Effect of Dietary Protein Level and Origin on the Redox Status in the Digestive Tract of Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1 Animals and diets
2.2 Sampling procedures
2.3 Analytical methods
2.3.1 Superoxide anion determination
2.3.2 Lipid peroxidation determination
2.3.3 Glutathione peroxidase assay
2.3.4 Superoxide dismutase assay
2.3.5 GSH assay
2.3.6 Na+ K+-ATPaseand catalase activity determination
2.4 Statistical analysis
3. Results
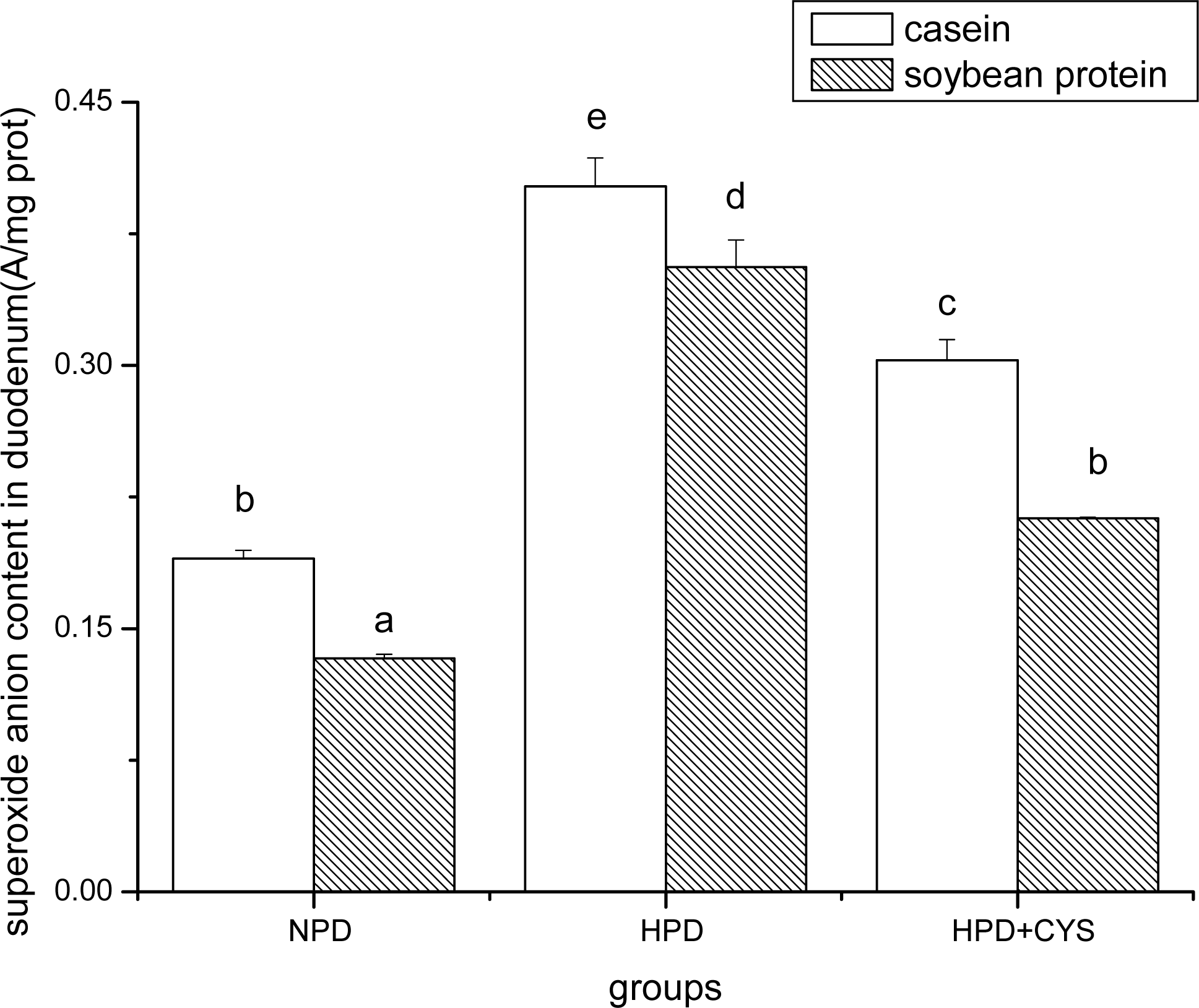
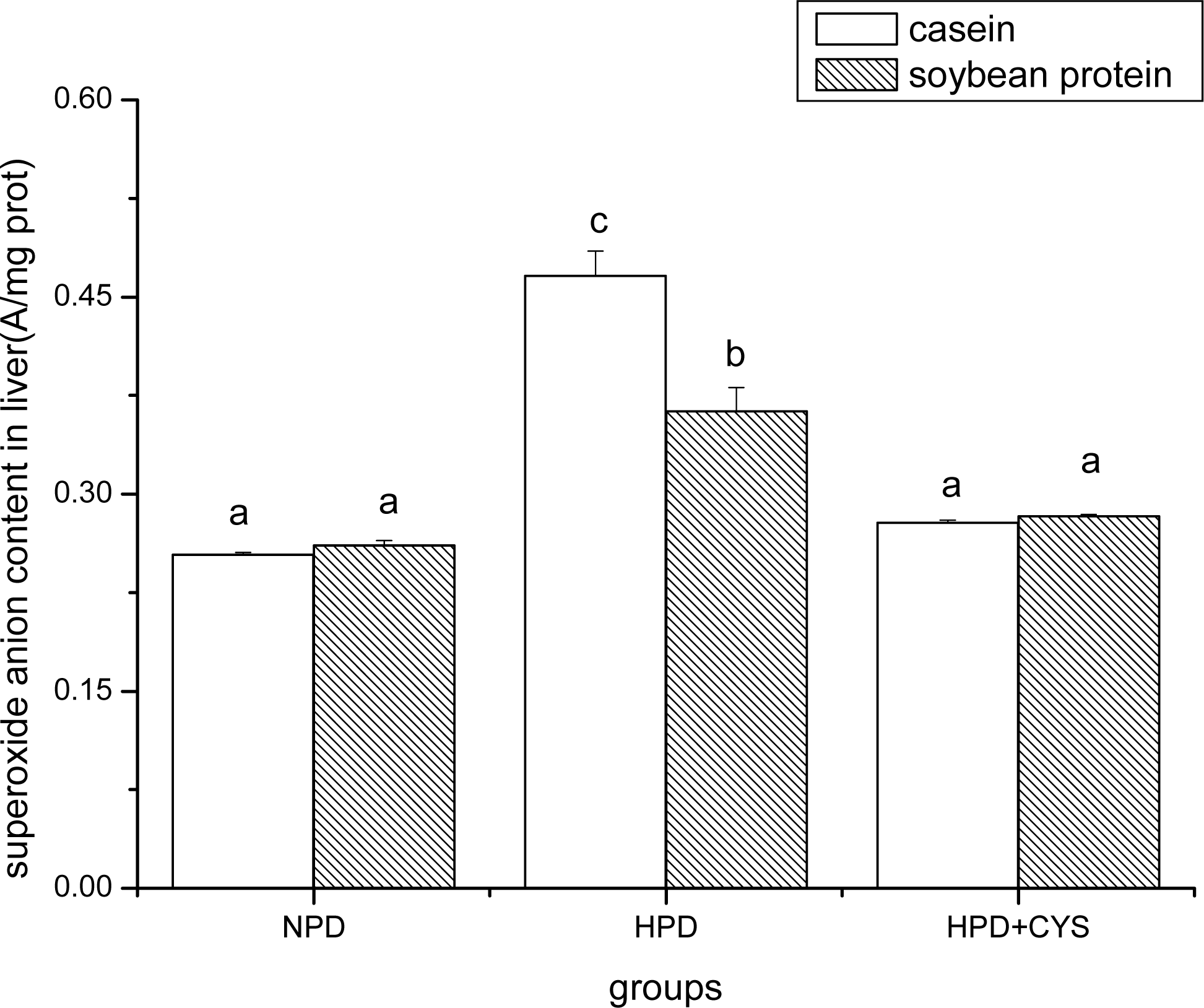
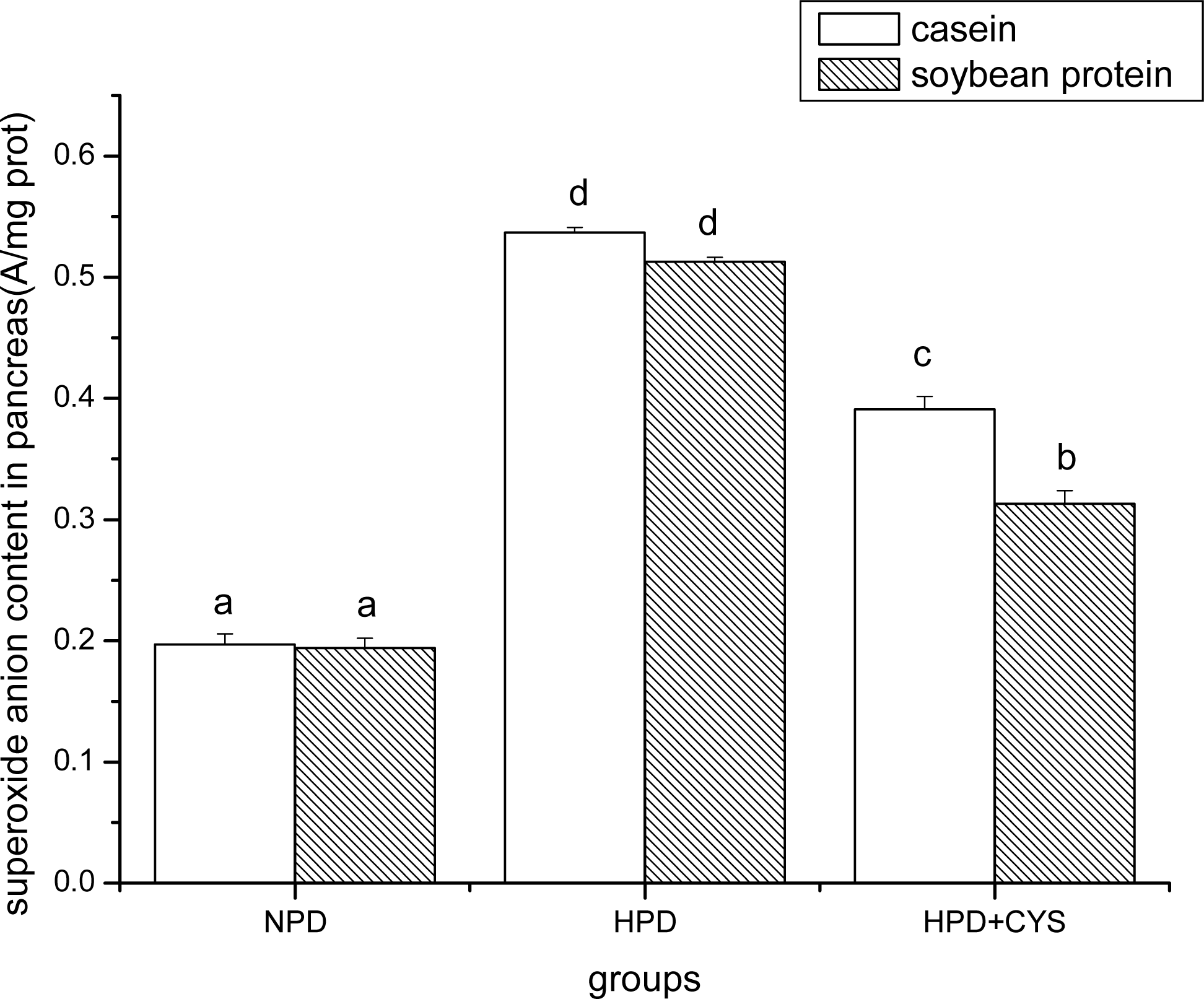
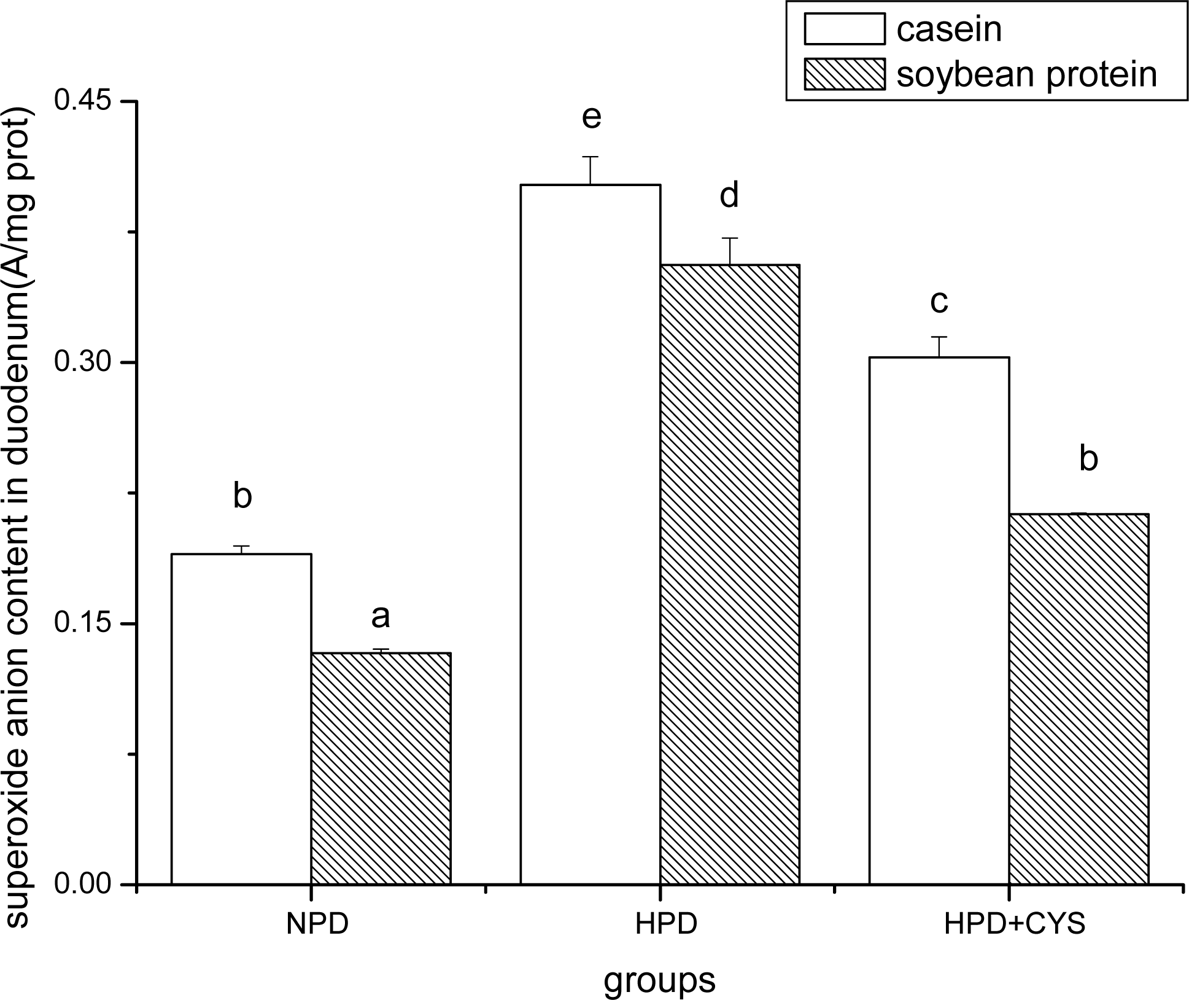
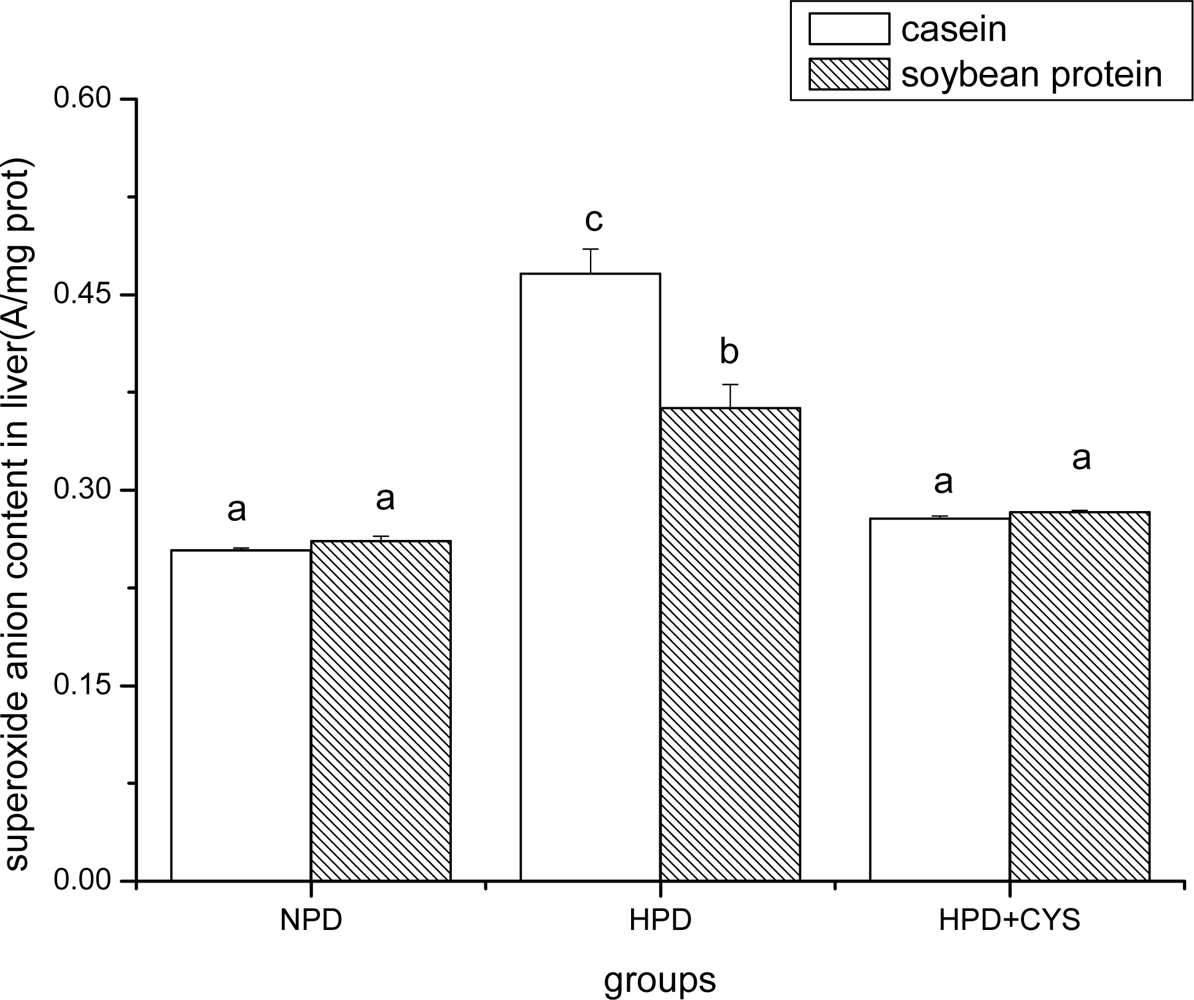
