
Non-Animal Hyaluronic Acid and Probiotics Enhance Skin Health via the Gut–Skin Axis: An In Vitro Study on Bioavailability and Cellular Impact



Abstract
:1. Introduction
2. Results
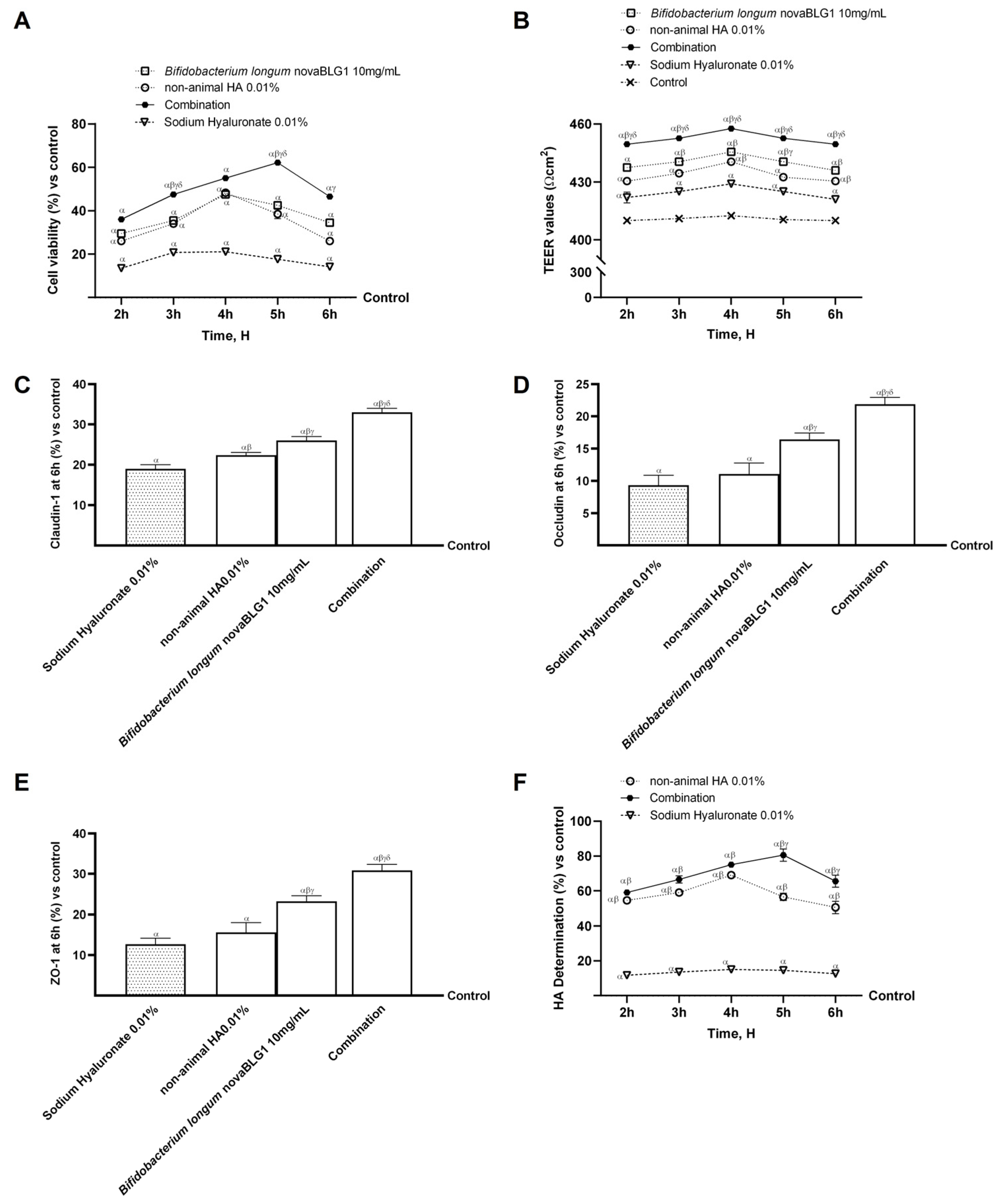
2.1. Dose–Response and Time-Course Study of Non-Animal HA and Bifidobacterium longum novaBLG1 in Caco-2 Cells
2.2. Absorption Evaluation of Non-Animal HA Alone and in Combination with Bifidobacterium longum in Intestinal In Vitro Model
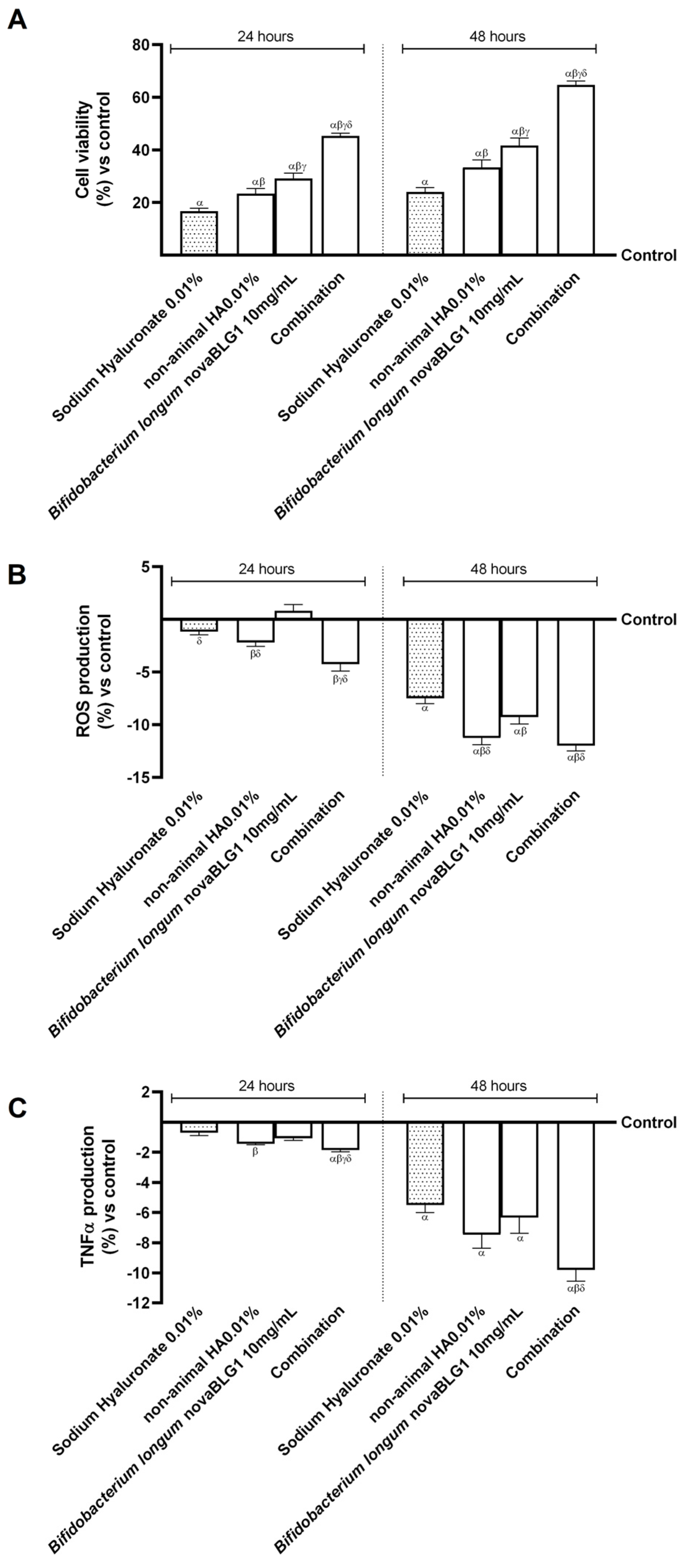
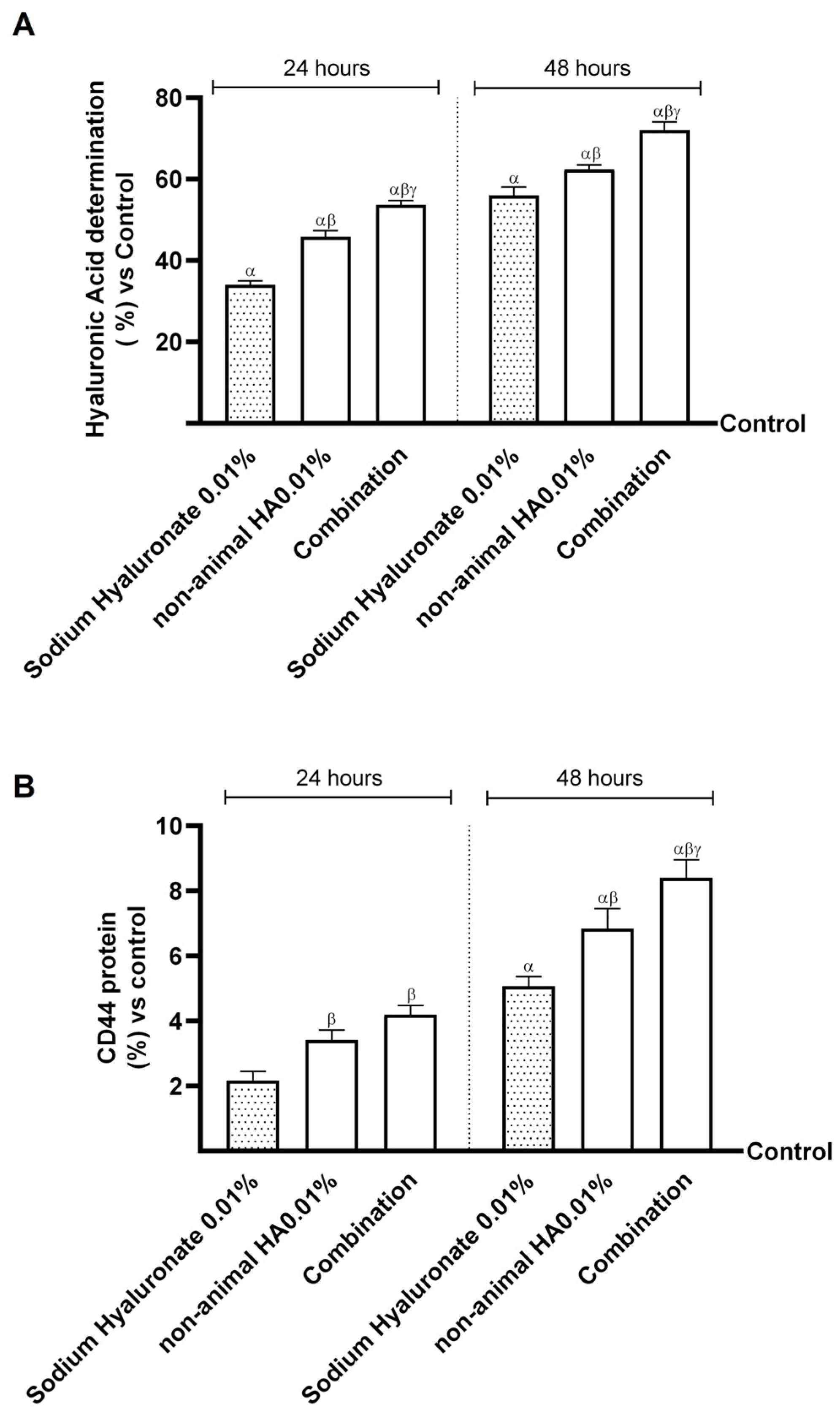
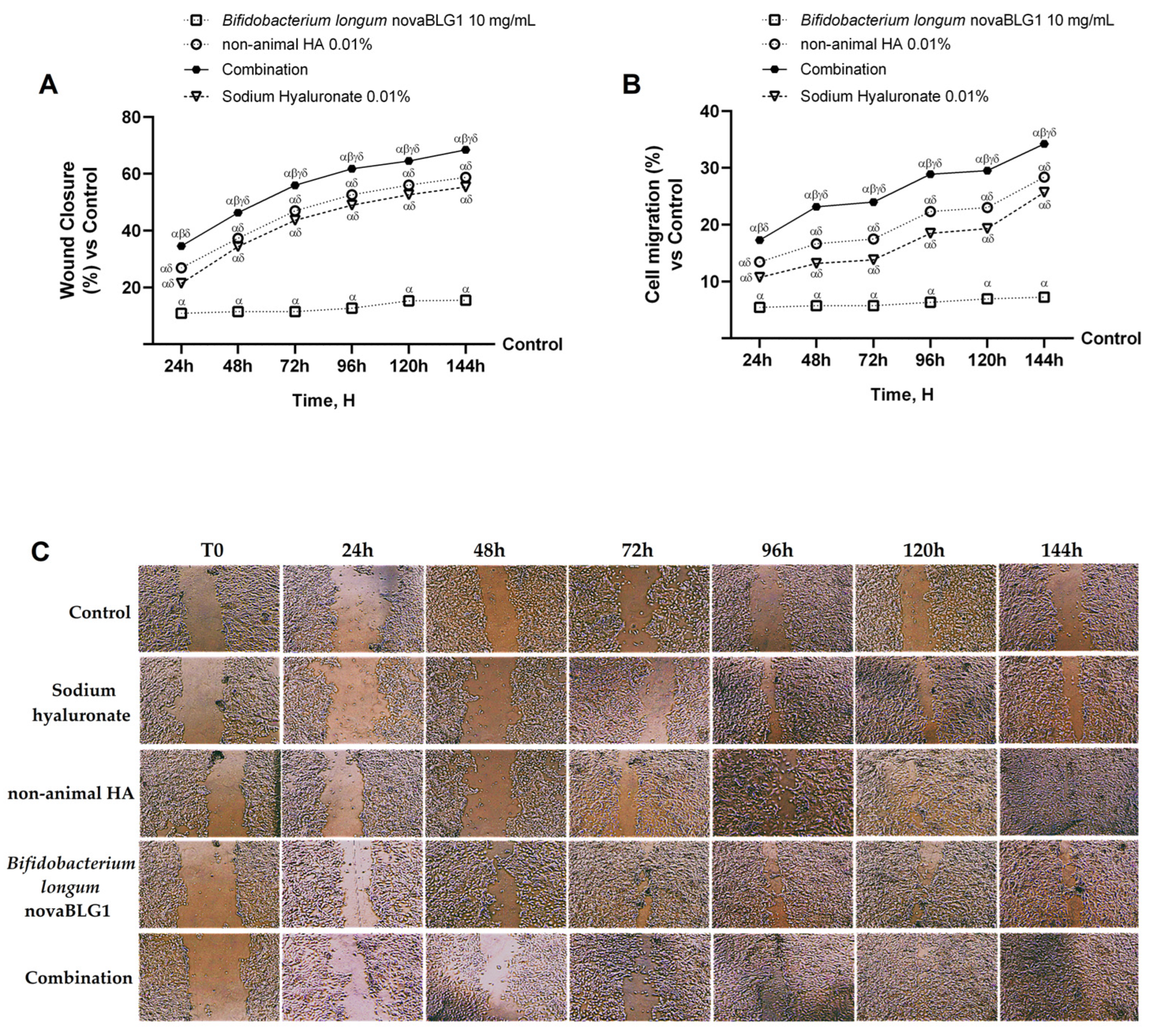
2.3. Effects of Non-Animal HA and Bifidobacterium longum on Gut–Skin Axis
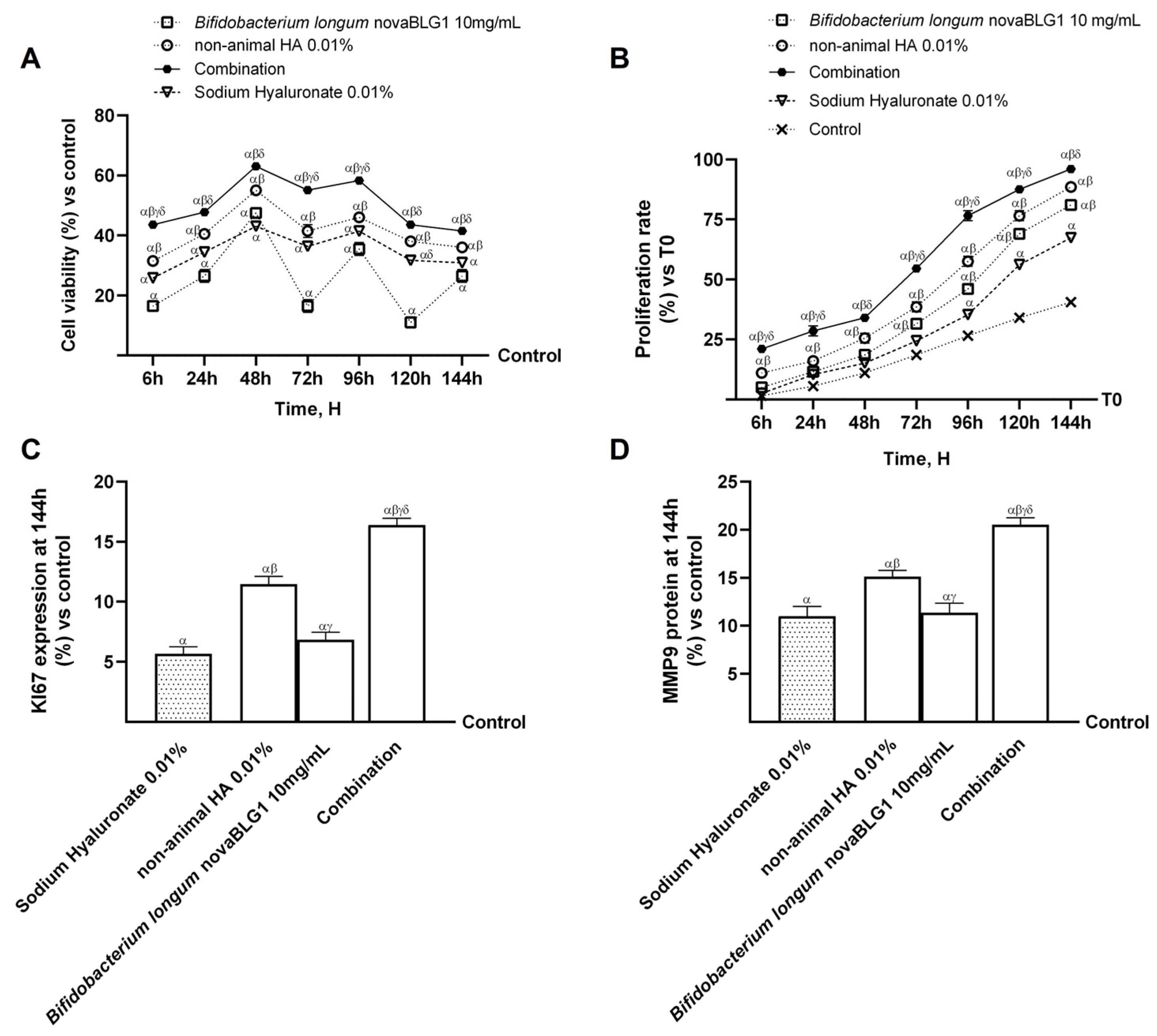
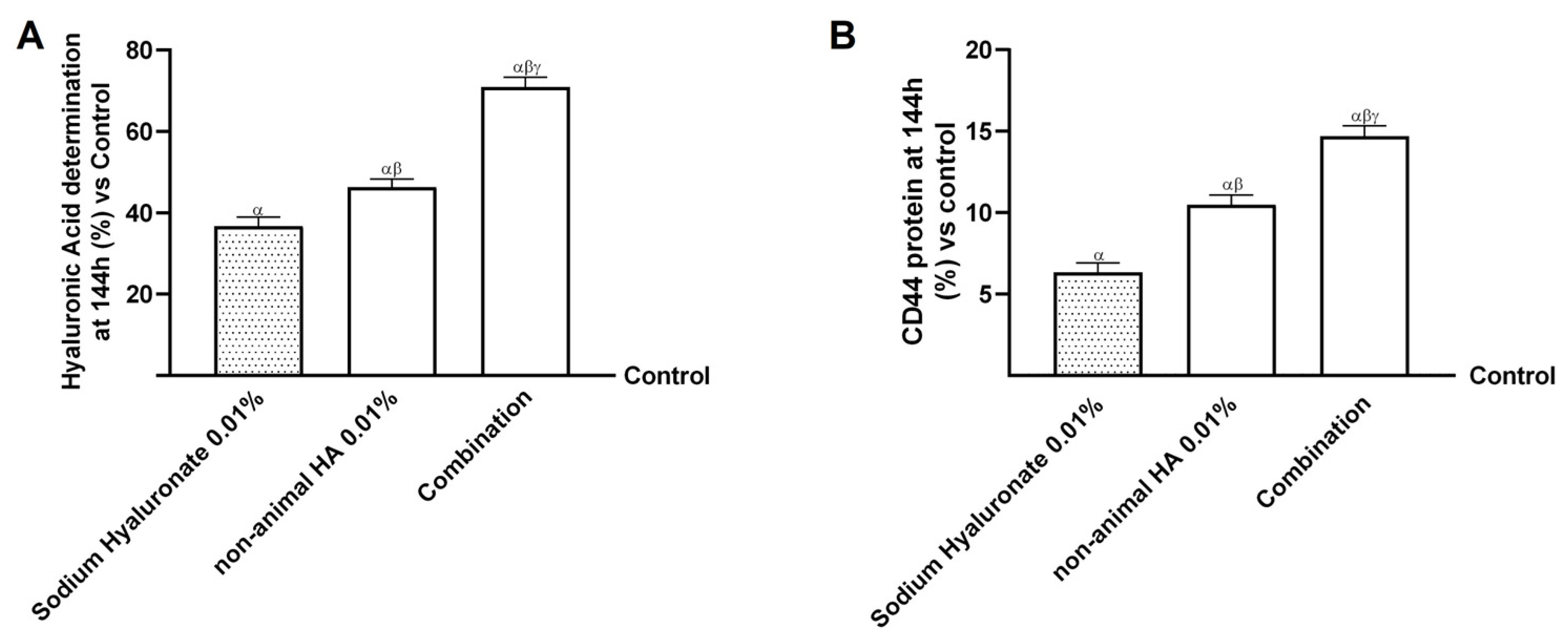
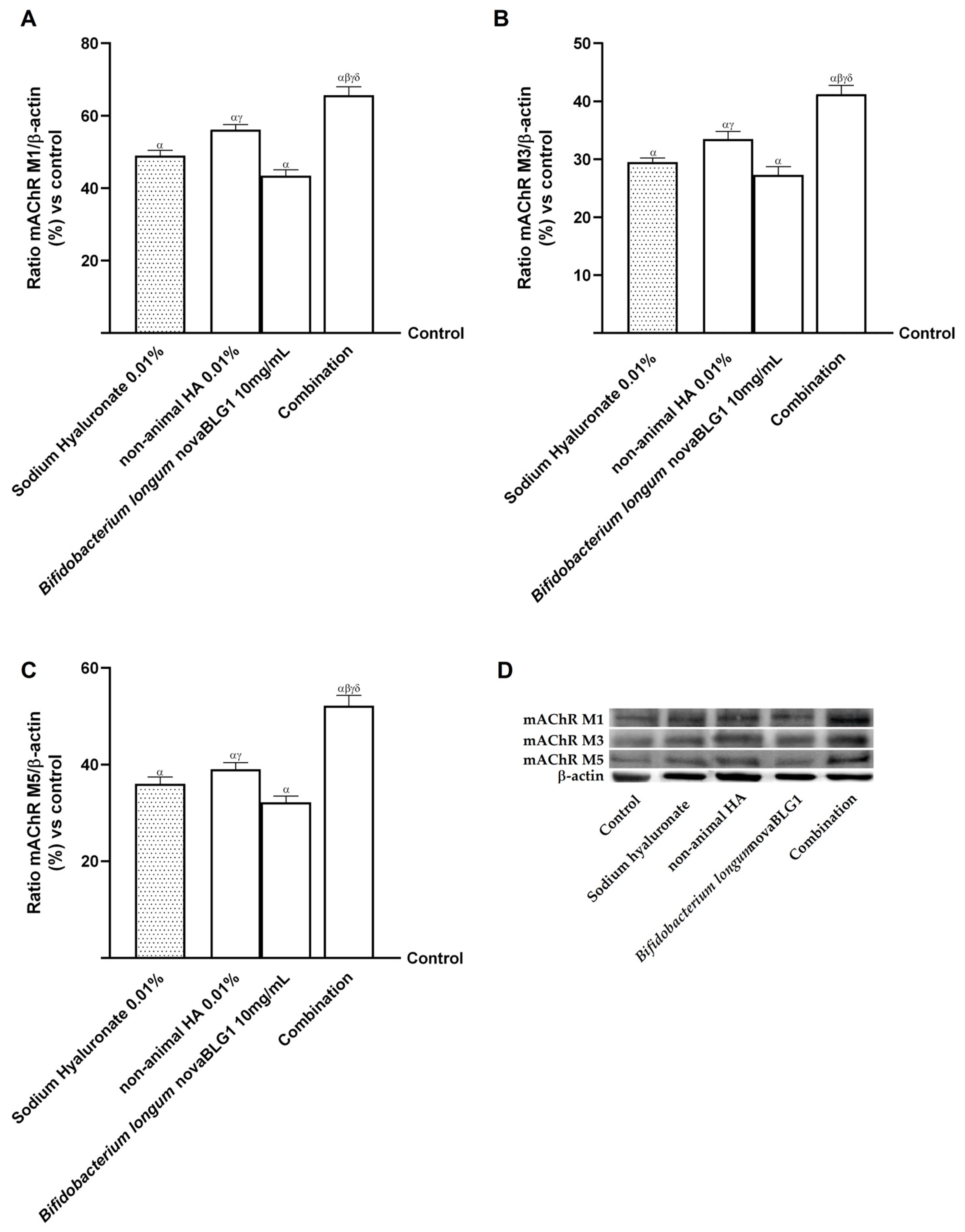
2.4. Effects of Non-Animal HA and Bifidobacterium longum novaBLG1 on Gut–Skin Axis After 144 h of Treatment
3. Discussion
4. Materials and Methods
4.1. Agent Preparation
4.2. Cell Culture
4.3. Experimental Protocol
4.4. In Vitro Intestinal Barrier Model
4.5. In Vitro Toxicology Assay Kit
4.6. Human Claudin-1 ELISA Kit
4.7. Human Occludin ELISA Kit
4.8. Human Tight Junction Protein 1 ELISA Kit
4.9. Gut–Skin Axis
4.10. ROS Production
4.11. TNFα Production ELISA Kit
4.12. Hyaluronic Acid ELISA Kit
4.13. CD44 ELISA Kit
4.14. Crystal Violet
4.15. MMP9 ELISA Kit
4.16. Ki67 ELISA Kit
4.17. Cell Migration Assay
4.18. Wound Healing Test
4.19. Western Blot Analysis
4.20. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Adv DMEM | Advanced Dulbecco’s Modified Eagle’s Medium |
| Adv DMEM-F12 | Advanced Dulbecco’s Modified Eagle’s Medium/Nutrient F-12 Ham’s |
| APQ3 | aquaporin-3 |
| CD44 | cluster of differentiation 44 |
| cfu | colony formant unit |
| ECM | extracellular matrix |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| EMA | European Medicines Agency |
| FBS | foetal bovine serum |
| FDA | Food and Drug Administration |
| HA | hyaluronic acid |
| HMW-HA | high-molecular-weight hyaluronic acid |
| KBM | keratinocyte basal medium |
| KGM2 | keratinocyte growth supplements |
| Ki67 | Kiel 67 protein |
| LMW-HA | low-molecular-weight HA |
| mAChR M1 | muscarinic acetylcholine receptor type 1 |
| mAChR M3 | muscarinic acetylcholine receptor type 3 |
