
Emerging Role of Hypoxia-Inducible Factors (HIFs) in Modulating Autophagy: Perspectives on Cancer Therapy

Abstract
1. Introduction
2. Important Function of HIFs in the Progression of Cancer Cells
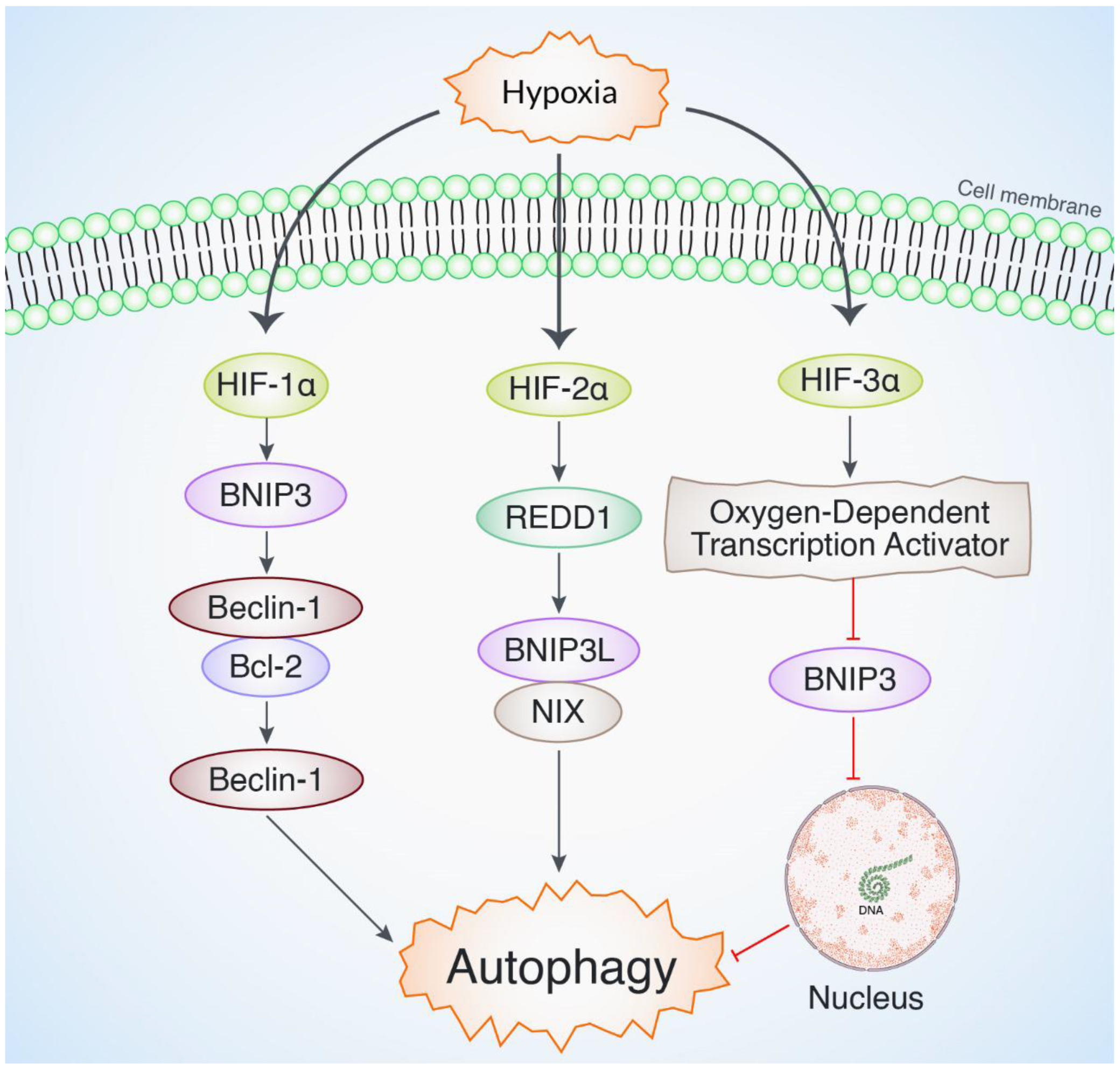
3. Role of Hypoxia-Inducible Factors in Modulating Autophagy and Maintaining Cellular Homeostasis
4. Recent Drug Targets for HIF-Mediated Autophagy Regulation in Cancer
4.1. Natural Products for HIF-Mediated Autophagy Modulation in Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Compound | Tumor Type | Experimental System | Effect on HIFs | Effect on Autophagy | References |
|---|---|---|---|---|---|
| Resveratrol | Breast cancer | In vitro (MCF-7 cells) | Downregulates HIF-1α expression | Activates autophagy via AMPK–mTOR pathway | [58,73] |
| Curcumin | Glioblastoma | In vitro (U87MG cells) | Inhibits HIF-1α stabilization | Induces autophagy by inhibiting PI3K/Akt/mTOR | [59,74] |
| Epigallocatechin gallate | Lung cancer | In vivo (mouse model) | Reduces HIF-1α activity | Promotes autophagy through AMPK activation | [60,75] |
| Berberine | Colorectal cancer | In vitro (HCT116 cells) | Suppresses HIF-1α signaling | Induces autophagy through ROS-mediated pathway | [61,76] |
| Quercetin | Pancreatic cancer | In vivo and in vitro | Downregulates HIF-1α protein levels | Activates autophagy through inhibition of Akt/mTOR | [62,77] |
| Genistein | Prostate cancer | In vitro (PC-3 cells) | Decreases HIF-1α expression | Promotes autophagy via JNK and Beclin-1 activation | [63,78] |
| Apigenin | Liver cancer | In vivo (rat model) | Inhibits HIF-1α transcription | Induces autophagy by inhibiting mTOR | [64,79] |
| Honokiol | Glioblastoma | In vivo and in vitro | Downregulates HIF-1α and VEGF expression | Triggers autophagy through AMPK activation | [65,80] |
| Withaferin A | Breast cancer | In vitro (MDA-MB-231) | Inhibits HIF-1α expression | Induces autophagy through ROS and ER stress | [66,81] |
| Capsaicin | Lung adenocarcinoma | In vitro (A549 cells) | Reduces HIF-1α transcription | Activates autophagy via AMPK signaling | [67,82] |
| Kaempferol | Ovarian cancer | In vivo (mouse model) | Inhibits HIF-1α signaling | Enhances autophagy via Akt/mTOR inhibition | [68,83] |
| Rhein | Hepatocellular carcinoma | In vitro (HepG2 cells) | Suppresses HIF-1α protein expression | Promotes autophagy through ROS-dependent pathways | [69,84] |
| Luteolin | Colon cancer | In vivo (mouse model) | Inhibits HIF-1α transcription | Induces autophagy by blocking PI3K/Akt signaling | [70,85] |
| Baicalin | Esophageal cancer | In vitro (ECA109 cells) | Downregulates HIF-1α expression | Activates autophagy via AMPK–mTOR signaling | [71,86] |
| Diallyl trisulfide | Gastric cancer | In vivo (mouse model) | Inhibits HIF-1α transcription | Triggers autophagy via inhibition of mTOR signaling | [72,87] |
4.2. Synthetic Drugs for HIF-Mediated Autophagy Regulation in Cancer
4.2.1. HIF-1α Inhibition and Autophagy Suppression
4.2.2. Targeting Hypoxia-Induced Autophagy
Chemotherapeutics Affecting HIF-Mediated Autophagy
Emerging Small Molecules and HIF-1α Modulation
| Synthetic Chemical | Target Pathway | Cancer Type | Concentration/ Dose | Mechanism of Action | Ref. |
| Digoxin | HIF-1α inhibitor | Breast cancer | 50–100 nM | Inhibits HIF-1α expression to reduce autophagy | [88] |
| PX-478 | HIF-1α inhibitor | Glioblastoma | 10–50 µM | Blocks HIF-1α transcription to suppress autophagy | [89] |
| 2-Methoxyestradiol (2-ME) | HIF-1α inhibitor | Pancreatic cancer | 1–10 µM | Destabilizes HIF-1α to reduce autophagy induction | [90] |
| Bortezomib | Proteasome inhibitor | Multiple myeloma | 10–50 nM | Increases HIF-1α degradation to reduce autophagic survival | [91] |
| YC-1 | HIF-1α inhibitor | Colon cancer | 1–50 µM | Inhibits HIF-1α transcriptional activity to reduce autophagy | [92] |
| Tirapazamine | Hypoxia-activated prodrug | Head and Neck cancer | 10–100 µM | Disrupts hypoxia-induced autophagy by suppressing HIF-1α | [93] |
| LW6 | HIF-1α degradation inducer | Lung cancer | 1–10 µM | Promotes HIF-1α proteasomal degradation and reduces autophagy | [94] |
| Vorinostat (SAHA) | HDAC inhibitor | Glioblastoma | 1–10 µM | Inhibits HIF-1α expression and blocks hypoxia-induced autophagy | [95] |
| Doxorubicin | Topoisomerase II inhibitor | Breast cancer | 0.1–5 µM | Suppresses HIF-1α activity and reduces autophagic cell survival | [96] |
| Camptothecin | DNA topoisomerase I inhibitor | Colon cancer | 10–100 nM | Inhibits HIF-1α stabilization, thereby affecting autophagy | [97] |
| Temsirolimus | mTOR inhibitor | Renal cell carcinoma | 10–50 nM | Blocks mTOR and enhances HIF-mediated autophagy | [98] |
| Sorafenib | Multi-kinase inhibitor | Liver cancer | 1–10 µM | Inhibits HIF-1α and downregulates autophagy in hypoxic environment | [99] |
| EZN-2208 | HIF-1α antisense oligonucleotide | Pancreatic cancer | 10–100 nM | Reduces HIF-1α expression and inhibits autophagy | [100] |
| CH5132799 | PI3K/mTOR inhibitor | Breast cancer | 1–10 µM | Suppresses HIF-mediated autophagy under hypoxia | [101] |
4.3. MicroRNAs for HIF-Mediated Autophagy Regulation in Cancer
| MicroRNA | Chemical Name | Cellular Model | HIF-Mediated Mechanism of Action | Autophagy Condition | References |
