
Effect of MiRNA 204-5P Mimics and Lipopolysaccharide-Induced Inflammation on Transcription Factor Levels, Cell Maintenance, and Retinoic Acid Signaling in Primary Limbal Epithelial Cells
 , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Results
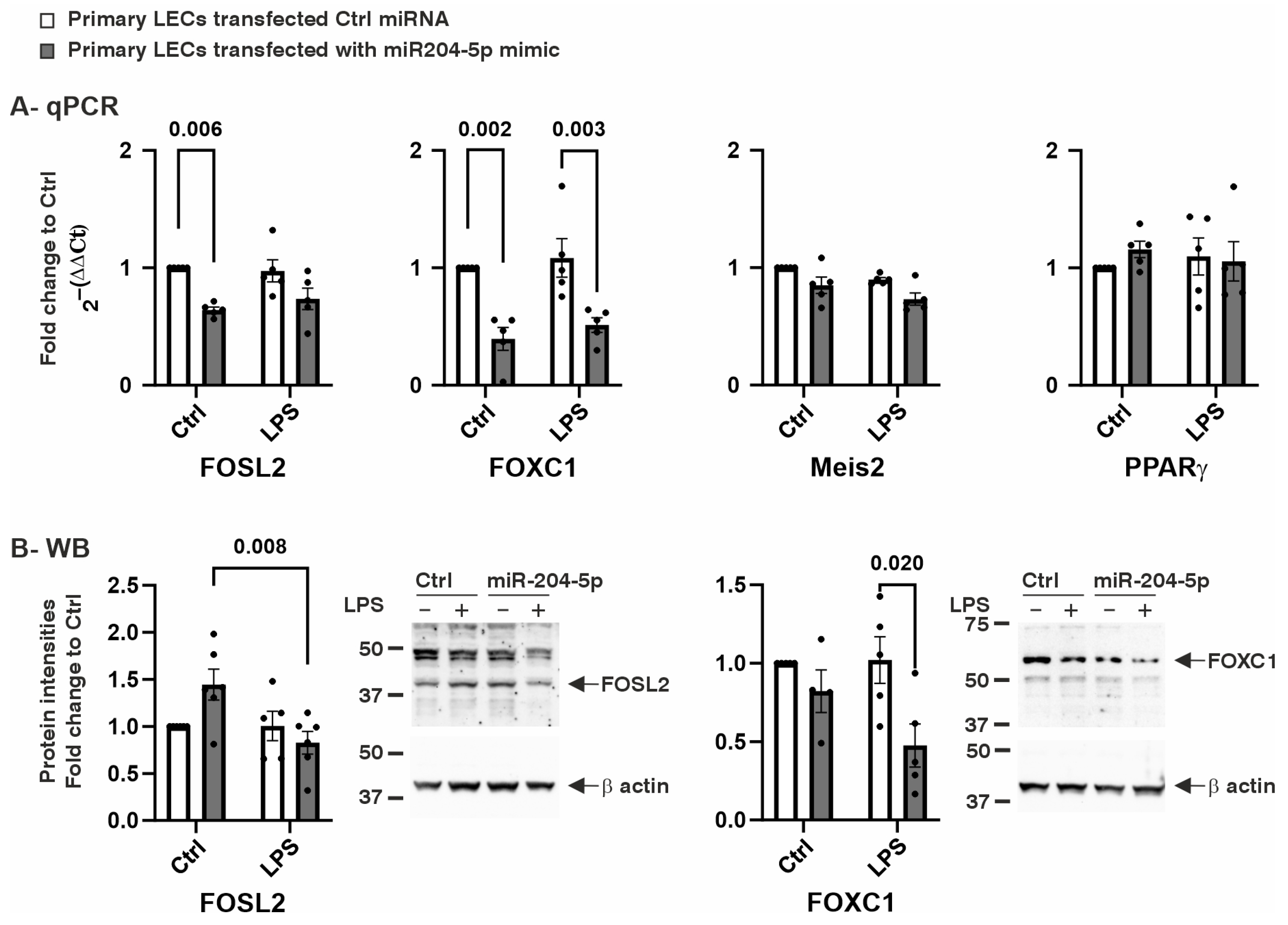
2.1. The Effect of miR204-5p Mimics on Cell Proliferation, mRNA, and Protein Levels of Transcription Factors
2.2. The Effects of the miR204-5p Mimics on the mRNA and Protein Levels of Genes Involved in Cell Maintenance and Inflammation
2.3. The Effects of the miR204-5p Mimics on the mRNA and Protein Levels of Genes Involved in Maintaining Cell Structure and Matrix Remodeling
2.4. The Effects of the miR204-5p Mimics on the mRNA and Protein Levels of Genes Involved in Retinoic Acid Signaling
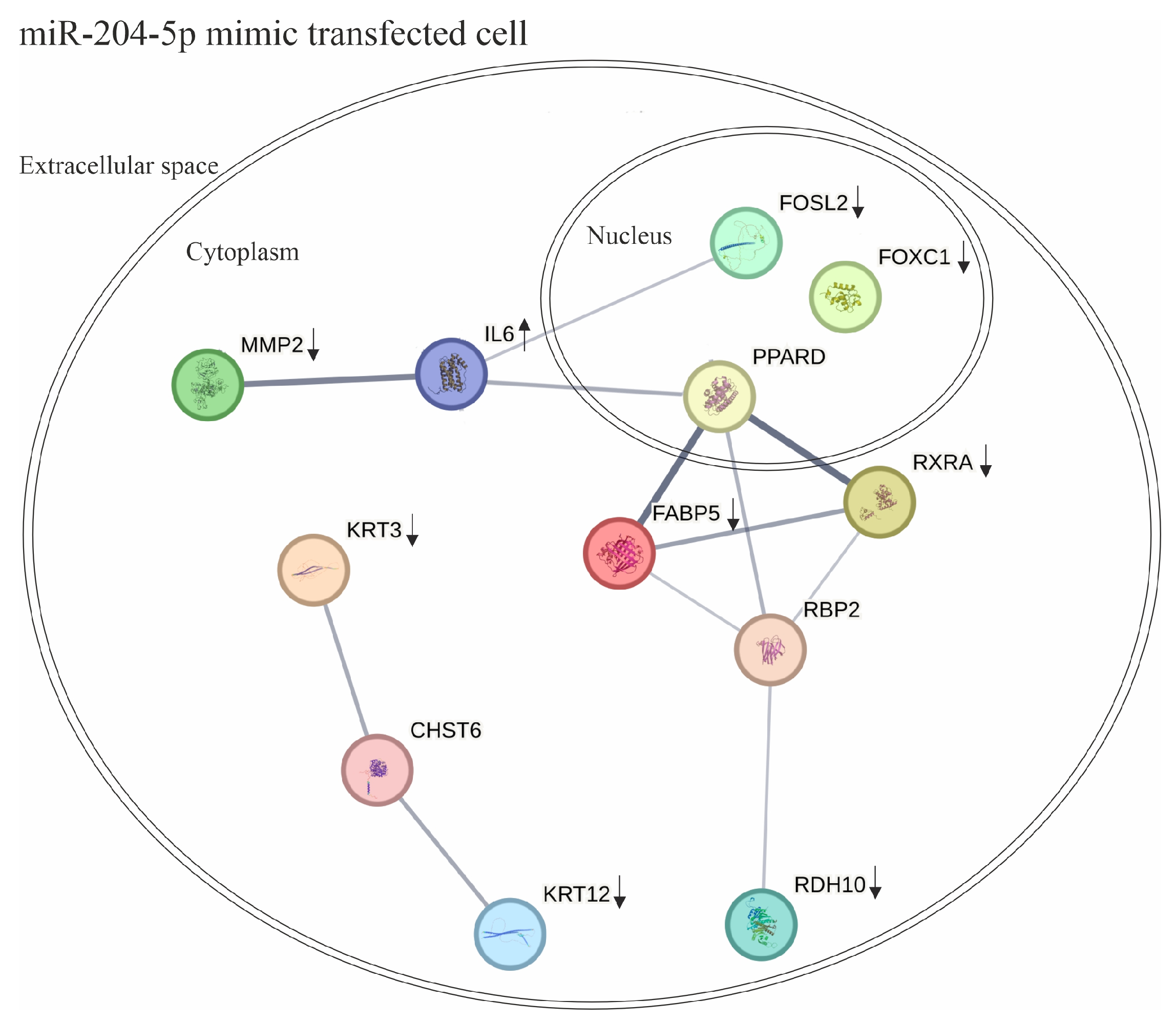
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of the Primary Limbal Epithelial Cells
4.2. Transfection and LPS Treatment of the Primary LECs
4.3. The BrdU Assay
4.4. Protein, RNA, and cDNA Extraction
4.5. Quantitative PCR (qPCR)
4.6. The Western Blot Analysis
4.7. The Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3’UTRs | 3’ untranslated regions |
| ABCG2 | ATP-binding cassette transporter subfamily G member 2 |
| ADH1A1 | Alcohol Dehydrogenase 1A1 |
| ADH7 | Alcohol Dehydrogenase 7 |
| ANGPT1 | Angiopoietin 1 |
| BSA | bovine serum albumin |
| CRABP2 | Cellular Retinoic-Acid-Binding Protein 2 |
| Ctrl | negative control |
| CYP1B1 | Cytochrome P450 Family 1 Subfamily B Member 1 |
| CYP26A1 | Cytochrome P450 26A1 |
| ECL | enhanced chemiluminescence |
| ELISA | enzyme-linked immunosorbent assay |
| FABP5 | Fatty-Acid-Binding Protein 5 |