| mAChR M5 | muscarinic acetylcholine receptor type 5 |
| MMP9 | matrix metalloproteinase-9 |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| Papp | apparent permeability coefficient |
| PBS | phosphate-buffered saline |
| PMSF | phenylmethanesulfonylfluoride |
| PVDF | polyvinylidene difluoride |
| ROS | reactive oxygen species |
| TEER | trans-epithelial electrical resistance |
| TJ | tight junction |
| TLR4 | Toll-like receptor 4 |
| TNFα | tumour necrosis factor alpha |
| ZO-1 | Zonula occludens-1 |
References
- Díaz-García, D.; Filipová, A.; Garza-Veloz, I.; Martinez-Fierro, M.L. A Beginner’s Introduction to Skin Stem Cells and Wound Healing. Int. J. Mol. Sci. 2021, 22, 11030. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Luo, Y.; Wang, T.; Wan, C.; Pan, L.; Pan, S.; He, K.; Neo, A.; Chen, X. Artificial Skin Perception. Adv. Mater. 2021, 33, 2003014. [Google Scholar] [CrossRef] [PubMed]
- Uberti, F.; Morsanuto, V.; Ghirlanda, S.; Ruga, S.; Clemente, N.; Boieri, C.; Boldorini, R.; Molinari, C. Highly Diluted Acetylcholine Promotes Wound Repair in an In Vivo Model. Adv. Wound Care 2018, 7, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, A.K.; Spano, F.; Derler, S.; Adlhart, C.; Spencer, N.D.; Rossi, R.M. The relationship between skin function, barrier properties, and body-dependent factors. Skin. Res. Technol. 2018, 24, 165–174. [Google Scholar] [CrossRef]
- Chng, W.Q.; Samuel, M.; Naidoo, K.; Jaffar, H.; Khor, I.W.; Yiong Huak, C.; Watson, R.E.B.; De Mozzi, P.; Tam, W.W.S.; Camargo, C.P.; et al. Topical treatments and skin-resurfacing techniques for skin ageing. Cochrane Database Syst. Rev. 2021, 2021, CD014391. [Google Scholar] [CrossRef]
- Wlaschek, M.; Tantcheva-Poór, I.; Naderi, L.; Ma, W.; Schneider, L.A.; Razi-Wolf, Z.; Schüller, J.; Scharffetter-Kochanek, K. Solar UV irradiation and dermal photoaging. J. Photochem. Photobiol. B Biol. 2001, 63, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.Y.; Huang, Y.L.; Pu, C.M.; Kang, Y.N.; Hoang, K.D.; Chen, K.H.; Chen, C. Effects of Oral Collagen for Skin Anti-Aging: A Systematic Review and Meta-Analysis. Nutrients 2023, 15, 2080. [Google Scholar] [CrossRef]
