
Calcium Sulfide Nanoclusters Trigger DNA Damage and Induce Cell Cycle Arrest in Non-Small-Cell Lung Adenocarcinoma Cells

 , , , , ,
, , , , ,  , ,
, ,  ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
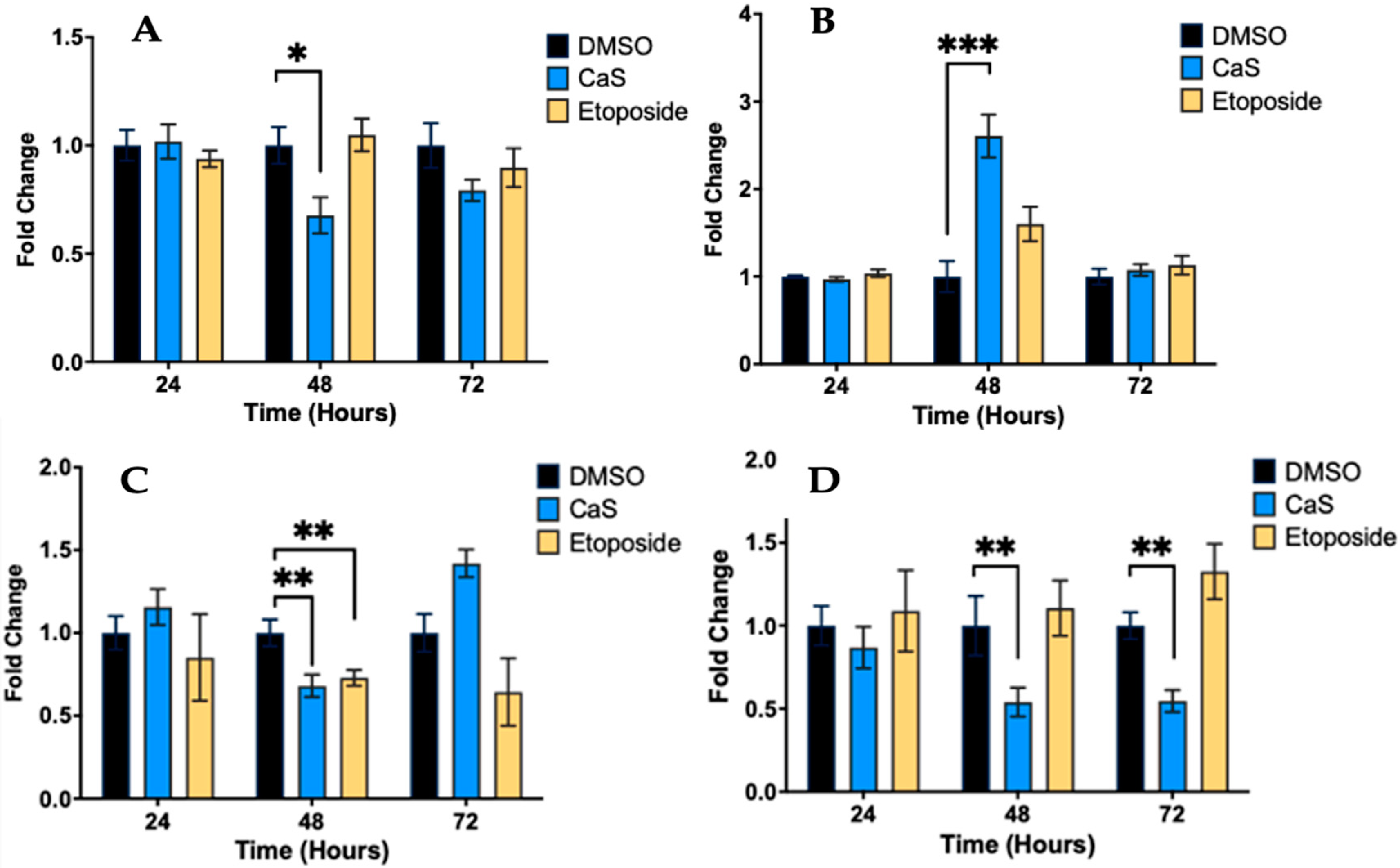
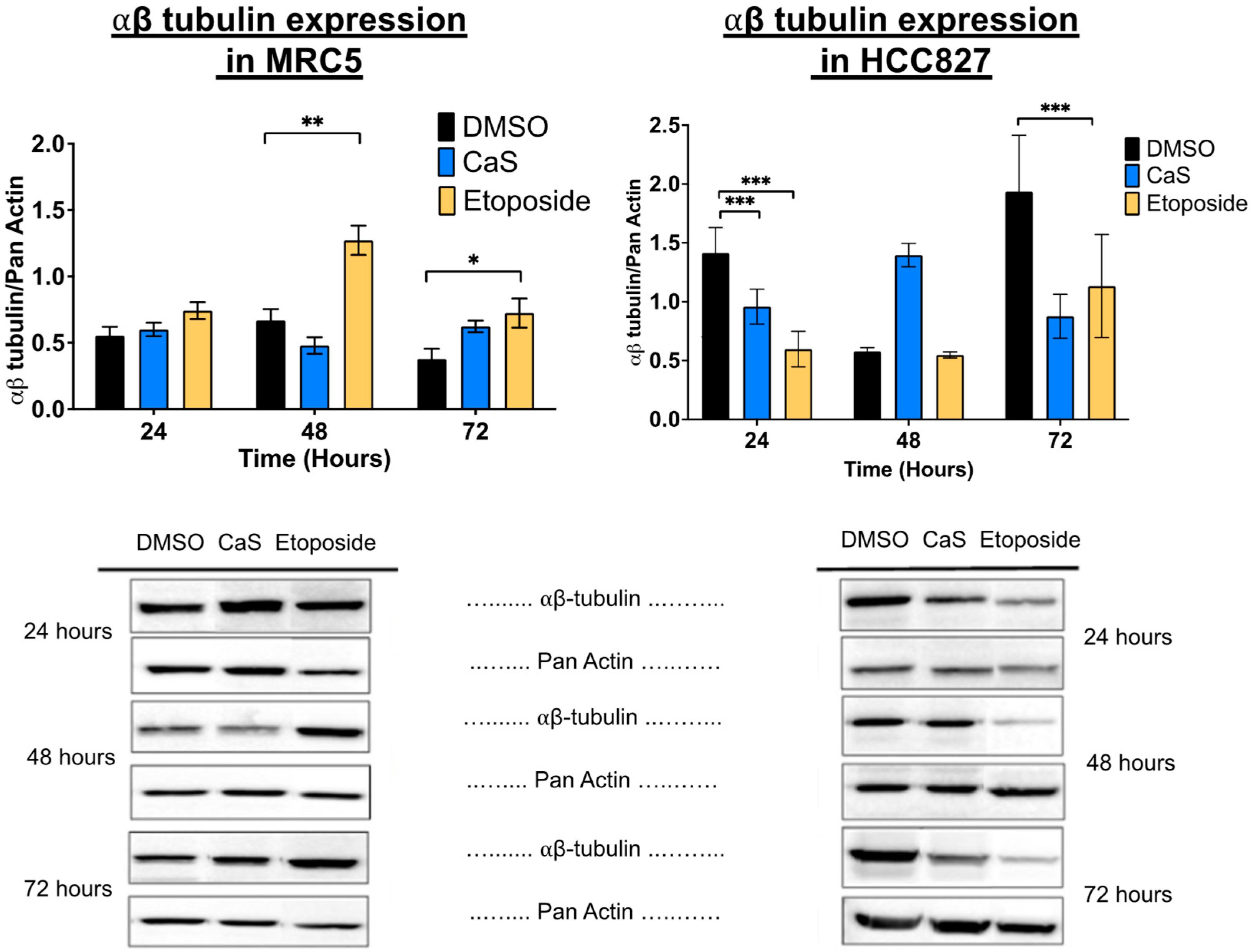
2.1. CaS Nanoclusters Induce Significant DNA Damage in Lung Adenocarcinoma Cells
2.2. CaS Nanoclusters Increase Calcium Concentrations in Lung Adenocarcinoma Cells at Early Time Points, Leading to Cell Death
2.3. CaS Nanoclusters Trigger Oxidative Stress in Lung Adenocarcinoma Cells
2.4. Molecular Mechanisms of CaS Nanoclusters in Lung Cancer Cells Are Less Likely to Be Regulated by Mitochondrial Pathways
2.5. CaS Nanoclusters Are Less Likely to Affect Intrinsic Apoptotic Pathways
2.6. CaS Nanoclusters Selectively Arrest Lung Adenocarcinoma Cells but Not Healthy Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Positive and Negative Controls
4.2. Synthesis of Calcium Sulfide (CaS) Nanoclusters and Cell Treatments
4.3. Annexin V FITC and Cell Cycle Assays
4.4. Whole-Cell Protein Extractions and Immunoblotting
4.5. Extracellular Calcium Free Ion Assay
4.6. Assays Using Flow Cytometry
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA A Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, A.; Akter, Z.; Modareszadeh, P.; Modareszadeh, P.; Berisha, E.; Alemi, P.S.; Chacon Castro, M.D.C.; Deese, A.R.; Zhang, L. Current Landscape of Therapeutic Resistance in Lung Cancer and Promising Strategies to Overcome Resistance. Cancers 2022, 14, 4562. [Google Scholar] [CrossRef] [PubMed]
- Choi, P.J.; Jeong, S.S.; Yoon, S.S. Prognosis of recurrence after complete resection in early-stage non-small cell lung cancer. Korean J. Thorac. Cardiovasc. Surg. 2013, 46, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Trieu, V.; Yao, Z.; Louie, L.; Ci, S.; Yang, A.; Tao, C.; De, T.; Beals, B.; Dykes, D.; et al. Increased antitumor activity, intratumor paclitaxel concentrations, and endothelial cell transport of cremophor-free, albumin-bound paclitaxel, ABI-007, compared with cremophor-based paclitaxel. Clin. Cancer Res. 2006, 12, 1317–1324. [Google Scholar] [CrossRef]
