitis/chronic fatigue syndrome. Proc. Natl. Acad. Sci. USA 2023, 120, e2302738120. [Google Scholar] [CrossRef] [PubMed]
- Holecek, M. Aspartic Acid in Health and Disease. Nutrients 2023, 15, 4023. [Google Scholar] [CrossRef]
- Butterworth, R.F.; McPhail, M.J.W. L-Ornithine L-Aspartate (LOLA) for Hepatic Encephalopathy in Cirrhosis: Results of Randomized Controlled Trials and Meta-Analyses. Drugs 2019, 79 (Suppl. 1), 31–37. [Google Scholar] [CrossRef]
- Canbay, A.; Sowa, J.-P. L-Ornithine L-Aspartate (LOLA) as a novel approach for therapy of non-alcoholic fatty liver disease. Drugs 2019, 79 (Suppl. 1), 39–44. [Google Scholar] [CrossRef]
- Kokubo, T.; Ikeshima, E.; Kirisako, T.; Miura, Y.; Horiuchi, M.; Tsuda, A. A randomized, double-masked, placebo-controlled crossover trial on the effects of L-ornithine on salivary cortisol and feelings of fatigue of flushers the morning after alcohol consumption. Biopsychosoc. Med. 2013, 7, 6. [Google Scholar] [CrossRef]
- Sugino, T.; Shirai, T.; Kajimoto, Y.; Kajimoto, O. L-ornithine supplementation attenuates physical fatigue in healthy volunteers by modulating lipid and amino acid metabolism. Nutr. Res. 2008, 28, 738–743. [Google Scholar] [CrossRef]
- Sisalli, M.J.; Della Notte, S.; Secondo, A.; Ventra, C.; Annunziato, L.; Scorziello, A. L-ornithine L-aspartate restores mitochondrial function and modulates intracellular calcium homeostasis in Parkinson’s disease models. Cells 2022, 11, 2909. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Fröhlich, D.; Achanta, L.B.; Rowlands, B.D.; Housley, G.D.; Klugmann, M.; Rae, C.D. L-aspartate, L-ornithine and L-ornithine-L-aspartate (LOLA) and their impact on brain energy metabolism. Neurochem. Res. 2020, 45, 1438–1450. [Google Scholar] [CrossRef] [PubMed]
- Tate, W.; Walker, M.; Sweetman, E.; Helliwell, A.; Peppercorn, K.; Edgar, C.; Blair, A.; Chatterjee, A. Molecular Mechanisms of Neuroinflammation in ME/CFS and Long COVID to Sustain Disease and Promote Relapses. Front. Neurol. 2022, 13, 877772. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J.; Smeeton, N.J.; Watt, P.W. Ammonia metabolism, the brain and fatigue; revisiting the link. Prog. Neurobiol. 2010, 91, 200–219. [Google Scholar] [CrossRef]
- Glass, K.A.; Germain, A.; Huang, Y.V.; Hanson, M.R. Urine Metabolomics Exposes Anomalous Recovery after Maximal Exertion in Female ME/CFS Patients. Int. J. Mol. Sci. 2023, 24, 3685. [Google Scholar] [CrossRef]
- Germain, A.; Giloteaux, L.; Moore, G.E.; Levine, S.M.; Chia, J.K.; Keller, B.A.; Stevens, J.; Franconi, C.J.; Mao, X.; Shungu, D.C.; et al. Plasma metabolomics reveals disrupted response and recovery following maximal exercise in myalgic encephalomyelitis/chronic fatigue syndrome. JCI Insight 2022, 7, e157621. [Google Scholar] [CrossRef]
- Bobermin, L.D.; Quincozes-Santos, A. COVID-19 and hyperammonemia: Potential interplay between liver and brain dysfunctions. Brain Behav. Immun.-Health 2021, 14, 100257. [Google Scholar] [CrossRef]
- Lopez-Hernandez, Y.; Monarrez-Espino, J.; Lopez, D.A.G.; Zheng, J.; Borrego, J.C.; Torres-Calzada, C.; Elizalde-Diaz, J.P.; Mandal, R.; Berjanskii, M.; Martinez-Martinez, E.; et al. The plasma metabolome of long COVID patients two years after infection. Sci. Rep. 2023, 13, 12420. [Google Scholar] [CrossRef]