- Kawano, Y.; Patrulea, V.; Sublet, E.; Borchard, G.; Iyoda, T.; Kageyama, R.; Morita, A.; Seino, S.; Yoshida, H.; Jordan, O.; et al. Wound healing promotion by hyaluronic acid: Effect of molecular weight on gene expression and in vivo wound closure. Pharmaceuticals 2021, 14, 301. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, N.; Kon, R.; Kaneko, M.; Mizukami, N.; Kusunoki, Y.; Sugiyama, K. Relationship between aging-related skin dryness and aquaporins. Int. J. Mol. Sci. 2017, 18, 1559. [Google Scholar] [CrossRef]
- Stern, R.; Maibach, H.I. Hyaluronan in skin: Aspects of aging and its pharmacologic modulation. Clin. Dermatol. 2008, 26, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.R.; Wang, R.P.; Zhang, L.; Fan, Y.; Luan, J.; Liu, Z.; Yuan, C. Oral administration of hyaluronic acid to improve skin conditions via a randomized double-blind clinical test. Skin Res. Technol. 2023, 29, e13531. [Google Scholar] [CrossRef] [PubMed]
- Asari, A.; Kanemitsu, T.; Kurihara, H. Oral administration of high molecular weight hyaluronan (900 kDa) controls immune system via toll-like receptor 4 in the intestinal epithelium. J. Biol. Chem. 2010, 285, 24751–24758. [Google Scholar] [CrossRef]
- Chistyakov, D.V.; Astakhova, A.A.; Azbukina, N.V.; Goriainov, S.V.; Chistyakov, V.V.; Sergeeva, M.G. High and low molecular weight hyaluronic acid differentially influences oxylipins synthesis in course of neuroinflammation. Int. J. Mol. Sci. 2019, 20, 3894. [Google Scholar] [CrossRef] [PubMed]
- Kawada, C.; Yoshida, T.; Yoshida, H.; Matsuoka, R.; Sakamoto, W.; Odanaka, W.; Sato, T.; Yamasaki, T.; Kanemitsu, T.; Masuda, Y.; et al. Ingested hyaluronan moisturizes dry skin. Nutr. J. 2014, 13, 70. [Google Scholar] [CrossRef] [PubMed]
- Juncan, A.M.; Moisă, D.G.; Santini, A.; Morgovan, C.; Rus, L.L.; Vonica-Țincu, A.L.; Loghin, F. Advantages of Hyaluronic Acid and Its Combination with Other Bioactive Ingredients in Cosmeceuticals. Molecules 2021, 26, 4429. [Google Scholar] [CrossRef] [PubMed]
- Goldman, L.; Schafer, A.I. Goldman’s Cecil Medicine E-Book; Elsevier Health Sciences: Philadelphia, PA, USA, 2011. [Google Scholar]
- Wu, X.R.; Mukewar, S.; Kiran, R.P.; Remzi, F.H.; Hammel, J.; Shen, B. Risk factors for peristomal pyoderma gangrenosum complicating inflammatory bowel disease. J. Crohns Colitis 2013, 7, e171–e177. [Google Scholar] [CrossRef]