- Dhar, S.; Kolishetti, N.; Lippard, S.J.; Farokhzad, O.C. Targeted delivery of a cisplatin prodrug for safer and more effective prostate cancer therapy in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 1850–1855. [Google Scholar] [CrossRef] [PubMed]
- Hauert, S.; Bhatia, S.N. Mechanisms of cooperation in cancer nanomedicine: Towards systems nanotechnology. Trends Biotechnol. 2014, 32, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Socinski, M.A.; Bondarenko, I.; Karaseva, N.A.; Makhson, A.M.; Vynnychenko, I.; Okamoto, I.; Hon, J.K.; Hirsh, V.; Bhar, P.; Zhang, H.; et al. Weekly nab-paclitaxel in combination with carboplatin versus solvent-based paclitaxel plus carboplatin as first-line therapy in patients with advanced non-small-cell lung cancer: Final results of a phase III trial. J. Clin. Oncol. 2012, 30, 2055–2062. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Wang, X.; Nie, S.; Chen, Z.G.; Shin, D.M. Therapeutic nanoparticles for drug delivery in cancer. Clin. Cancer Res. 2008, 14, 1310–1316. [Google Scholar] [CrossRef]
- Peng, X.; Fang, J.; Lou, C.; Yang, L.; Shan, S.; Wang, Z.; Chen, Y.; Li, H.; Li, X. Engineered nanoparticles for precise targeted drug delivery and enhanced therapeutic efficacy in cancer immunotherapy. Acta. Pharm. Sin. B 2024, 14, 3432–3456. [Google Scholar] [CrossRef]
- Naahidi, S.; Jafari, M.; Edalat, F.; Raymond, K.; Khademhosseini, A.; Chen, P. Biocompatibility of engineered nanoparticles for drug delivery. J. Control. Release 2013, 166, 182–194. [Google Scholar] [CrossRef]
- He, X.W.; Liu, T.; Chen, Y.X.; Cheng, D.J.; Li, X.R.; Xiao, Y.; Feng, Y.L. Calcium carbonate nanoparticle delivering vascular endothelial growth factor-C siRNA effectively inhibits lymphangiogenesis and growth of gastric cancer in vivo. Cancer Gene Ther. 2008, 15, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Maleki Dizaj, S.; Barzegar-Jalali, M.; Zarrintan, M.H.; Adibkia, K.; Lotfipour, F. Calcium carbonate nanoparticles as cancer drug delivery system. Expert Opin. Drug Deliv. 2015, 12, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, M.; Kodela, R.; Olson, K.R.; Kashfi, K. NOSH-aspirin (NBS-1120), a novel nitric oxide- and hydrogen sulfide-releasing hybrid is a potent inhibitor of colon cancer cell growth in vitro and in a xenograft mouse model. Biochem. Biophys. Res. Commun. 