- Ali, R.; Nagalli, S. Hyperammonemia. In StatPearls [Internet]; StatPearls Publishing: Tampa, FL, USA, 2023. [Google Scholar]
- Bosoi, C.R.; Rose, C.F. Identifying the direct effects of ammonia on the brain. Metab. Brain Dis. 2009, 24, 95–102. [Google Scholar] [CrossRef]
- Hernandez-Rabaza, V.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Malaguarnera, M.; Agusti, A.; Llansola, M.; Felipo, V. Hyperammonemia induces glial activation, neuroinflammation and alters neurotransmitter receptors in hippocampus, impairing spatial learning: Reversal by sulforaphane. J. Neuroinflamm. 2016, 13, 41. [Google Scholar] [CrossRef]
- Kircheis, G.; Lüth, S. Pharmacokinetic and pharmacodynamic properties of L-ornithine L-aspartate (LOLA) in hepatic encephalopathy. Drugs 2019, 79 (Suppl. 1), 23–29. [Google Scholar] [CrossRef] [PubMed]
- Kars, M.; Yang, L.; Gregor, M.F.; Mohammed, B.S.; Pietka, T.A.; Finck, B.N.; Patterson, B.W.; Horton, J.D.; Mittendorfer, B.; Hotamisligil, G.S.; et al. Tauroursodeoxycholic Acid may improve liver and muscle but not adipose tissue insulin sensitivity in obese men and women. Diabetes 2010, 59, 1899–1905. [Google Scholar] [CrossRef]
- Jain, M.; Nilsson, R.; Sharma, S.; Madhusudhan, N.; Kitami, T.; Souza, A.L.; Kafri, R.; Kirschner, M.W.; Clish, C.B.; Mootha, V.K. Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation. Science 2012, 336, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Brunk, E.; Sahoo, S.; Zielinski, D.C.; Altunkaya, A.; Dräger, A.; Mih, N.; Gatto, F.; Nilsson, A.; Preciat Gonzalez, G.A.; Aurich, M.K. Recon3D enables a three-dimensional view of gene variation in human metabolism. Nat. Biotechnol. 2018, 36, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-H.; Dai, S.; Han, F.; Li, W.; Huang, J.; Xiao, W. FastMM: An efficient toolbox for personalized constraint-based metabolic modeling. BMC Bioinform. 2020, 21, 67. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Hakimi, A.A.; Reznik, E.; Lee, C.-H.; Creighton, C.J.; Brannon, A.R.; Luna, A.; Aksoy, B.A.; Liu, E.M.; Shen, R.; Lee, W.; et al. An Integrated Metabolic Atlas of Clear Cell Renal Cell Carcinoma. Cancer Cell 2016, 29, 104–116. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.-H.; Han, F.-F.; Kalafatis, E.; Kong, Q.-P.; Xiao, W. Systems Modeling Reveals Shared Metabolic Dysregulation and Potential Treatments in ME/CFS and Long COVID. Int. J. Mol. Sci. 2025, 26, 6082. https://doi.org/10.3390/ijms26136082
Li G-H, Han F-F, Kalafatis E, Kong Q-P, Xiao W. Systems Modeling Reveals Shared Metabolic Dysregulation and Potential Treatments in ME/CFS and Long COVID. International Journal of Molecular Sciences. 2025; 26(13):6082. https://doi.org/10.3390/ijms26136082
Chicago/Turabian StyleLi, Gong-Hua, Fei-Fei Han, Efthymios Kalafatis, Qing-Peng Kong, and Wenzhong Xiao. 2025. "Systems Modeling Reveals Shared Metabolic Dysregulation and Potential Treatments in ME/CFS and Long COVID" International Journal of Molecular Sciences 26, no. 13: 6082. https://doi.org/10.3390/ijms26136082
APA StyleLi, G.-H., Han, F.-F., Kalafatis, E., Kong, Q.-P., & Xiao, W. (2025). Systems Modeling Reveals Shared Metabolic Dysregulation and Potential Treatments in ME/CFS and Long COVID. International Journal of Molecular Sciences, 26(13), 6082. https://doi.org/10.3390/ijms26136082