- Wu, J.J.; Nguyen, T.U.; Poon KY, T.; Herrinton, L.J. The association of psoriasis with autoimmune diseases. J. Am. Acad. Dermatol. 2012, 67, 924–930. [Google Scholar] [CrossRef]
- Parodi, A.; Paolino, S.; Greco, A.; Drago, F.; Mansi, C.; Rebora, A.; Savarino, V. Small intestinal bacterial overgrowth in rosacea: Clinical effectiveness of its eradication. Clin. Gastroenterol. Hepatol. 2008, 6, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Li, R.; Yan, N.; Chen, G.; Qian, W.; Jiang, H.L.; Bi, Z.G. Astragaloside IV controls collagen reduction in photoaging skin by improving transforming growth factor-β/Smad signaling suppression and inhibiting matrix metalloproteinase-1. Mol. Med. Rep. 2015, 11, 3344–3348. [Google Scholar] [CrossRef]
- Xia, W.; Quan, T.; Hammerberg, C.; Voorhees, J.J.; Fisher, G.J. A mouse model of skin aging: Fragmentation of dermal collagen fibrils and reduced fibroblast spreading due to expression of human matrix metalloproteinase-1. J. Dermatol. Sci. 2015, 78, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Lee, C.Y.; Chung, D.K. Probiotic lactic acid bacteria and skin health. Crit. Rev. Food Sci. Nutr. 2016, 56, 2331–2337. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Lee, D.E.; Park, S.D.; Kim, Y.T.; Kim, Y.J.; Jeong, J.W.; Lee, J.H. Oral administration of Lactobacillus plantarum HY7714 protects hairless mouse against ultraviolet B-induced photoaging. J. Microbiol. Biotechnol. 2014, 24, 1583–1591. [Google Scholar] [CrossRef]
- Lee, D.E.; Huh, C.S.; Ra, J.; Choi, I.D.; Jeong, J.W.; Kim, S.H.; Ahn, Y.T. Clinical evidence of effects of Lactobacillus plantarum HY7714 on skin aging: A randomized, double blind, placebo-controlled study. J. Microbiol. Biotechnol. 2015, 25, 2160–2168. [Google Scholar] [CrossRef]
- Szántó, M.; Dózsa, A.; Antal, D.; Szabó, K.; Kemény, L.; Bai, P. Targeting the gut-skin axis-Probiotics as new tools for skin disorder management? Exp. Dermatol. 2019, 28, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Yakoob, R.; Pradeep, B.V. Bifidobacterium sp. as Probiotic Agent—Roles and Applications. J. Pure Appl. Microbiol. 2019, 13, 1407–1417. [Google Scholar] [CrossRef]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Båtsman, A.; Salminen, S. Probiotics for the skin: A new area of potential application? Lett. Appl. Microbiol. 2003, 36, 327–331. [Google Scholar] [CrossRef]