| FOSL2 | Fos-related antigen 2 |
| FOXC1 | Forkhead Box C1 |
| IL-1β | interleukin-1β |
| IL-6 | interleukin-6 |
| KRT12 | keratin 12 |
| KRT3 | keratin 3 |
| LECs | limbal epithelial cells |
| LPS | lipopolysaccharide |
| Meis2 | Meis homeobox 2 |
| mg | milligrams |
| miR-204-5p | MicroRNA-204-5p |
| MMP2 | Matrix Metalloproteinases 2 |
| MMP9 | Matrix Metalloproteinases 9 |
| OD | optical density |
| pg | picograms |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| PTGES2 | Prostaglandin E Synthase 2 |
| qPCR | quantitative PCR |
| RARA | Retinoic Acid Receptor α |
| RARB | Retinoic Acid Receptor β |
| RBP1 | Retinol-Binding Protein 1 |
| RDH10 | Retinol Dehydrogenase 10 |
| RXRA | Retinoid X Receptor α |
| RXRB | Retinoid X Receptor β |
| TRPM3 | Transient Receptor Potential Melastatin 3 |
| VEGFα | Vascular Endothelial Growth Factor α |
| μL | microliters |
References
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef]
- Yang, F.; Bian, Z.; Xu, P.; Sun, S.; Huang, Z. MicroRNA-204-5p: A pivotal tumor suppressor. Cancer Med. 2022, 12, 3185–3200. [Google Scholar] [CrossRef] [PubMed]
- Conte, I.; Carrella, S.; Avellino, R.; Karali, M.; Marco-Ferreres, R.; Bovolenta, P.; Banfi, S. miR-204 is required for lens and retinal development via Meis2 targeting. Proc. Natl. Acad. Sci. USA 2010, 107, 15491–15496. [Google Scholar] [CrossRef] [PubMed]
- Shaham, O.; Gueta, K.; Mor, E.; Oren-Giladi, P.; Grinberg, D.; Xie, Q.; Cvekl, A.; Shomron, N.; Davis, N.; Keydar-Prizant, M.; et al. Pax6 Regulates Gene Expression in the Vertebrate Lens through miR-204. PLoS Genet. 2013, 9, e1003357. [Google Scholar] [CrossRef]
