Expression of VDAC Regulated by Extracts of Limonium sinense Ktze root Against CCl4-induced Liver Damage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant material
2.2. Chemicals
2.3. Animals
2.4. Carbon tetrachloride (CCl4)-induced hepatotoxicity in mice
2.5 Aminotransferase activity determination
2.6 Isolation of liver mitochondria
2.7. Measurement of mitochondrial membrane potential
2.8 Evaluation of VDAC mRNA level by RT-PCR assay
2.9 Western Blot analysis for VDAC
2.10 Statistical analysis
3. Results
3.1 Inhibition in the elevation of sALT and sAST level induced by CCl4
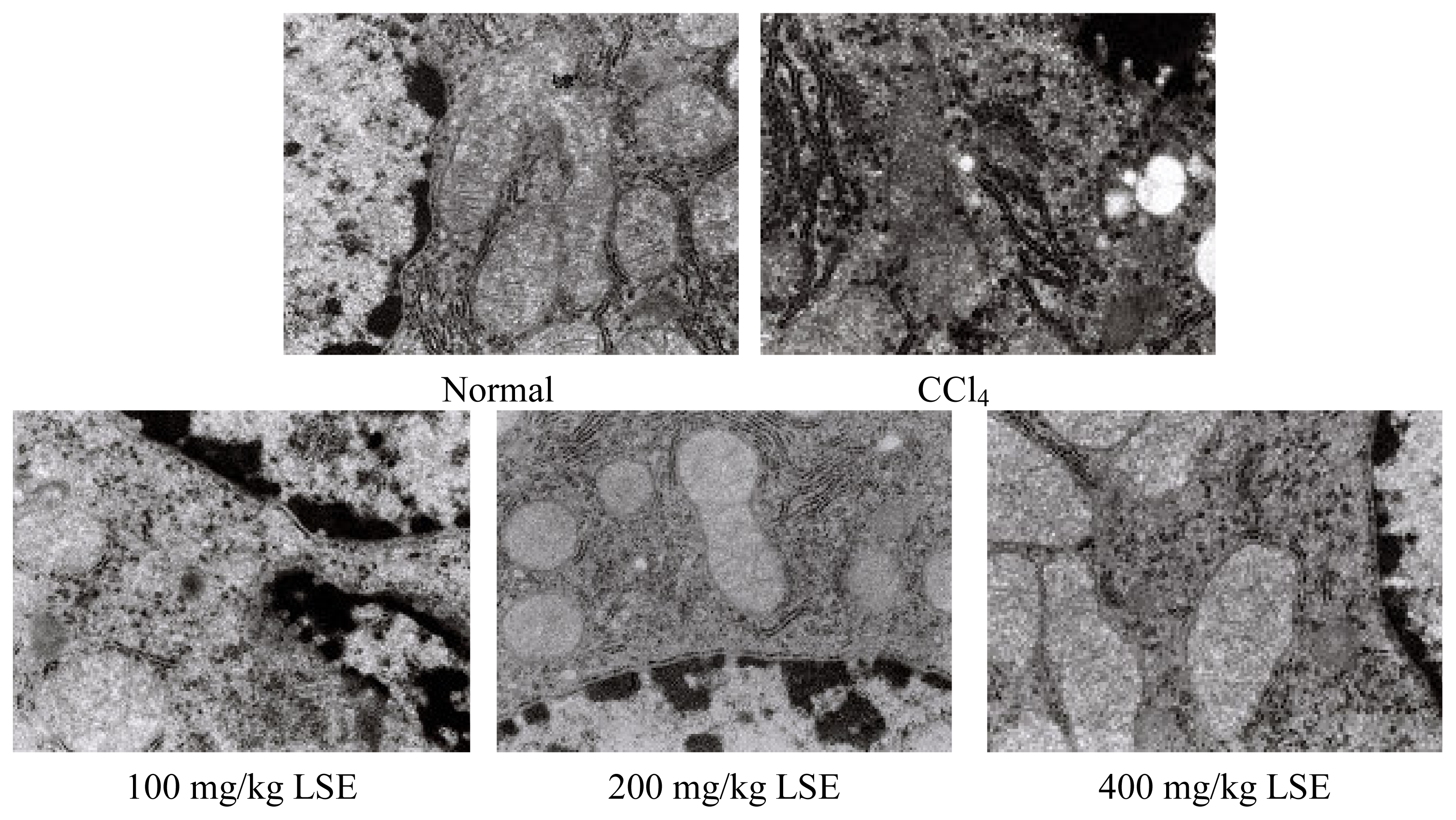
3.2 Protection on the ultrastructure of liver insulted by CCl4
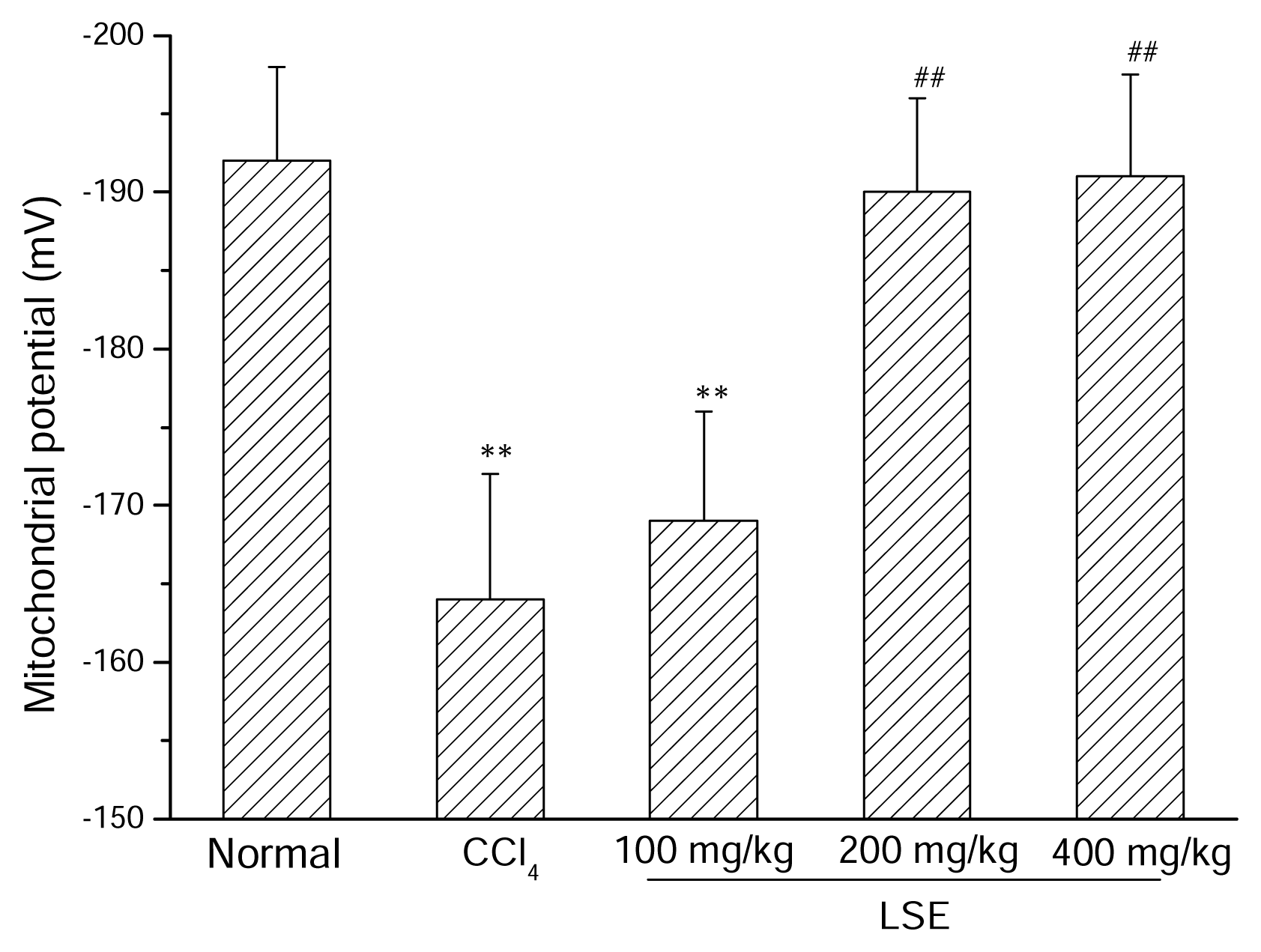
3.3. Effect of LSE on mitochondrial membrane potential dissipation
3.4 Effect of LSE on mitochondrial VDAC expression in CCl4-insulted mouse livers
Down-regulation on liver VDAC mRNA level in CCl4-stimulated mice
Down-regulation on liver VDAC protein level in CCl4-stimulated mice
4. Discussion




Acknowledgements
References and Notes
- Kaplowitz, N. Mechanisms of liver cell injury. J. Hepatol 2000, 32 (1 Suppl), 39–47. [Google Scholar]
- Newmeyer, D.D.; Ferguson-Miller, S. Mitochondria: releasing power for live and unleashing the machineries of death. Cell 2003, 112, 481–490. [Google Scholar]
- Orrenius, S. Mitochondrial regulation of apoptotic cell death. Toxicol. Lett 2004, 149, 19–23. [Google Scholar]
- Wallace, D.C. Mitochondrial diseases in man and mouse. Science 1999, 283(5407), 1482–1488. [Google Scholar]
- Tang, X.H.; Gao, J.; Fang, F.; et al. Hepatoprotection of oleanolic acid is related to its inhibition on mitochondrial permeability transition. Am. J. Chin. Med 2005, 33(4), 627–637. [Google Scholar]
- Tang, X.H.; Gao, J.; Wang, Y.P.; et al. Effective protection of Terminalia catappa L. leaves from damage-induced by carbon tetrachloride in liver mitochondria. J. Nutr. Biochem 2006, 17(3), 177–182. [Google Scholar]