- Hong, Y.K.; An, S.; Lee, Y.H.; Yang, S.A.; Yoon, Y.K.; Lee, J.; Lee, G.; Chung, M.J.; Bae, S. Potential anti-ageing effects of probiotic-derived conditioned media on human skin cells. Acta Pharm. 2022, 72, 359–374. [Google Scholar] [CrossRef]
- Gao, T.; Li, Y.; Wang, X.; Tao, R.; Ren, F. Bifidobacterium longum 68S mediated gut-skin axis homeostasis improved skin barrier damage in aging mice. Phytomedicine 2023, 120, 155051. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jackson, C.J. Autocrine actions of matrix metalloproteinase (MMP)-2 counter the effects of MMP-9 to promote survival and prevent terminal differentiation of cultured human keratinocytes. J. Investig. Dermatol. 2008, 128, 2676–2685. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.K.; Wilson, C.H.; Harding, K.G.; Finlay, A.Y.; Bowden, P.E. Numerous keratinocyte subtypes involved in wound re-epithelialization. J. Investig. Dermatol. 2006, 126, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Coolen, N.A.; Verkerk, M.; Reijnen, L.; Vlig, M.; van den Bogaerdt, A.J.; Breetveld, M.; Gibbs, S.; Middelkoop, E.; Ulrich, M.M. Culture of keratinocytes for transplantation without the need of feeder layer cells. Cell Transplant. 2007, 16. [Google Scholar] [CrossRef] [PubMed]
- Ockenga, W.; Kühne, S.; Bocksberger, S.; Banning, A.; Tikkanen, R. Epidermal growth factor receptor transactivation is required for mitogen-activated protein kinase activation by muscarinic acetylcholine receptors in HaCaT keratinocytes. Int. J. Mol. Sci. 2014, 15, 21433–21454. [Google Scholar] [CrossRef]
- Anaba, E. The Role of Nutraceuticals In Skin Anti-aging. Niger. J. Dermatol. 2020, 10, 13–19. [Google Scholar]
- Shang, L.; Li, M.; Xu, A.; Zhuo, F. Recent applications and molecular mechanisms of hyaluronic acid in skin aging and wound healing. Med. Nov. Technol. Devices 2024, 23, 100320. [Google Scholar] [CrossRef]
- Shin, S.H.; Lee, Y.H.; Rho, N.K.; Park, K.Y. Skin aging from mechanisms to interventions: Focusing on dermal aging. Front. Physiol. 2023, 14, 1195272. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, E.; Dizaj, S.M.; Eftekhari, A.; Dalir, E.; Vahedi, P.; Hasanzadeh, A.; Samiei, M. The potential applications of hyaluronic acid hydrogels in biomedicine. Drug Res. 2020, 70, 6–11. [Google Scholar] [CrossRef]