2012, 419, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Lencesova, L.; Hudecova, S.; Csaderova, L.; Markova, J.; Soltysova, A.; Pastorek, M.; Sedlak, J.; Wood, M.E.; Whiteman, M.; Ondrias, K.; et al. Sulphide signalling potentiates apoptosis through the up-regulation of IP3 receptor types 1 and 2. Acta Physiol. 2013, 208, 350–361. [Google Scholar] [CrossRef]
- Wu, X.; Kassie, F.; Mersch-Sundermann, V. Induction of apoptosis in tumor cells by naturally occurring sulfur-containing compounds. Mutat. Res. 2005, 589, 81–102. [Google Scholar] [CrossRef]
- Martinez, O.M.R.; Ramos, M.A.N.; Acevedo, A.A.S.; Colon, C.C.C.; Ramos, D.M.; Rivera, C.C.; Rosario, M.E.C. pH-Selective Reactions to Selectively Reduce Cancer Cell Proliferation: Effect of CaS Nanostructures in Human Skin Melanoma and Benign Fibroblasts. BioChem 2023, 3, 15–30. [Google Scholar] [CrossRef]
- Vazquez, D.R.; Munoz Forti, K.; Figueroa Rosado, M.M.; Gutierrez Mirabal, P.I.; Suarez-Martinez, E.; Castro-Rosario, M.E. Effect of CaS Nanostructures in the Proliferation of Human Breast Cancer and Benign Cells In Vitro. Appl. Sci. 2022, 12, 10494. [Google Scholar] [CrossRef]
- Gruosso, T.; Mieulet, V.; Cardon, M.; Bourachot, B.; Kieffer, Y.; Devun, F.; Dubois, T.; Dutreix, M.; Vincent-Salomon, A.; Miller, K.M.; et al. Chronic oxidative stress promotes H2AX protein degradation and enhances chemosensitivity in breast cancer patients. EMBO Mol. Med. 2016, 8, 527–549. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.L.; Orr-Weaver, T.L. Replication fork instability and the consequences of fork collisions from rereplication. Genes Dev. 2016, 30, 2241–2252. [Google Scholar] [CrossRef]
- Rivera, D.; Carrasco, N.; Maldonado, L.; Suarez, E.; Poventud, C.; Marrero, C.; Castro, M. Killing cancer cells with nanotechnology: The effect of CaS nanostructures on the cell replication rate and survival of human mammary adenocarcinoma cell lines (780.9). FASEB J. 2014, 28, 780–789. [Google Scholar] [CrossRef]