3.1 Superoxide anion content in duodenum, liver and pancreas of mice
3.2 MDA contents and SOD activities in duodenum, liver and pancreas of mice
3.3 GSH-Px activities and GSH contents in duodenum, liver and pancreas of mice
3.4 CAT and Na+ K+-ATPase activities in duodenum, liver and pancreas of mice
4. Discussion
Acknowledgments
References
- Bankir, L; Kriz, W. Adaptation of the kidney to protein intake and to urine concentrating activity: similar consequences in health and CRF. Kidney Int 1995, 47, 7–24. [Google Scholar]
- Brändle, E; Sieberth, HG; Hautmann, RE. Effect of chronic dietary protein intake on the renal function in healthy subjects. Eur J Clin Nutr 1996, 50, 734–740. [Google Scholar]
- Yanagisawa, H; Wada, O. Effects of dietary protein on eicosanoid production in rat renal tubules. Nephron 1998, 78, 179–186. [Google Scholar]
- Chow, WH; Gridley, G; McLaughlin, JK; Mandel, JS; Wacholder, S; Blot, WJ; Niwa, S; Fraumeni, JF, Jr. Protein intake and risk of renal cell cancer. J Natl Cancer Inst 1994, 86, 1131–1139. [Google Scholar]
- Kitagawa, T; Owada, M; Urakami, T; Yamauchi, K. Increased incidence of non-insulin dependent diabetes mellitus among Japanese schoolchildren correlates with an increased intake of animal protein and fat. Clin Pediatr 1998, 37, 111–115. [Google Scholar]