- Bukhari, S.N.A.; Roswandi, N.L.; Waqas, M.; Habib, H.; Hussain, F.; Khan, S.; Sohail, M.; Ramli, N.A.; Thu, H.E.; Hussain, Z. Hyaluronic acid, a promising skin rejuvenating biomedicine: A review of recent updates and pre-clinical and clinical investigations on cosmetic and nutricosmetic effects. Int. J. Biol. Macromol. 2018, 120 Pt. B, 1682–1695. [Google Scholar] [CrossRef]
- Cortes, H.; Caballero-Florán, I.H.; Mendoza-Muñoz, N.; Córdova-Villanueva, E.N.; Escutia-Guadarrama, L.; Figueroa-González, G.; Reyes-Hernández, O.D.; González-Del Carmen, M.; Varela-Cardoso, M.; Magaña, J.J.; et al. Hyaluronic acid in wound dressings. Cell. Mol. Biol. 2020, 66, 191–198. [Google Scholar] [CrossRef]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Deng, F.; Li, Y.; Wei, X.; Zhao, J. Editorial: The gut-skin axis: Interaction of gut microbiome and skin diseases. Front. Microbiol. 2024, 15, 1427770. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, B.; Tang, X.; Mao, B.; Zhang, Q.; Zhang, T.; Chen, W. Absorption, metabolism, and functions of hyaluronic acid and its therapeutic prospects in combination with microorganisms: A review. Carbohydr. Polym. 2023, 299, 120153. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight Junction in the Intestinal Epithelium: Its Association with Diseases and Regulation by Phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef]
- Nicholas-Haizelden, K.; Murphy, B.; Hoptroff, M.; Horsburgh, M.J. Bioprospecting the Skin Microbiome: Advances in Therapeutics and Personal Care Products. Microorganisms 2023, 11, 1899. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.Y.; Lee, Y.J.; Kim, C.M.; Lee, Y.-M. Revolutionizing Cosmetic Ingredients: Harnessing the Power of Antioxidants, Probiotics, Plant Extracts, and Peptides in Personal and Skin Care Products. Cosmetics 2024, 11, 157. [Google Scholar] [CrossRef]