| miR-21 | microRNA-21 | Cancer cell lines | Upregulation of HIF-1α | Enhanced autophagy | [106] |
| miR-210 | microRNA-210 | Hypoxic cells | Direct targeting of HIF-1α mRNA | Enhanced autophagy | [107] |
| miR-155 | microRNA-155 | Various | Inhibition of HIF-1α translation | Suppressed autophagy | [108] |
| miR-31 | microRNA-31 | Breast cancer cells | Regulation of HIF-1α stability | Enhanced autophagy | [109] |
| miR-29a | microRNA-29a | Lung cancer cells | Modulation of HIF-1α expression | Suppressed autophagy | [110] |
| miR-23a | microRNA-23a | Renal cell carcinoma | Inhibition of HIF-1α pathway | Suppressed autophagy | [111] |
| miR-519c | microRNA-519c | Gastric cancer cells | Targeting HIF-1α | Suppressed autophagy | [112] |
| miR-20a | microRNA-20a | Colon cancer cells | Downregulation of HIF-1α | Enhanced autophagy | [113] |
| miR-424 | microRNA-424 | Breast cancer cells | Regulation of HIF-1α expression | Suppressed autophagy | [114] |
| miR-27a | microRNA-27a | Pancreatic cancer | Modulation of HIF-1α activity | Enhanced autophagy | [115] |
4.4. Nanoparticle-Mediated HIF-Induced Autophagy Regulation in Cancer
5. Future Perspectives and Challenges of Hypoxia-Inducible Factor-Induced Autophagy Regulation in Cancer
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Emami Nejad, A.; Najafgholian, S.; Rostami, A.; Sistani, A.; Shojaeifar, S.; Esparvarinha, M.; Nedaeinia, R.; Haghjooy Javanmard, S.; Taherian, M.; Ahmadlou, M. The role of hypoxia in the tumor microenvironment and development of cancer stem cell: A novel approach to developing treatment. Cancer Cell Int. 2021, 21, 62. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lan, F.; Liu, M.; Wang, F.; Ning, X.; Yang, H.; Sun, H. Hypoxia inducible factor-1ɑ as a potential therapeutic target for osteosarcoma metastasis. Front. Pharmacol. 2024, 15, 1350187. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Ruan, W.; Bobrow, B.; Carmeliet, P.; Eltzschig, H.K. Targeting hypoxia-inducible factors: Therapeutic opportunities and challenges. Nat. Rev. Drug Discov. 2024, 23, 175–200. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Lin, J.; Dai, Z.; Lin, B.; Zhang, J. Hypoxia-associated autophagy flux dysregulation in human cancers. Cancer Lett. 2024, 590, 216823. [Google Scholar] [CrossRef]
- Bae, T.; Hallis, S.P.; Kwak, M.-K. Hypoxia, oxidative stress, and the interplay of HIFs and NRF2 signaling in cancer. Exp. Mol. Med. 2024, 56, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, T.M.; Audero, M.M.; Greco, M.R.; Ardone, M.; Maggi, T.; Mallamaci, R.; Rolando, B.; Arpicco, S.; Ruffinatti, F.A.; Pla, A.F. Tumor Microenvironment Modulates Invadopodia Activity of Non-Selected and Acid-Selected Pancreatic Cancer Cells and Its Sensitivity to Gemcitabine and C18-Gemcitabine. Cells 2024, 13, 730. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.R.; Rahman, M.M.; Islam, M.N.; Fahim, M.M.H.; Rahman, M.A.; Kim, B. Antioxidants activities of phytochemicals perspective modulation of autophagy and apoptosis to treating cancer. Biomed. Pharmacother. 2024, 174, 116497. [Google Scholar] [CrossRef] [PubMed]
- Musleh Ud Din, S.; Streit, S.G.; Huynh, B.T.; Hana, C.; Abraham, A.-N.; Hussein, A. Therapeutic Targeting of Hypoxia-Inducible Factors in Cancer. Int. J. Mol. Sci. 2024, 25, 2060. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.S.; Mishra, R.; Patil, V.M.; Dewangan, S. Role of natural secondary metabolites as HIF-1 inhibitors in cancer therapy. Med. Chem. Res. 2024, 33, 721–734. [Google Scholar] [CrossRef]
- Lee, P.W.T.; Koseki, L.R.; Haitani, T.; Harada, H.; Kobayashi, M. Hypoxia-Inducible Factor-Dependent and Independent Mechanisms Underlying Chemoresistance of Hypoxic Cancer Cells. Cancers 2024, 16, 1729. [Google Scholar] [CrossRef]
- Vongthip, W.; Nilkhet, S.; Boonruang, K.; Sukprasansap, M.; Tencomnao, T.; Baek, S.J. Neuroprotective mechanisms of luteolin in glutamate-induced oxidative stress and autophagy-mediated neuronal cell death. Sci. Rep. 2024, 14, 7707. [Google Scholar] [CrossRef] [PubMed]
- Ortmann, B.M. Hypoxia-inducible factor in cancer: From pathway regulation to therapeutic opportunity. BMJ Oncol. 2024, 3, e000154. [Google Scholar] [CrossRef] [PubMed]
- Zamanian, M.Y.; Golmohammadi, M.; Yumashev, A.; Hjazi, A.; Toama, M.A.; AbdRabou, M.A.; Gehlot, A.; Alwaily, E.R.; Shirsalimi, N.; Yadav, P.K. Effects of metformin on cancers in experimental and clinical studies: Focusing on autophagy and AMPK/mTOR Signaling Pathways. Cell Biochem. Funct. 2024, 42, e4071. [Google Scholar] [CrossRef] [PubMed]
- Imanaga, H.; Semba, Y.; Sasaki, K.; Setoguchi, K.; Maniriho, H.; Yamauchi, T.; Terasaki, T.; Hirabayashi, S.; Nakao, F.; Nogami, J. Central role of the mTORC1 pathway in glucocorticoid activity against B-ALL cells. Blood Neoplasia 2024, 1, 100015. [Google Scholar] [CrossRef]
- Zhao, Y.; Xing, C.; Deng, Y.; Ye, C.; Peng, H. HIF-1α signaling: Essential roles in tumorigenesis and implications in targeted therapies. Genes Dis. 2023, 11, 234–251. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Tsuyama, T.; Sato, Y. Roles of β-Cell Hypoxia in the Progression of Type 2 Diabetes. Int. J. Mol. Sci. 2024, 25, 4186. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-A.; Cho, G.-J.; Kim, D.; Kim, D.-H. Biophysical interplay between extracellular matrix remodeling and hypoxia signaling in regulating cancer metastasis. Front. Cell Dev. Biol. 2024, 12, 1335636. [Google Scholar] [CrossRef] [PubMed]