- Zheng, Y.; Ji, X.; Ji, K.; Wang, B. Hydrogen sulfide prodrugs—A review. Acta Pharm. Sin. B 2015, 5, 367–377. [Google Scholar] [CrossRef]
- Danhier, F.; Feron, O.; Préat, V. To exploit the tumor microenvironment: Passive and active tumor targeting of nanocarriers for anti-cancer drug delivery. J. Control. Release 2010, 148, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, K.S.; Kuttikrishnan, S.; Ahmad, N.; Habeeba, U.; Mariyam, Z.; Suleman, M.; Bhat, A.A.; Uddin, S. H2AX: A key player in DNA damage response and a promising target for cancer therapy. Biomed. Pharmacother. 2024, 175, 116663. [Google Scholar] [CrossRef]
- Danese, A.; Leo, S.; Rimessi, A.; Wieckowski, M.R.; Fiorica, F.; Giorgi, C.; Pinton, P. Cell death as a result of calcium signaling modulation: A cancer-centric prospective. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119061. [Google Scholar] [CrossRef]
- Thomas, N.L.; Dart, C.; Helassa, N. Editorial: The role of calcium and calcium binding proteins in cell physiology and disease. Front. Physiol. 2023, 14, 1228885. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, C.; Müller, M.; McNeal, A.; Lovy, A.; Jaňa, F.; Bustos, G.; Urra, F.; Smith, N.; Molgó, J.; Diehl, J.A.; et al. Selective Vulnerability of Cancer Cells by Inhibition of Ca(2+) Transfer from Endoplasmic Reticulum to Mitochondria. Cell Rep. 2016, 14, 2313–2324. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, M.; Günther, S.D.; Schwarzer, R.; Albert, M.-C.; Schorn, F.; Werthenbach, J.P.; Schiffmann, L.M.; Stair, N.; Stocks, H.; Seeger, J.M.; et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature 2019, 575, 683–687. [Google Scholar] [CrossRef]
- Yoo, S.H.; Yoon, Y.G.; Lee, J.S.; Song, Y.S.; Oh, J.S.; Park, B.S.; Kwon, T.K.; Park, C.; Choi, Y.H.; Yoo, Y.H. Etoposide induces a mixed type of programmed cell death and overcomes the resistance conferred by Bcl-2 in Hep3B hepatoma cells. Int. J. Oncol. 2012, 41, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.J.; Souès, S.; Gottlieb, T.; Falk, S.J.; Watson, J.V.; Osborne, R.J.; Bleehen, N.M. Etoposide-induced cell cycle delay and arrest-dependent modulation of DNA topoisomerase II in small-cell lung cancer cells. Br. J. Cancer 1994, 70, 914–921. [Google Scholar] [CrossRef]