- Gruber, J.V.; Holtz, R.; Riemer, J. Hyaluronic acid (HA) stimulates the in vitro expression of CD44 proteins but not HAS1 proteins in normal human epidermal keratinocytes (NHEKs) and is HA molecular weight dependent. J. Cosmet. Dermatol. 2022, 21, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.F.; Su, Z.R.; Hsieh, Y.H.; Wang, M.F.; Oe, M.; Matsuoka, R.; Masuda, Y. Oral Hyaluronan Relieves Wrinkles and Improves Dry Skin: A 12-Week Double-Blinded, Placebo-Controlled Study. Nutrients 2021, 13, 2220. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Sakamoto, W.; Odanaka, W.; Yoshida, K.; Urushibata, O. Clinical effects of hyaluronic acid diet for dry and rough skin. Aesthe. Derma. 2002, 12, 109–120. [Google Scholar]
- Sato, T.; Yoshida, T.; Kanemitsu, T.; Yoshida, K.; Hasegawa, M.; Urushibata, O. Clinical effects of hyaluronic acid diet for moisture content of dry skin. Aesthe. Derma. 2007, 17, 33–39. [Google Scholar]
- Yoshida, T.; Kanemitsu, T.; Narabe, O.; Tobita, M. Improvement of dry skin by a food containing hyaluronic acids derived from microbial fermentation. J. New Rem. Clin. 2009, 58, 143–155. [Google Scholar]
- Kawada, C.; Yoshida, T.; Yoshida, H.; Sakamoto, W.; Odanaka, W.; Sato, T.; Yamasaki, T.; Kanemitsu, T.; Masuda, Y.; Urushibata, O. Ingestion of hyaluronans (molecular weights 800 k and 300k) improves dry skin conditions: A randomized, double blind, controlled study. J. Clin. Biochem. Nutr. 2015, 56, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.R.; Park, J. Ingestion of BioCell Collagen®, a novel hydrolyzed chicken sternal cartilage extract; enhanced blood microcirculation and reduced facial aging signs. Clin. Interv. Aging. 2012, 7, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, Y.; Zhang, L. Tremella polysaccharide: The molecular mechanisms of its drug action. Prog. Mol. Biol. Transl. Sci. 2019, 163, 383–421. [Google Scholar] [CrossRef]
- Vivatis Pharma GMBH. Plant Producing Hyaluronic Acid. PCT/IB2021/055031, 8 June 2020. Available online: https://patents.google.com/patent/WO2021250566A1/en?oq=WO2021250566+(A1)++-++PRO-CESS+FOR+EXTRACTING+A+HYALURONIC+ACID+FROM+A+FUNGUS%2c+A+HYALURONIC+ACID+OF+PLANT+ORIGIN+AND+USE+THEREOF (accessed on 1 June 2022).
- Identification and Selection of a Plant Starting Material of a Plant Chondroitin Sulfate and Hyaluronic Acid, and Transformation of such Plant Starting Material to Obtain Ingredients for Use in Foods, Supplements, Medical Devices or Drugs. Italian Patent 102019000008409, 8 June 2021. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2020245809 (accessed on 18 September 2024).