- Fiorini, G.; Schofield, C.J. Biochemistry of the hypoxia-inducible factor hydroxylases. Curr. Opin. Chem. Biol. 2024, 79, 102428. [Google Scholar] [CrossRef]
- Mitroshina, E.V.; Vedunova, M.V. The Role of Oxygen Homeostasis and the HIF-1 Factor in the Development of Neurodegeneration. Int. J. Mol. Sci. 2024, 25, 4581. [Google Scholar] [CrossRef]
- eun Lee, Y.; Lee, S.-H.; Kim, W.-U. Cytokines, vascular endothelial growth factors, and PlGF in autoimmunity: Insights from rheumatoid arthritis to multiple sclerosis. Immune Netw. 2024, 24, e10. [Google Scholar]
- Huang, M.; Liu, M.; Wang, R.; Man, Y.; Zhou, H.; Xu, Z.-X.; Wang, Y. The crosstalk between glucose metabolism and telomerase regulation in cancer. Biomed. Pharmacother. 2024, 175, 116643. [Google Scholar] [CrossRef]
- Mathew, M.; Nguyen, N.T.; Bhutia, Y.D.; Sivaprakasam, S.; Ganapathy, V. Metabolic signature of Warburg effect in cancer: An effective and obligatory interplay between nutrient transporters and catabolic/anabolic pathways to promote tumor growth. Cancers 2024, 16, 504. [Google Scholar] [CrossRef] [PubMed]
- Akl, M.M.; Ahmed, A. Exploring the Interplay between the Warburg Effect and Glucolipotoxicity in Cancer Development: A Novel Perspective on Cancer Etiology. Adv. Pharm. Bull. 2024, 14, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.S.; Zheng, A.; Xiao, Q. Mitochondrial Reactive Oxygen Species Dysregulation in Heart Failure with Preserved Ejection Fraction: A Fraction of the Whole. Antioxidants 2024, 13, 1330. [Google Scholar] [CrossRef]
- Rahman, M.A.; Yadab, M.K.; Ali, M.M. Emerging role of extracellular pH in tumor microenvironment as a therapeutic target for cancer immunotherapy. Cells 2024, 13, 1924. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Virmani, T.; Kumar, G.; Sharma, A.; Virmani, R.; Gugulothu, D.; Singh, K.; Misra, S.K.; Pathak, K.; Chitranshi, N. Mitochondrial signaling pathways and their role in cancer drug resistance. Cell. Signal. 2024, 122, 111329. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Li, W.; Du, R.; Liu, M.; Cheng, Y.; Ma, J.; Yan, S. Impact of glycolysis enzymes and metabolites in regulating DNA damage repair in tumorigenesis and therapy. Cell Commun. Signal. 2025, 23, 44. [Google Scholar] [CrossRef]
- Zhang, Q.; Dunbar, K.B.; Odze, R.D.; Agoston, A.T.; Wang, X.; Su, T.; Nguyen, A.D.; Zhang, X.; Spechler, S.J.; Souza, R.F. Hypoxia-inducible factor-1α mediates reflux-induced epithelial-mesenchymal plasticity in Barrett’s oesophagus patients. Gut 2024, 73, 1269–1279. [Google Scholar] [CrossRef]
- San, S.H.; Ngai, S.C. E-cadherin re-expression: Its potential in combating TRAIL resistance and reversing epithelial-to-mesenchymal transition. Gene 2024, 909, 148293. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Campa, C.; Álvarez-García, V.; Alonso-González, C.; González, A.; Cos, S. Melatonin and Its Role in the Epithelial-to-Mesenchymal Transition (EMT) in Cancer. Cancers 2024, 16, 956. [Google Scholar] [CrossRef]
- Bigos, K.J.; Quiles, C.G.; Lunj, S.; Smith, D.J.; Krause, M.; Troost, E.G.; West, C.M.; Hoskin, P.; Choudhury, A. Tumour response to hypoxia: Understanding the hypoxic tumour microenvironment to improve treatment outcome in solid tumours. Front. Oncol. 2024, 14, 1331355. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhang, M.; Xu, T.; Xue, S.; Li, D.; Zhao, Y.; Zhi, F.; Ding, D. Enhancing photodynamic immunotherapy by reprograming the immunosuppressive tumor microenvironment with hypoxia relief. J. Control. Release 2024, 368, 233–250. [Google Scholar] [CrossRef]
- Rezaie, J.; Chodari, L.; Mohammadpour-Asl, S.; Jafari, A.; Niknam, Z. Cell-mediated barriers in cancer immunosurveillance. Life Sci. 2024, 342, 122528. [Google Scholar] [CrossRef]
- Totu, T.; Bossart, J.; Hast, K.; Li, C.; Rottmar, M.; Sobottka, B.; Yu, G.; Ayala-Nunez, V.; Buljan, M. Delineation of signaling routes that underlie differences in macrophage phenotypic states. bioRxiv 2024. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, P.; Yin, N.; Xu, Z.; Liu, X.; Wu, A.; Zhang, X.; Li, Z.; Zhang, Z.; Zhong, T. Glioblastoma stem cell-derived exosomal miR-374b-3p promotes tumor angiogenesis and progression through inducing M2 macrophages polarization. iScience 2024, 27, 109270. [Google Scholar] [CrossRef]
- Chamani, F.K.; Etebari, A.; Hajivalili, M.; Mosaffa, N.; Jalali, S.A. Hypoxia and programmed cell death-ligand 1 expression in the tumor microenvironment: A review of the effects of hypoxia-induced factor-1 on immunotherapy. Mol. Biol. Rep. 2024, 51, 88. [Google Scholar] [CrossRef]
- Krishnan, S.; Shrestha, Y.; Jayatunga, D.P.W.; Rea, S.; Martins, R.; Bharadwaj, P. Activate or Inhibit? Implications of Autophagy Modulation as a Therapeutic Strategy for Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 6739. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Rhim, H. Therapeutic implication of autophagy in neurodegenerative diseases. BMB Rep. 2017, 50, 345–354. [Google Scholar] [CrossRef]