- Rivera, D.; Castro, M.E. Synthesis of Calcium Sulfide (CaS) Nanoparticles. U.S. Patent 8945494B1, 3 February 2015. [Google Scholar]
- Montecucco, A.; Zanetta, F.; Biamonti, G. Molecular mechanisms of etoposide. Excli. J. 2015, 14, 95–108. [Google Scholar] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueroa Rosado, M.M.; Muñoz Forti, K.; Rodríguez-Rodríguez, P.; Arroyo-Martínez, G.; Rodríguez-Irizarry, V.J.; Ruiz-Rivera, A.; Quinones-Rodriguez, J.I.; Santiago-Cardona, P.G.; Rodriguez Martinez, O.M.; Almodovar, S.; et al. Calcium Sulfide Nanoclusters Trigger DNA Damage and Induce Cell Cycle Arrest in Non-Small-Cell Lung Adenocarcinoma Cells. Int. J. Mol. Sci. 2025, 26, 1665. https://doi.org/10.3390/ijms26041665
Figueroa Rosado MM, Muñoz Forti K, Rodríguez-Rodríguez P, Arroyo-Martínez G, Rodríguez-Irizarry VJ, Ruiz-Rivera A, Quinones-Rodriguez JI, Santiago-Cardona PG, Rodriguez Martinez OM, Almodovar S, et al. Calcium Sulfide Nanoclusters Trigger DNA Damage and Induce Cell Cycle Arrest in Non-Small-Cell Lung Adenocarcinoma Cells. International Journal of Molecular Sciences. 2025; 26(4):1665. https://doi.org/10.3390/ijms26041665
Chicago/Turabian StyleFigueroa Rosado, María M., Kevin Muñoz Forti, Patricia Rodríguez-Rodríguez, Gerardo Arroyo-Martínez, Valerie J. Rodríguez-Irizarry, Abigail Ruiz-Rivera, Jailenne I. Quinones-Rodriguez, Pedro G. Santiago-Cardona, Olga M. Rodriguez Martinez, Sharilyn Almodovar, and et al. 2025. "Calcium Sulfide Nanoclusters Trigger DNA Damage and Induce Cell Cycle Arrest in Non-Small-Cell Lung Adenocarcinoma Cells" International Journal of Molecular Sciences 26, no. 4: 1665. https://doi.org/10.3390/ijms26041665
APA StyleFigueroa Rosado, M. M., Muñoz Forti, K., Rodríguez-Rodríguez, P., Arroyo-Martínez, G., Rodríguez-Irizarry, V. J., Ruiz-Rivera, A., Quinones-Rodriguez, J. I., Santiago-Cardona, P. G., Rodriguez Martinez, O. M., Almodovar, S., Castro, M. E., & Suárez Martínez, E. (2025). Calcium Sulfide Nanoclusters Trigger DNA Damage and Induce Cell Cycle Arrest in Non-Small-Cell Lung Adenocarcinoma Cells. International Journal of Molecular Sciences, 26(4), 1665. https://doi.org/10.3390/ijms26041665