- Zhai, M.; Zhu, Y.; Yang, M.; Mao, C. Human Mesenchymal Stem Cell Derived Exosomes Enhance Cell-Free Bone Regeneration by Altering Their miRNAs Profiles. Adv. Sci. 2020, 7, 2001334. [Google Scholar] [CrossRef] [PubMed]
- Divoux, A.; Xie, H.; Li, J.; Karastergiou, K.; Perera, R.J.; Chang, R.J.; Fried, S.K.; Smith, S.R. MicroRNA-196 regulates HOX Genes Expression in Human Gluteal Adipose Tissue. Obes. Silver Spring Md. 2017, 25, 1375–1383. [Google Scholar] [CrossRef]
- An, J.; Chen, X.; Chen, W.; Liang, R.; Reinach, P.S.; Yan, D.; Tu, L. MicroRNA Expression Profile and the Role of miR-204 in Corneal Wound Healing. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3673–3683. [Google Scholar] [CrossRef]
- Lu, Y.; Tai, P.W.; Ai, J.; Gessler, D.J.; Su, Q.; Yao, X.; Zheng, Q.; Zamore, P.D.; Xu, X.; Gao, G. Transcriptome Profiling of Neovascularized Corneas Reveals miR-204 as a Multi-target Biotherapy Deliverable by rAAVs. Mol. Ther. Nucleic Acids 2018, 10, 349–360. [Google Scholar] [CrossRef]
- Abbasi, M.; Amini, M.; Moustardas, P.; Gutsmiedl, Q.; Javidjam, D.; Suiwal, S.; Seitz, B.; Fries, F.N.; Dashti, A.; Rautavaara, Y.; et al. Effects of miR-204-5p modulation on PAX6 regulation and corneal inflammation. Sci. Rep. 2024, 14, 26436. [Google Scholar] [CrossRef]
- Hingorani, M.; Hanson, I.; van Heyningen, V. Aniridia. Eur. J. Hum. Genet. EJHG 2012, 20, 1011–1017. [Google Scholar] [CrossRef]
- Latta, L.; Ludwig, N.; Krammes, L.; Stachon, T.; Fries, F.; Mukwaya, A.; Szentmáry, N.; Seitz, B.; Wowra, B.; Kahraman, M.; et al. Abnormal neovascular and proliferative conjunctival phenotype in limbal stem cell deficiency is associated with altered microRNA and gene expression modulated by PAX6 mutational status in congenital aniridia. Ocul. Surf. 2021, 19, 115–127. Available online: https://pubmed.ncbi.nlm.nih.gov/32422284/ (accessed on 12 January 2025). [CrossRef] [PubMed]