- Hurley, J.H.; Young, L.N. Mechanisms of Autophagy Initiation. Annu. Rev. Biochem. 2017, 86, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Kotani, T.; Kirisako, H.; Koizumi, M.; Ohsumi, Y.; Nakatogawa, H. The Atg2-Atg18 complex tethers pre-autophagosomal membranes to the endoplasmic reticulum for autophagosome formation. Proc. Natl. Acad. Sci. USA 2018, 115, 10363–10368. [Google Scholar] [CrossRef] [PubMed]
- Alers, S.; Loffler, A.S.; Wesselborg, S.; Stork, B. Role of AMPK-mTOR-Ulk1/2 in the Regulation of Autophagy: Cross Talk, Shortcuts, and Feedbacks. Mol. Cell. Biol. 2012, 32, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, A.F.C.; Jackson, W.T. So Many Roads: The Multifaceted Regulation of Autophagy Induction. Mol. Cell. Biol. 2018, 38, e00303-18. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Shpilka, T.; Elazar, Z. Mechanisms of Autophagosome Biogenesis. Curr. Biol. 2012, 22, R29–R34. [Google Scholar] [CrossRef]
- Kardideh, B.; Samimi, Z.; Norooznezhad, F.; Kiani, S.; Mansouri, K. Autophagy, cancer and angiogenesis: Where is the link? Cell Biosci. 2019, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Rahman, M.S.; Rahman, M.H.; Rasheduzzaman, M.; Mamun-Or-Rashid, A.N.M.; Uddin, M.J.; Rahman, M.R.; Hwang, H.; Pang, M.G.; Rhim, H. Modulatory Effects of Autophagy on APP Processing as a Potential Treatment Target for Alzheimer’s Disease. Biomedicines 2020, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Huang, J. Role of estrogen and its receptors mediated-autophagy in cell fate and human diseases. J. Steroid Biochem. Mol. Biol. 2019, 191, 105380. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Rahman, M.H.; Hossain, M.S.; Biswas, P.; Islam, R.; Uddin, M.J.; Rahman, M.H.; Rhim, H. Molecular Insights into the Multifunctional Role of Natural Compounds: Autophagy Modulation and Cancer Prevention. Biomedicines 2020, 8, 517. [Google Scholar] [CrossRef] [PubMed]
- Mialet-Perez, J.; Belaidi, E. Interplay between Hypoxia Inducible Factor-1 and mitochondria in cardiac diseases. Free Radic. Biol. Med. 2024, 221, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Biswas, U.; Roy, R.; Ghosh, S.; Chakrabarti, G. The interplay between autophagy and apoptosis: Its implication in lung cancer and therapeutics. Cancer Lett. 2024, 585, 216662. [Google Scholar] [CrossRef]
- Zheng, Y.; Zha, X.; Zhang, B.; Elsabagh, M.; Wang, H.; Wang, M.; Zhang, H. The interaction of ER stress and autophagy in trophoblasts: Navigating pregnancy outcome. Biol. Reprod. 2024, 111, 292–311. [Google Scholar] [CrossRef]
- Danish, F.; Qureshi, M.A.; Mirza, T.; Amin, W.; Sufiyan, S.; Naeem, S.; Arshad, F.; Mughal, N. Investigating the Association between the Autophagy Markers LC3B, SQSTM1/p62, and DRAM and Autophagy-Related Genes in Glioma. Int. J. Mol. Sci. 2024, 25, 572. [Google Scholar] [CrossRef] [PubMed]
- Ting, K.K. Revisiting the role of hypoxia-inducible factors and nuclear factor erythroid 2-related factor 2 in regulating macrophage inflammation and metabolism. Front. Cell. Infect. Microbiol. 2024, 14, 1403915. [Google Scholar] [CrossRef] [PubMed]
- Mitaishvili, E.; Feinsod, H.; David, Z.; Shpigel, J.; Fernandez, C.; Sauane, M.; de la Parra, C. The Molecular Mechanisms behind Advanced Breast Cancer Metabolism: Warburg Effect, OXPHOS, and Calcium. Front. Biosci. 2024, 29, 99. [Google Scholar] [CrossRef] [PubMed]
- Zaarour, R.F.; Azakir, B.; Hajam, E.Y.; Nawafleh, H.; Zeinelabdin, N.A.; Engelsen, A.S.; Thiery, J.; Jamora, C.; Chouaib, S. Role of hypoxia-mediated autophagy in tumor cell death and survival. Cancers 2021, 13, 533. [Google Scholar] [CrossRef] [PubMed]
- Troise, D.; Infante, B.; Mercuri, S.; Piccoli, C.; Lindholm, B.; Stallone, G. Hypoxic Inducible Factor Stabilization in Pericytes beyond Erythropoietin Production: The Good and the Bad. Antioxidants 2024, 13, 537. [Google Scholar] [CrossRef]
- He, K.; Chen, M.; Liu, J.; Du, S.; Ren, C.; Zhang, J. Nanomedicine for cancer targeted therapy with autophagy regulation. Front. Immunol. 2024, 14, 1238827. [Google Scholar] [CrossRef]
- Hamed, A.A.; Ghareeb, M.A.; Soliman, N.R.; Bakchiche, B.; Bardaweel, S.K. Insights into bioactive microbial natural products and drug discovery. Egypt. Pharm. J. 2024, 23, 1–15. [Google Scholar] [CrossRef]
- Samec, M.; Liskova, A.; Koklesova, L.; Mersakova, S.; Strnadel, J.; Kajo, K.; Pec, M.; Zhai, K.; Smejkal, K.; Mirzaei, S. Flavonoids targeting HIF-1: Implications on cancer metabolism. Cancers 2021, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Benameur, T.; Giacomucci, G.; Panaro, M.A.; Ruggiero, M.; Trotta, T.; Monda, V.; Pizzolorusso, I.; Lofrumento, D.D.; Porro, C.; Messina, G. New promising therapeutic avenues of curcumin in brain diseases. Molecules 2022, 27, 236. [Google Scholar] [CrossRef] [PubMed]
- Dhatwalia, S.K.; Kumar, M.; Dhawan, D.K. Role of EGCG in containing the progression of lung tumorigenesis—A multistage targeting approach. Nutr. Cancer 2018, 70, 334–349. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; Alsahli, M.A.; Rahmani, A.H. Berberine: An important emphasis on its anticancer effects through modulation of various cell signaling pathways. Molecules 2022, 27, 5889. [Google Scholar] [CrossRef]