- Godbole, A.; Varghese, J.; Sarin, A.; et al. VDAC is a conserved element of death pathways in plant and animal systems. Biochim. et Biophys. Acta 2003, 1642, 87–96. [Google Scholar]
- Gao, J.; Chen, J.; Tang, X.H.; et al. Mechanisms underlying mitochondrial protection of Asiatic acid against hepatotoxicity in mice. J. Pharm and Pharmacol. 2006, 58, 227–233. [Google Scholar]
- Li, H.L. Plumbaginaceae, Flora of Taiwan. In Editorial Committee of the Flora of Taiwan; Volume IV, Edited, & Published, Taipei; 1978; pp. 90–93. [Google Scholar]
- Chaung, S.S.; Lin, C.C.; Lin, J.; et al. The hepatoprotective effects of Limonium sinenseagainst carbon tetrachloride and beta-D-galactosamine intoxication in rats. Phytother Res 2003, 17(7), 784–791. [Google Scholar]
- Lin, L.C.; Chou, C.J. Flavonoids and phenolics from Limonium sinense. Planta Med 2000, 66, 382–383. [Google Scholar]
- Aprille, J.R.; Hom, J.A.; Rulfs, J. Liver and skeletal muscle mitochondrial function following burn injury. J. Trauma 1977, 17, 279–287. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Emaus, R.K.; Grunwald, R.; Lemaster, J.J. Rhodamine 123 as a probe of transmembrane potential in isolated rat liver mitochondria. Biochim. Biophy. Act 1986, 850, 436–448. [Google Scholar]
- Elimadi, A.; Sapena, R.; Settaf, A.; et al. Attenuation of liver normothermic ischemia--reperfusion injury by preservation of mitochondrial functions with S-15176, a potent trimetazidine derivative. Biochem. Pharmacol 2001, 62(4), 509–516. [Google Scholar]
- Hewawasam, R.P.; Jayatilaka, K.A.; Pathirana, C.; et al. Protective effect of Asteracantha longifolia extract in mouse liver injury induced by carbon tetrachloride and paracetamol. J. Pharm. Pharmacol 2003, 55, 1413–1418. [Google Scholar]
- Itokazu, Y.; Segawa, Y.; Omata, T.; et al. Effects of ZNC-2381, a new oral compound, on several hepatic injury models and on hepatocellular apoptosis in mice and rats. J. Pharm. Pharmacol 2000, 52, 531–538. [Google Scholar]
- Daba, M.H.; Abdel-Rahman, M.S. Hepatoprotective activity of thymoquinone in isolated rat hepatocytes. Toxicol. Lett 1998, 95, 23–29. [Google Scholar]