- Latta, L.; Nordström, K.; Stachon, T.; Langenbucher, A.; Fries, F.N.; Szentmáry, N.; Seitz, B.; Käsmann-Kellner, B. Expression of retinoic acid signaling components ADH7 and ALDH1A1 is reduced in aniridia limbal epithelial cells and a siRNA primary cell based aniridia model. Exp. Eye Res. 2019, 179, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Girardi, C.S.; Rostirolla, D.C.; Lini, F.J.M.; Brum, P.O.; Delgado, J.; Ribeiro, C.T.; Teixeira, A.A.; Peixoto, D.O.; Heimfarth, L.; Kunzler, A.; et al. Nuclear RXRα and RXRβ receptors exert distinct and opposite effects on RA-mediated neuroblastoma differentiation. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2019, 1866, 317–328. [Google Scholar] [CrossRef]
- Janesick, A.; Wu, S.C.; Blumberg, B. Retinoic acid signaling and neuronal differentiation. Cell. Mol. Life Sci. 2015, 72, 1559–1576. Available online: https://link.springer.com/article/10.1007/S00018-014-1815-9 (accessed on 12 January 2025). [CrossRef]
- Latta, L.; Viestenz, A.; Stachon, T.; Colanesi, S.; Szentmáry, N.; Seitz, B.; Käsmann-Kellner, B. Human aniridia limbal epithelial cells lack expression of keratins K3 and K12. Exp. Eye Res. 2018, 167, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Szentmáry, N.; Berger, T.; Seitz, B.; Fries, F.N.; Suiwal, S.; Amini, M.; Stachon, T. Reciprocal effects of conditioned medium on gene and protein expression of limbal epithelial cells and limbal fibroblasts in congenital aniridia. DOG Poster, 2024.