- Hu, Y.; Li, R.; Jin, J.; Wang, Y.; Ma, R. Quercetin improves pancreatic cancer chemo-sensitivity by regulating oxidative-inflammatory networks. J. Food Biochem. 2022, 46, e14453. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Di Domizio, A.; Limonta, P. Natural compounds in prostate cancer prevention and treatment: Mechanisms of action and molecular targets. Cells 2020, 9, 460. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Tian, D.; Liao, X.; Zhang, Y.; Xiao, J.; Chen, W.; Liu, Q.; Chen, Y.; Li, D.; Zhu, L. Apigenin combined with gefitinib blocks autophagy flux and induces apoptotic cell death through inhibition of HIF-1α, c-Myc, p-EGFR, and glucose metabolism in EGFR L858R+ T790M-mutated H1975 cells. Front. Pharmacol. 2019, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.-X.; Li, Y.; Liu, Z.-Q.; Fan, X.-X.; Duan, F.-G.; Li, R.-Z.; Yao, X.-J.; Leung, E.L.-H.; Liu, L. Honokiol induces apoptosis, G1 arrest, and autophagy in KRAS mutant lung cancer cells. Front. Pharmacol. 2017, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kim, H.; Yuan, T.; Zhang, Z.; Kaul, S.C.; Wadhwa, R. Molecular Characterization of Cancer Preventive and Therapeutic Potential of Three Antistress Compounds, Triethylene Glycol, Withanone, and Withaferin A. Int. J. Mol. Sci. 2025, 26, 493. [Google Scholar] [CrossRef]
- Kammath, A.J.; Nair, B.; P, S.; Nath, L.R. Curry versus cancer: Potential of some selected culinary spices against cancer with in vitro, in vivo, and human trials evidences. J. Food Biochem. 2021, 45, e13285. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Rankin, G.O.; Liu, L.; Daddysman, M.K.; Jiang, B.-H.; Chen, Y.C. Kaempferol inhibits angiogenesis and VEGF expression through both HIF dependent and independent pathways in human ovarian cancer cells. Nutr. Cancer 2009, 61, 554–563. [Google Scholar] [CrossRef]
- Fernand, V.E.; Losso, J.N.; Truax, R.E.; Villar, E.E.; Bwambok, D.K.; Fakayode, S.O.; Lowry, M.; Warner, I.M. Rhein inhibits angiogenesis and the viability of hormone-dependent and-independent cancer cells under normoxic or hypoxic conditions in vitro. Chem. Biol. Interact. 2011, 192, 220–232. [Google Scholar] [CrossRef]
- Çetinkaya, M.; Baran, Y. Therapeutic potential of luteolin on cancer. Vaccines 2023, 11, 554. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Peng, C.; Zheng, C.; He, X.-H.; Huang, W.; Han, B. Recent advances in characterizing natural products that regulate autophagy. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem.-Anti-Cancer Agents) 2019, 19, 2177–2196. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.D.; Son, S.W.; Choi, S.Y.; Kuh, H.J.; Oh, T.-J.; Park, J.K. Anti-cancer activity of phytochemicals targeting hypoxia-inducible factor-1 alpha. Int. J. Mol. Sci. 2021, 22, 9819. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Lertpiriyapong, K.; Steelman, L.S.; Abrams, S.L.; Yang, L.V.; Murata, R.M.; Rosalen, P.L.; Scalisi, A.; Neri, L.M.; Cocco, L. Effects of resveratrol, curcumin, berberine and other nutraceuticals on aging, cancer development, cancer stem cells and microRNAs. Aging 2017, 9, 1477–1536. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Scott, J.; Sengupta, D.; Al-Gharaibeh, A.; Dunbar, G.L. Curcumin and solid lipid curcumin particles induce autophagy, but inhibit mitophagy and the PI3K-Akt/mTOR pathway in cultured glioblastoma cells. Int. J. Mol. Sci. 2019, 20, 399. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Pezzani, R.; Redaelli, M.; Zorzan, M.; Imran, M.; Ahmed Khalil, A.; Salehi, B.; Sharopov, F.; Cho, W.C.; Sharifi-Rad, J. Preclinical activities of epigallocatechin gallate in signaling pathways in cancer. Molecules 2020, 25, 467. [Google Scholar] [CrossRef]
- Davoodvandi, A.; Sadeghi, S.; Alavi, S.M.A.; Alavi, S.S.; Jafari, A.; Khan, H.; Aschner, M.; Mirzaei, H.; Sharifi, M.; Asemi, Z. The therapeutic effects of berberine for gastrointestinal cancers. Asia-Pac. J. Clin. Oncol. 2024, 20, 152–167. [Google Scholar] [CrossRef]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR-and hypoxia-induced factor 1α-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, M.; Albonici, L.; Focaccetti, C.; Ciuffa, S.; Fazi, S.; Cifaldi, L.; Miele, M.T.; De Maio, F.; Tresoldi, I.; Manzari, V. Polyphenol-mediated autophagy in cancer: Evidence of in vitro and in vivo studies. Int. J. Mol. Sci. 2020, 21, 6635. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Lee, H.G. Apigenin induces autophagy and cell death by targeting EZH2 under hypoxia conditions in gastric cancer cells. Int. J. Mol. Sci. 2021, 22, 13455. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.; Lee, W.; Tang, Y.; Yap, W. Honokiol: A Review of Its Anticancer Potential and Mechanisms. Cancers 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; De, S.; Mukherjee, S.; Das, S.; Ghosh, A.N.; Sengupta, S.B. Withaferin A induced impaired autophagy and unfolded protein response in human breast cancer cell-lines MCF-7 and MDA-MB-231. Toxicol. In Vitro 2017, 44, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Han, T.-H.; Park, M.K.; Nakamura, H.; Ban, H.S. Capsaicin inhibits HIF-1α accumulation through suppression of mitochondrial respiration in lung cancer cells. Biomed. Pharmacother. 2022, 146, 112500. [Google Scholar] [CrossRef]