- Hernandez-Munoz, R.; Diaz-Munoz, M.; Sanchez, V.C. Effects of adenosine administration on the function and membrane composition of liver mitochondria in carbon tetrachloride-induced cirrhosis. Arch. Biochem. Biophys 1992, 294, 160–167. [Google Scholar]
- Premkumar, A.; Simantov, R. Mitochondrial voltage-dependent anion channel is involved in dopamine-induced apoptosis. J. Neurochem 2002, 82(2), 345–352. [Google Scholar]
- Massa, R.; Marliera, L.N.; Martorana, A. Intracellular localization and isoform expression of the voltage-dependent anion channel (VDAC) in normal and dystrophic skeletal muscle. J. Muscle Res. Cell Motil. 2000, 21(5), 433–442. [Google Scholar]
- Shinohara, Y.; Ishida, T.; Hino, M. Characterization of porin isoforms expressed in tumor cells. Eur. J. Biochem 2000, 267(19), 6067–6073. [Google Scholar]
© 2007 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Tang, X.; Ga, J.; Chen, J.; Xu, L.; Tang, Y.; Dou, H.; Yu, W.; Zhao, X. Expression of VDAC Regulated by Extracts of Limonium sinense Ktze root Against CCl4-induced Liver Damage. Int. J. Mol. Sci. 2007, 8, 204-213. https://doi.org/10.3390/i8030204
Tang X, Ga J, Chen J, Xu L, Tang Y, Dou H, Yu W, Zhao X. Expression of VDAC Regulated by Extracts of Limonium sinense Ktze root Against CCl4-induced Liver Damage. International Journal of Molecular Sciences. 2007; 8(3):204-213. https://doi.org/10.3390/i8030204
Chicago/Turabian StyleTang, Xinhui, Jing Ga, Jin Chen, Lizhi Xu, Yahong Tang, Huan Dou, Wen Yu, and Xiaoning Zhao. 2007. "Expression of VDAC Regulated by Extracts of Limonium sinense Ktze root Against CCl4-induced Liver Damage" International Journal of Molecular Sciences 8, no. 3: 204-213. https://doi.org/10.3390/i8030204
APA StyleTang, X., Ga, J., Chen, J., Xu, L., Tang, Y., Dou, H., Yu, W., & Zhao, X. (2007). Expression of VDAC Regulated by Extracts of Limonium sinense Ktze root Against CCl4-induced Liver Damage. International Journal of Molecular Sciences, 8(3), 204-213. https://doi.org/10.3390/i8030204