- Katiyar, P.; Stachon, T.; Fries, F.N.; Parow, F.; Ulrich, M.; Langenbucher, A.; Cayless, A.; Seitz, B.; Käsmann-Kellner, B.; Latta, L.; et al. Decreased FABP5 and DSG1 protein expression following PAX6 knockdown of differentiated human limbal epithelial cells. Exp. Eye Res. 2022, 215, 108904. [Google Scholar] [CrossRef]
- Chung, T.; Lau, T.; Yim, S.; Lo, K.; Siu, N.; Chan, L.; Yu, M.; Kwong, J.; Doran, G.; Barroilhet, L.; et al. Dysregulation of microRNA-204 mediates migration and invasion of endometrial cancer by regulating FOXC1. Int. J. Cancer 2012, 130, 1036–1045. [Google Scholar] [CrossRef]
- Gao, W.; Wu, Y.; He, X.; Zhang, C.; Zhu, M.; Chen, B.; Liu, Q.; Qu, X.; Li, W.; Wen, S.; et al. MicroRNA-204-5p inhibits invasion and metastasis of laryngeal squamous cell carcinoma by suppressing forkhead box C1. J. Cancer 2017, 8, 2356–2368. [Google Scholar] [CrossRef]
- Birnhuber, A.; Biasin, V.; Schnoegl, D.; Marsh, L.M.; Kwapiszewska, G. Transcription factor Fra-2 and its emerging role in matrix deposition, proliferation and inflammation in chronic lung diseases. Cell Signal 2019, 64, 109408. [Google Scholar] [CrossRef]
- Smits, J.G.A.; Cunha, D.L.; Amini, M.; Bertolin, M.; Laberthonnière, C.; Qu, J.; Owen, N.; Latta, L.; Seitz, B.; Roux, L.N.; et al. Identification of the regulatory circuit governing corneal epithelial fate determination and disease. PLoS Biol. 2023, 21, e3002336. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yin, S.; Chen, Y.; Wu, Y.; Zheng, W.; Dong, H.; Bai, Y.; Qin, Y.; Li, J.; Feng, S.; et al. LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Mol. Med. Rep. 2018, 17, 5484–5491. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Liu, X.; Cheng, Q. MicroRNA-204-5p suppresses IL6-mediated inflammatory response and chemokine generation in HK-2 renal tubular epithelial cells by targeting IL6R. Biochem. Cell Biol. Biochim. Biol. Cell 2019, 97, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-T.; Wang, C.-Y.; Ho, C.-H.; Tzeng, S.-F. Interleukin-6 Inhibits Expression of miR-204-5p, a Regulator of Oligodendrocyte Differentiation: Involvement of miR-204-5p in the Prevention of Chemical-Induced Oligodendrocyte Impairment. Mol. Neurobiol. 2024, 61, 1953–1968. [Google Scholar] [CrossRef]
- Webb, A.H.; Gao, B.T.; Goldsmith, Z.K.; Irvine, A.S.; Saleh, N.; Lee, R.P.; Lendermon, J.B.; Bheemreddy, R.; Zhang, Q.; Brennan, R.C.; et al. Inhibition of MMP-2 and MMP-9 decreases cellular migration, and angiogenesis in in vitro models of retinoblastoma. BMC Cancer 2017, 17, 434. [Google Scholar] [CrossRef]
- Cunningham, T.J.; Duester, G. Mechanisms of retinoic acid signalling and its roles in organ and limb development. Nat. Rev. Mol. Cell Biol. 2015, 16, 110–123. [Google Scholar] [CrossRef]