- Huang, W.-W.; Tsai, S.-C.; Peng, S.-F.; Lin, M.-W.; Chiang, J.-H.; Chiu, Y.-J.; Fushiya, S.; Tseng, M.T.; Yang, J.-S. Kaempferol induces autophagy through AMPK and AKT signaling molecules and causes G2/M arrest via downregulation of CDK1/cyclin B in SK-HEP-1 human hepatic cancer cells. Int. J. Oncol. 2013, 42, 2069–2077. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Jiang, H.; Liu, Y.; Chen, J.; Zhou, X.; Zhao, C.; Chen, X.; Lin, M. Rhein induces liver cancer cells apoptosis via activating ROS-dependent JNK/Jun/caspase-3 signaling pathway. J. Cancer 2020, 11, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Raina, R.; Pramodh, S.; Rais, N.; Haque, S.; Shafarin, J.; Bajbouj, K.; Hamad, M.; Hussain, A. Luteolin inhibits proliferation, triggers apoptosis and modulates Akt/mTOR and MAP kinase pathways in HeLa cells. Oncol. Lett. 2021, 21, 1. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Relitti, N.; Brindisi, M.; Magnano, S.; Zisterer, D.; Gemma, S.; Butini, S.; Campiani, G. Autophagy modulators for the treatment of oral and esophageal squamous cell carcinomas. Med. Res. Rev. 2020, 40, 1002–1060. [Google Scholar] [CrossRef]
- Lu, L.; Gao, Z.; Song, J.; Jin, L.; Liang, Z. The potential of diallyl trisulfide for cancer prevention and treatment, with mechanism insights. Front. Cell Dev. Biol. 2024, 12, 1450836. [Google Scholar] [CrossRef]
- Zhang, H.; Qian, D.Z.; Tan, Y.S.; Lee, K.; Gao, P.; Ren, Y.R.; Rey, S.; Hammers, H.; Chang, D.; Pili, R. Digoxin and other cardiac glycosides inhibit HIF-1α synthesis and block tumor growth. Proc. Natl. Acad. Sci. USA 2008, 105, 19579–19586. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, J.J.; Erez, B.; Korshunova, M.V.; Williams, R.R.; Furutani, K.; Takahashi, O.; Kirkpatrick, L.; Lippman, S.M.; Powis, G.; O’Reilly, M.S. Treatment with HIF-1α antagonist PX-478 inhibits progression and spread of orthotopic human small cell lung cancer and lung adenocarcinoma in mice. J. Thorac. Oncol. 2010, 5, 940–949. [Google Scholar] [CrossRef]
- Batth, I.S.; Huang, S.-B.; Villarreal, M.; Gong, J.; Chakravarthy, D.; Keppler, B.; Jayamohan, S.; Osmulski, P.; Xie, J.; Rivas, P. Evidence for 2-methoxyestradiol-mediated inhibition of receptor tyrosine kinase RON in the management of prostate cancer. Int. J. Mol. Sci. 2021, 22, 1852. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Engelsen, A.S.; Sarowar, S.; Bindesbøll, C.; Birkeland, E.; Goplen, D.; Lotsberg, M.L.; Knappskog, S.; Simonsen, A.; Chekenya, M. Bortezomib abrogates temozolomide-induced autophagic flux through an ATG5 dependent pathway. Front. Cell Dev. Biol. 2022, 10, 1022191. [Google Scholar] [CrossRef] [PubMed]
- Selvakumaran, M.; Amaravadi, R.K.; Vasilevskaya, I.A.; O’Dwyer, P.J. Autophagy inhibition sensitizes colon cancer cells to antiangiogenic and cytotoxic therapy. Clin. Cancer Res. 2013, 19, 2995–3007. [Google Scholar] [CrossRef]
- Hu, Y.-L.; DeLay, M.; Jahangiri, A.; Molinaro, A.M.; Rose, S.D.; Carbonell, W.S.; Aghi, M.K. Hypoxia-induced autophagy promotes tumor cell survival and adaptation to antiangiogenic treatment in glioblastoma. Cancer Res. 2012, 72, 1773–1783. [Google Scholar] [CrossRef]
- Lee, K.; Kang, J.E.; Park, S.-K.; Jin, Y.; Chung, K.-S.; Kim, H.-M.; Lee, K.; Kang, M.R.; Lee, M.K.; Song, K.B. LW6, a novel HIF-1 inhibitor, promotes proteasomal degradation of HIF-1α via upregulation of VHL in a colon cancer cell line. Biochem. Pharmacol. 2010, 80, 982–989. [Google Scholar] [CrossRef]
- Hutt, D.M.; Roth, D.M.; Vignaud, H.; Cullin, C.; Bouchecareilh, M. The histone deacetylase inhibitor, Vorinostat, represses hypoxia inducible factor 1 alpha expression through translational inhibition. PLoS ONE 2014, 9, e106224. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Eble, J.M.; Moon, E.; Yuan, H.; Weitzel, D.H.; Landon, C.D.; Yu-Chih Nien, C.; Hanna, G.; Rich, J.N.; Provenzale, J.M. Tumor cells upregulate normoxic HIF-1α in response to doxorubicin. Cancer Res. 2013, 73, 6230–6242. [Google Scholar] [CrossRef]
- Bertozzi, D.; Marinello, J.; Manzo, S.G.; Fornari, F.; Gramantieri, L.; Capranico, G. The natural inhibitor of DNA topoisomerase I, camptothecin, modulates HIF-1α activity by changing miR expression patterns in human cancer cells. Mol. Cancer Ther. 2014, 13, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Le Tourneau, C.; Faivre, S.; Serova, M.; Raymond, E. mTORC1 inhibitors: Is temsirolimus in renal cancer telling us how they really work? Br. J. Cancer 2008, 99, 1197–1203. [Google Scholar] [CrossRef]
- Yang, Q.; Gao, L.; Huang, X.; Weng, J.; Chen, Y.; Lin, S.; Yin, Q. Sorafenib prevents the proliferation and induces the apoptosis of liver cancer cells by regulating autophagy and hypoxia-inducible factor-1. Exp. Ther. Med. 2021, 22, 980. [Google Scholar]
- Sapra, P.; Kraft, P.; Pastorino, F.; Ribatti, D.; Dumble, M.; Mehlig, M.; Wang, M.; Ponzoni, M.; Greenberger, L.M.; Horak, I.D. Potent and sustained inhibition of HIF-1α and downstream genes by a polyethyleneglycol-SN38 conjugate, EZN-2208, results in anti-angiogenic effects. Angiogenesis 2011, 14, 245–253. [Google Scholar] [PubMed]
- Peng, Y.; Wang, Y.; Zhou, C.; Mei, W.; Zeng, C. PI3K/Akt/mTOR pathway and its role in cancer therapeutics: Are we making headway? Front. Oncol. 2022, 12, 819128. [Google Scholar] [CrossRef]