- Galla, R.; Grisenti, P.; Farghali, M.; Saccuman, L.; Ferraboschi, P.; Uberti, F. Ovotransferrin supplementation improves the iron absorption: An in vitro gastro-intestinal model. Biomedicines 2021, 9, 1543. [Google Scholar] [CrossRef] [PubMed]
- Uberti, F.; Morsanuto, V.; Ruga, S.; Galla, R.; Farghali, M.; Notte, F.; Bozzo, C.; Magnani, C.; Nardone, A.; Molinari, C. Study of magnesium formulations on intestinal cells to influence myometrium cell relaxation. Nutrients 2020, 12, 573. [Google Scholar] [CrossRef] [PubMed]
- Galla, R.; Ruga, S.; Aprile, S.; Ferrari, S.; Brovero, A.; Grosa, G.; Molinari, C.; Uberti, F. New Hyaluronic Acid from Plant Origin to Improve Joint Protection—An In Vitro Study. Int. J. Mol. Sci. 2022, 23, 8114. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.D.; Kang, T.J.; Lee, C.H.; Lee, A.Y.; Noh, M. HaCaT Keratinocytes and Primary Epidermal Keratinocytes Have Different Transcriptional Profiles of Cornified Envelope-Associated Genes to T Helper Cell Cytokines. Biomol. Ther. 2012, 20, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Ruga, S.; Galla, R.; Ferrari, S.; Invernizzi, M.; Uberti, F. Novel Approach to the Treatment of Neuropathic Pain Using a Combination with Palmitoylethanolamide and Equisetum arvense L. in an In Vitro Study. Int. J. Mol. Sci. 2023, 24, 5503. [Google Scholar] [CrossRef]
- Fda.Gov. Available online: https://www.fda.gov/media/117974/download (accessed on 23 May 2024).
- Ema.Eu. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/ich-m9-biopharmaceutics-classification-system-based-biowaivers-step-2b-first-version_en.pdf (accessed on 23 May 2024).
- Guha, S.; Alvarez, S.; Majumder, K. Transport of dietary anti-inflammatory peptide, γ-glutamyl valine (γ-ev), across the intestinal caco-2 monolayer. Nutrients 2021, 13, 1448. [Google Scholar] [CrossRef] [PubMed]
- Molinari, C.; Morsanuto, V.; Ruga, S.; Notte, F.; Farghali, M.; Galla, R.; Uberti, F. The role of BDNF on aging-modulation markers. Brain Sci. 2020, 10, 285. [Google Scholar] [CrossRef]
- Ahn, M.; Cho, W.W.; Lee, H.; Park, W.; Lee, S.H.; Back, J.W.; Gao, Q.; Gao, G.; Cho, D.W.; Kim, B.S. Engineering of Uniform Epidermal Layers via Sacrificial Gelatin Bioink-Assisted 3D Extrusion Bioprinting of Skin. Adv. Healthc. Mater. 2023, 12, 2301015. [Google Scholar] [CrossRef]
- Ieiri, H.; Kameda, N.; Naito, J.; Kawano, T.; Nishida, N.; Takahashi, M.; Katakura, Y. Paramylon extracted from Euglena gracilis EOD-1 augmented the expression of SIRT1. Cytotechnology 2021, 73, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Molinari, C.; Ruga, S.; Farghali, M.; Galla, R.; Bassiouny, A.; Uberti, F. Preventing c2c12 muscular cells damage combining magnesium and potassium with vitamin D3 and curcumin. J. Tradit. Complement. Med. 2021, 11, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Aborehab, N.M.; El Bishbishy, M.H.; Refaiy, A.; Waly, N.E. A putative Chondroprotective role for IL-1β and MPO in herbal treatment of experimental osteoarthritis. BMC Complement. Altern. Med. 2017, 17, 495. [Google Scholar] [CrossRef]
- Roberts, H.M.; Moore, J.P.; Thom, J.M. The effect of aerobic walking and lower body resistance exercise on serum COMP and hyaluronan, in both males and females. Eur. J. Appl. Physiol. 2018, 118, 1095–1105. [Google Scholar] [CrossRef]
- Wöhner, B.; Li, W.; Hey, S.; Drobny, A.; Werny, L.; Becker-Pauly, C.; Lucius, R.; Zunke, F.; Linder, S.; Arnold, P. Proteolysis of CD44 at the cell surface controls a downstream protease network. Front. Mol. Biosci. 2023, 10, 1026810. [Google Scholar] [CrossRef] [PubMed]
- Kramer, N.; Walzl, A.; Unger, C.; Rosner, M.; Krupitza, G.; Hengstschläger, M.; Dolznig, H. In vitro cell migration and invasion assays. Mutat. Res. 2013, 752, 10–24. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Hours (h) | 2 h | 3 h | 4 h | 5 h | 6 h | |
|---|---|---|---|---|---|---|
| Substances | ||||||
| Sodium hyaluronate 0.01% | 0.28 × 10−6 | 0.33 × 10−6 | 0.36 × 10−6 | 0.35 × 10−6 | 0.30 × 10−6 | |
| α Non-animal HA 0.01% | 1.32 × 10−6 | 1.43 × 10−6 | 1.67 × 10−6 | 1.37 × 10−6 | 1.22 × 10−6 | |
| αβ Combination | 1.43 × 10−6 | 1.61 × 10−6 | 1.81 × 10−6 | 1.95 × 10−6 | 1.58 × 10−6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galla, R.; Mulè, S.; Ferrari, S.; Molinari, C.; Uberti, F. Non-Animal Hyaluronic Acid and Probiotics Enhance Skin Health via the Gut–Skin Axis: An In Vitro Study on Bioavailability and Cellular Impact. Int. J. Mol. Sci. 2025, 26, 897. https://doi.org/10.3390/ijms26030897
Galla R, Mulè S, Ferrari S, Molinari C, Uberti F. Non-Animal Hyaluronic Acid and Probiotics Enhance Skin Health via the Gut–Skin Axis: An In Vitro Study on Bioavailability and Cellular Impact. International Journal of Molecular Sciences. 2025; 26(3):897. https://doi.org/10.3390/ijms26030897
Chicago/Turabian StyleGalla, Rebecca, Simone Mulè, Sara Ferrari, Claudio Molinari, and Francesca Uberti. 2025. "Non-Animal Hyaluronic Acid and Probiotics Enhance Skin Health via the Gut–Skin Axis: An In Vitro Study on Bioavailability and Cellular Impact" International Journal of Molecular Sciences 26, no. 3: 897. https://doi.org/10.3390/ijms26030897
APA StyleGalla, R., Mulè, S., Ferrari, S., Molinari, C., & Uberti, F. (2025). Non-Animal Hyaluronic Acid and Probiotics Enhance Skin Health via the Gut–Skin Axis: An In Vitro Study on Bioavailability and Cellular Impact. International Journal of Molecular Sciences, 26(3), 897. https://doi.org/10.3390/ijms26030897