- Sandell, L.L.; Sanderson, B.W.; Moiseyev, G.; Johnson, T.; Mushegian, A.; Young, K.; Rey, J.-P.; Ma, J.-X.; Staehling-Hampton, K.; Trainor, P.A. RDH10 is essential for synthesis of embryonic retinoic acid and is required for limb, craniofacial, and organ development. Genes Dev. 2007, 21, 1113. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor Nr. | Age (Years) | Gender |
|---|---|---|
| Donor 1 | 85 | Female |
| Donor 2 | n/a | Male |
| Donor 3 | 73 | Female |
| Donor 4 | n/a | Male |
| Donor 5 | n/a | Male |
| Donor 6 | n/a | Female |
| Donor 7 | n/a | Male |
| Primer | Gene Globe ID | Manufacturer |
|---|---|---|
| Hs_RARA_1_SG QuantiTect Primer Assay | QT00095865 | Qiagen N.V., Venlo, The Netherlands |
| Hs_RARB_1_SG QuantiTect Primer Assay | QT00062741 | Qiagen N.V., Venlo, The Netherlands |
| Hs_RXRA_1_SG QuantiTect Primer Assay | QT00005726 | Qiagen N.V., Venlo, The Netherlands |
| Hs_RXRB_1_SG QuantiTect Primer Assay | QT00061117 | Qiagen N.V., Venlo, The Netherlands |
| Hs_CRABP2_1_SG QuantiTect Primer Assay | QT00063434 | Qiagen N.V., Venlo, The Netherlands |
| Hs_RBP1_2_SG QuantiTect Primer Assay | QT01850296 | Qiagen N.V., Venlo, The Netherlands |
| Hs_RDH10_1_SG QuantiTect Primer Assay | QT00029176 | Qiagen N.V., Venlo, The Netherlands |
| Hs_ADH7_1_SG QuantiTect Primer Assay | QT00000217 | Qiagen N.V., Venlo, The Netherlands |
| Hs_ALDH1A1_1_SG QuantiTect Primer Assay | QT00013286 | Qiagen N.V., Venlo, The Netherlands |
| Hs_FABP5_1_SG QuantiTect Primer Assay | QT0022556 | Qiagen N.V., Venlo, The Netherlands |
| Hs_CYP1B1_1_SG QuantiTect Primer Assay | QT00209496 | Qiagen N.V., Venlo, The Netherlands |
| Hs_CYP26A1_1_SG QuantiTect Primer Assay | QT00026817 | Qiagen N.V., Venlo, The Netherlands |
| Hs_ABCG2_1_SG QuantiTect Primer Assay | QT00073206 | Qiagen N.V., Venlo, The Netherlands |
| Hs_PTGES2_1_SG QuantiTect Primer Assay | QT00082068 | Qiagen N.V., Venlo, The Netherlands |
| Hs_IL1B_1_SG QuantiTect Primer Assay | QT00021385 | Qiagen N.V., Venlo, The Netherlands |
| Hs_IL16_1_SG QuantiTect Primer Assay | QT00075138 | Qiagen N.V., Venlo, The Netherlands |
| Hs_FOSL2_1_SG QuantiTect Primer Assay | QT01000881 | Qiagen N.V., Venlo, The Netherlands |
| Hs_PPARG_1_SG QuantiTect Primer Assay | QT00029841 | Qiagen N.V., Venlo, The Netherlands |
| Hs_FOXC1_1_SG QuantiTect Primer Assay | QT00217161 | Qiagen N.V., Venlo, The Netherlands |
| Hs_MEIS2_1_SG QuantiTect Primer Assay | QT00077315 | Qiagen N.V., Venlo, The Netherlands |
| Hs_KRT3_1_SG QuantiTect Primer Assay | QT00050365 | Qiagen N.V., Venlo, The Netherlands |
| Hs_KRT12_1_SG QuantiTect Primer Assay | QT00011949 | Qiagen N.V., Venlo, The Netherlands |
| Hs_MMP2_vb.1_SG QuantiTect Primer Assay | QT02395778 | Qiagen N.V., Venlo, The Netherlands |