- Pandey, S.; Singh, R.; Habib, N.; Tripathi, R.M.; Kushwaha, R.; Mahdi, A.A. Regulation of Hypoxia Dependent Reprogramming of Cancer Metabolism: Role of HIF-1 and Its Potential Therapeutic Implications in Leukemia. Asian Pac. J. Cancer Prev. 2024, 25, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-W.; Zheng, S.-Q.; Li, T.; Fei, Y.-F.; Wang, C.; Zhang, S.; Wang, F.; Jiang, G.-M.; Wang, H. RNA modifications in cellular metabolism: Implications for metabolism-targeted therapy and immunotherapy. Signal Transduct. Target. Ther. 2024, 9, 70. [Google Scholar] [CrossRef]
- Bhattacharjee, B.; Syeda, A.F.; Rynjah, D.; Hussain, S.M.; Chandra Bora, S.; Pegu, P.; Sahu, R.K.; Khan, J. Pharmacological impact of microRNAs in head and neck squamous cell carcinoma: Prevailing insights on molecular pathways, diagnosis, and nanomedicine treatment. Front. Pharmacol. 2023, 14, 1174330. [Google Scholar] [CrossRef] [PubMed]
- Ashrafizadeh, M.; Zarrabi, A.; Orouei, S.; Hushmandi, K.; Hakimi, A.; Zabolian, A.; Daneshi, S.; Samarghandian, S.; Baradaran, B.; Najafi, M. MicroRNA-mediated autophagy regulation in cancer therapy: The role in chemoresistance/chemosensitivity. Eur. J. Pharmacol. 2021, 892, 173660. [Google Scholar] [CrossRef]
- He, C.; Dong, X.; Zhai, B.; Jiang, X.; Dong, D.; Li, B.; Jiang, H.; Xu, S.; Sun, X. MiR-21 mediates sorafenib resistance of hepatocellular carcinoma cells by inhibiting autophagy via the PTEN/Akt pathway. Oncotarget 2015, 6, 28867–28881. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xing, X.; Liu, Q.; Wang, Z.; Xin, Y.; Zhang, P.; Hu, C.; Liu, Y. Hypoxia-induced autophagy reduces radiosensitivity by the HIF-1α/miR-210/Bcl-2 pathway in colon cancer cells. Int. J. Oncol. 2015, 46, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Bruning, U.; Cerone, L.; Neufeld, Z.; Fitzpatrick, S.F.; Cheong, A.; Scholz, C.C.; Simpson, D.A.; Leonard, M.O.; Tambuwala, M.M.; Cummins, E.P. MicroRNA-155 promotes resolution of hypoxia-inducible factor 1α activity during prolonged hypoxia. Mol. Cell. Biol. 2011, 31, 4087–4096. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Carina, V.; Conigliaro, A.; Raimondi, L.; De Luca, A.; Bellavia, D.; Salamanna, F.; Setti, S.; Alessandro, R.; Fini, M. miR-31-5p is a LIPUS-mechanosensitive microRNA that targets HIF-1α signaling and cytoskeletal proteins. Int. J. Mol. Sci. 2019, 20, 1569. [Google Scholar] [CrossRef] [PubMed]
- Donati, S.; Aurilia, C.; Palmini, G.; Falsetti, I.; Iantomasi, T.; Brandi, M.L. Autophagy-Related ncRNAs in Pancreatic Cancer. Pharmaceuticals 2022, 15, 1547. [Google Scholar] [CrossRef]
- Cui, M.; Liu, Z.; Wang, S.; Bae, S.; Guo, H.; Zhou, J.; Liu, R.; Wang, L. CRISPR-based dissection of microRNA-23a~ 27a~ 24-2 cluster functionality in hepatocellular carcinoma. Oncogene 2024, 43, 2708–2721. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Ma, J.; Zhou, W.; Cao, B.; Zhou, X.; Zhang, H.; Zhao, Q.; Hong, L.; Fan, D. Reciprocal regulations between miRNAs and HIF-1α in human cancers. Cell. Mol. Life Sci. 2019, 76, 453–471. [Google Scholar] [CrossRef]
- Che, J.; Wang, W.; Huang, Y.; Zhang, L.; Zhao, J.; Zhang, P.; Yuan, X. miR-20a inhibits hypoxia-induced autophagy by targeting ATG5/FIP200 in colorectal cancer. Mol. Carcinog. 2019, 58, 1234–1247. [Google Scholar] [CrossRef]
- Yu, M.; Ozaki, T.; Sun, D.; Xing, H.; Wei, B.; An, J.; Yang, J.; Gao, Y.; Liu, S.; Kong, C. HIF-1α-dependent miR-424 induction confers cisplatin resistance on bladder cancer cells through down-regulation of pro-apoptotic UNC5B and SIRT4. J. Exp. Clin. Cancer Res. 2020, 39, 108. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Gao, H.; Xu, R.; Wang, H.; Mei, J.; Liu, C. The interplay between HIF-1α and noncoding RNAs in cancer. J. Exp. Clin. Cancer Res. 2020, 39, 27. [Google Scholar] [CrossRef]
- Jain, S.; Hirst, D.; O’Sullivan, J. Gold nanoparticles as novel agents for cancer therapy. Br. J. Radiol. 2012, 85, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.R.; Sanati, M.; Kesharwani, P.; Sahebkar, A. Recent Advances in Curcumin-Based Combination Nanomedicines for Cancer Therapy. J. Funct. Biomater. 2023, 14, 408. [Google Scholar] [CrossRef] [PubMed]
- Strużyńska, L. Dual Implications of Nanosilver-Induced Autophagy: Nanotoxicity and Anti-Cancer Effects. Int. J. Mol. Sci. 2023, 24, 15386. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-S.; Chen, L.; Xiong, Y.-Q.; Xu, J.; Wang, J.-P.; Meng, Z.-L. Iron oxide magnetic nanoparticles combined with actein suppress non-small-cell lung cancer growth in a p53-dependent manner. Int. J. Nanomed. 2017, 12, 7627–7651. [Google Scholar] [CrossRef]
- Kumbhar, P.R.; Kumar, P.; Lasure, A.; Velayutham, R.; Mandal, D. An updated landscape on nanotechnology-based drug delivery, immunotherapy, vaccinations, imaging, and biomarker detections for cancers: Recent trends and future directions with clinical success. Discov. Nano 2023, 18, 156. [Google Scholar] [CrossRef] [PubMed]
- Andreani, T.; Cheng, R.; Elbadri, K.; Ferro, C.; Menezes, T.; Dos Santos, M.R.; Pereira, C.M.; Santos, H.A. Natural compounds-based nanomedicines for cancer treatment: Future directions and challenges. Drug Deliv. Transl. Res. 2024, 14, 2845–2916. [Google Scholar] [CrossRef]