| Hs_MMP9_1_SG QuantiTect Primer Assay | QT00040040 | Qiagen N.V., Venlo, The Netherlands |
| Hs_TBP_1_SG QuantiTect Primer Assay | QT00000721 | Qiagen N.V., Venlo, The Netherlands |
| Antibody | Class | Dilution | Cat. No. | Manufacturer |
|---|---|---|---|---|
| RXRa | Polyclonal | 1:2000 | 21218-1-AP | Proteintech, Planegg-Martinsried, Germany |
| RDH10 | Polyclonal | 1:1000 | 14644-1-AP | Proteintech, Planegg-Martinsried, Germany |
| FABP5 | Polyclonal | 1:1000 | 12348-1-AP | Proteintech, Planegg-Martinsried, Germany |
| FOSL2 | Monoclonal | 1:3000 | TA809660S | OriGene Technologies, Rockville, MD, USA |
| FOXC1 | Polyclonal | 1:500 | 55365-1-AP | Proteintech, Planegg-Martinsried, Germany |
| Keratin K3/K76 | Monoclonal | 1:500 | CBL218 | MERCK, Darmstadt, Germany |
| Keratin 12 (E-8) | Monoclonal | 1:2000 | sc-515882 | Santa Cruz biotechnology, Dallas, TX, USA |
| MMP2 | Monoclonal | 1:250 | 66366-1-Ig | Proteintech, Planegg-Martinsried, Germany |
| β-actin | Polyclonal | 1:10,000 | Ab8227 | Abcam, Cambridge, UK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amini, M.; Stachon, T.; Hsu, S.-L.; Li, Z.; Chai, N.; Fries, F.N.; Seitz, B.; Kundu, S.; Suiwal, S.; Szentmáry, N. Effect of MiRNA 204-5P Mimics and Lipopolysaccharide-Induced Inflammation on Transcription Factor Levels, Cell Maintenance, and Retinoic Acid Signaling in Primary Limbal Epithelial Cells. Int. J. Mol. Sci. 2025, 26, 3809. https://doi.org/10.3390/ijms26083809
Amini M, Stachon T, Hsu S-L, Li Z, Chai N, Fries FN, Seitz B, Kundu S, Suiwal S, Szentmáry N. Effect of MiRNA 204-5P Mimics and Lipopolysaccharide-Induced Inflammation on Transcription Factor Levels, Cell Maintenance, and Retinoic Acid Signaling in Primary Limbal Epithelial Cells. International Journal of Molecular Sciences. 2025; 26(8):3809. https://doi.org/10.3390/ijms26083809
Chicago/Turabian StyleAmini, Maryam, Tanja Stachon, Shao-Lun Hsu, Zhen Li, Ning Chai, Fabian N. Fries, Berthold Seitz, Swarnali Kundu, Shweta Suiwal, and Nóra Szentmáry. 2025. "Effect of MiRNA 204-5P Mimics and Lipopolysaccharide-Induced Inflammation on Transcription Factor Levels, Cell Maintenance, and Retinoic Acid Signaling in Primary Limbal Epithelial Cells" International Journal of Molecular Sciences 26, no. 8: 3809. https://doi.org/10.3390/ijms26083809
APA StyleAmini, M., Stachon, T., Hsu, S.-L., Li, Z., Chai, N., Fries, F. N., Seitz, B., Kundu, S., Suiwal, S., & Szentmáry, N. (2025). Effect of MiRNA 204-5P Mimics and Lipopolysaccharide-Induced Inflammation on Transcription Factor Levels, Cell Maintenance, and Retinoic Acid Signaling in Primary Limbal Epithelial Cells. International Journal of Molecular Sciences, 26(8), 3809. https://doi.org/10.3390/ijms26083809