- Slapak, E.J.; El Mandili, M.; Bijlsma, M.F.; Spek, C.A. Mesoporous silica nanoparticle-based drug delivery systems for the treatment of pancreatic cancer: A systematic literature overview. Pharmaceutics 2022, 14, 390. [Google Scholar] [CrossRef] [PubMed]
- Park, T.; Lee, S.; Amatya, R.; Cheong, H.; Moon, C.; Kwak, H.D.; Min, K.A.; Shin, M.C. ICG-loaded pegylated BSA-silver nanoparticles for effective photothermal cancer therapy. Int. J. Nanomed. 2020, 15, 5459–5471. [Google Scholar] [CrossRef]
- Tosat-Bitrián, C.; Palomo, V. CdSe quantum dots evaluation in primary cellular models or tissues derived from patients. Nanomed. Nanotechnol. Biol. Med. 2020, 30, 102299. [Google Scholar] [CrossRef]
- Wang, T.; He, M.; Zhang, X.; Guo, Z.; Wang, P.; Long, F. Deciphering the impact of circRNA-mediated autophagy on tumor therapeutic resistance: A novel perspective. Cell. Mol. Biol. Lett. 2024, 29, 60. [Google Scholar] [CrossRef] [PubMed]
- Foglia, B.; Novo, E.; Protopapa, F.; Maggiora, M.; Bocca, C.; Cannito, S.; Parola, M. Hypoxia, hypoxia-inducible factors and liver fibrosis. Cells 2021, 10, 1764. [Google Scholar] [CrossRef]
- Soni, S.; Padwad, Y.S. HIF-1 in cancer therapy: Two decade long story of a transcription factor. Acta Oncol. 2017, 56, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xiong, Z.; Wang, J.; Gao, Y.; Cao, Q.; Wang, D.; Shi, J.; Chen, Z.; Yang, X. TBC1D5 reverses the capability of HIF-2α in tumor progression and lipid metabolism in clear cell renal cell carcinoma by regulating the autophagy. J. Transl. Med. 2024, 22, 212. [Google Scholar] [CrossRef] [PubMed]



| Nanoparticle Type | Nanoparticle Composition | Cancer Type | Mechanism of Action | HIF-Mediated Autophagy Regulation | Concentration/ Dose (In Vitro/In Vivo) | Ref. |
| Gold nanoparticles (AuNPs) | Gold | Breast cancer, lung cancer | Oxidative stress induction and activation of autophagy via ROS generation | Modulates HIF-1α activity and autophagy induction | 10–50 µg/mL (in vitro), 10–30 mg/kg (in vivo) | [116] |
| Liposomes | Lipid bilayer (phospholipids) | Glioblastoma, prostate cancer | Delivery of drugs and enhanced uptake by tumor cells | Reduces HIF-1α levels and promotes autophagic degradation | 1–10 µg/mL (in vitro), 5–20 mg/kg (in vivo) | [117] |
| Polymeric nanoparticles | Polylactic-co-glycolic acid (PLGA) | Colon cancer, melanoma | Slow release of therapeutic agents and targeted delivery | Suppresses HIF-1α expression and enhances autophagic flux | 20–100 µg/mL (in vitro), 5–25 mg/kg (in vivo) | [78] |
| Silver nanoparticles (AgNPs) | Silver | Lung cancer, breast cancer | Induction of apoptosis and autophagy via ROS generation | Enhances HIF-1α degradation and upregulates autophagy in cancer cells | 10–50 µg/mL (in vitro), 5–15 mg/kg (in vivo) | [118] |
| Magnetic nanoparticles (MNPs) | Iron oxide | Liver cancer, pancreatic cancer | Magnetic targeting for tumor site-specific drug delivery | Modulates HIF-1α activity and influences autophagic processes | 1–5 µg/mL (in vitro), 5–10 mg/kg (in vivo) | [119] |
| Carbon nanotubes (CNTs) | Carbon | Cervical cancer, leukemia | Facilitation of drug delivery and ROS generation | Inhibits HIF-1α and induces autophagic cell death in tumor cells | 0.1–10 µg/mL (in vitro), 5–20 mg/kg (in vivo) | [120] |
| Dendrimers | Poly(amidoamine) (PAMAM) | Ovarian cancer, brain tumor | Targeted drug delivery and cell uptake enhancement | Reduces HIF-1α levels and promotes autophagic cell death | 1–10 µg/mL (in vitro), 10–30 mg/kg (in vivo) | [121] |
| Mesoporous silica nanoparticles (MSNs) | Silica | Breast cancer, pancreatic cancer | Drug encapsulation and controlled release | Enhances autophagy through HIF-1α suppression | 5–50 µg/mL (in vitro), 10–40 mg/kg (in vivo) | [122] |
| Gold–silver alloy nanoparticles (Au-Ag NPs) | Gold–silver alloy | Colorectal cancer, lung cancer | Combination therapy with photothermal properties | Inhibits HIF-1α and enhances autophagic clearance | 5–50 µg/mL (in vitro), 10–30 mg/kg (in vivo) | [123] |
| Quantum dots (QDs) | Semiconductor materials (CdSe) | Breast cancer, prostate cancer | Fluorescence for tracking drug delivery | Inhibits HIF-1α and accelerates autophagy-mediated cell death | 0.1–1 µg/mL (in vitro), 5–15 mg/kg (in vivo) | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalouli, M. Emerging Role of Hypoxia-Inducible Factors (HIFs) in Modulating Autophagy: Perspectives on Cancer Therapy. Int. J. Mol. Sci. 2025, 26, 1752. https://doi.org/10.3390/ijms26041752
Jalouli M. Emerging Role of Hypoxia-Inducible Factors (HIFs) in Modulating Autophagy: Perspectives on Cancer Therapy. International Journal of Molecular Sciences. 2025; 26(4):1752. https://doi.org/10.3390/ijms26041752
Chicago/Turabian StyleJalouli, Maroua. 2025. "Emerging Role of Hypoxia-Inducible Factors (HIFs) in Modulating Autophagy: Perspectives on Cancer Therapy" International Journal of Molecular Sciences 26, no. 4: 1752. https://doi.org/10.3390/ijms26041752
APA StyleJalouli, M. (2025). Emerging Role of Hypoxia-Inducible Factors (HIFs) in Modulating Autophagy: Perspectives on Cancer Therapy. International Journal of Molecular Sciences, 26(4), 1752. https://doi.org/10.3390/ijms26041752