- Tsunehara, CH; Leonetti, DL; Fujimoto, WY. Diet of second-generation Japanese-American mean with and without non-insulin-dependent diabetes. Am J Clin Nutr 1990, 52, 731–738. [Google Scholar]
- Vlajinac, HD; Marinkovic, JM; Ilic, MD; Kocev, NI. Diet and prostate cancer: a case-control study. Eur J Cancer 1997, 33, 101–107. [Google Scholar]
- Holmes, RP; Goodman, HO; Hart, LJ; Assimos, DG. Relationship of protein intake to urinary oxalate and glycolate excretion. Kidney Int 1993, 44, 366–372. [Google Scholar]
- Price, GM; Halliday, D; Pacy, PJ; Quevedo, MR; Millward, DJ. Nitrogen homeostasis in man: Influence of protein intake on the amplitude of diurnal cycling of body nitrogen. Clin Sci 1994, 86, 91–102. [Google Scholar]
- Porrata-Maury, C; Aust, L; Noack, R; Eschrich, H. Studies on the postprandial thermogenic action of proteins and protein mixtures in rats. Nahrung 1987, 31, 311–319. [Google Scholar]
- Robinson, SM; Jaccard, C; Persaud, C; Jackson, AA; Jequier, E; Schutz, Y. Protein turnover and thermogenesis in response to high-protein and high-carbohydrate feeding in men. Am J Clin Nutr 1990, 52, 72–80. [Google Scholar]
- Petzke, KJ; Proll, J. [NAD1] [NADH] redox-state metabolites of freezed-clamped livers of rats fed casein or gelatin diets. J Nutr Biochem 1994, 5, 495–498. [Google Scholar]
- Park, MS; Kudchodkar, BJ; Liepa, GU. Effects of dietary animal and plant proteins on the cholesterol metabolism in immature and mature rats. J Nutr 1987, 117, 30–35. [Google Scholar]
- Messina, M. Soy intake and cancer risk: a review of the in vitro and in vivo data. Nutr Cancer 1994, 21, 113–131. [Google Scholar]
- Madani, S; Lopez, S; Blond, JP; Prost, J; Belleville, J. Highly purified soybean protein is not hypocholesterolemic in rats but stimulates cholesterol synthesis and excretion and reduces polyunsaturated fatty acid biosynthesis. J Nutr 1998, 128, 1084–1091. [Google Scholar]
- Choi, YS; Goto, S; Ikeda, I; Sugano, M. Interaction of dietary protein, cholesterol and age on lipid metabolism of the rat. Br J Nutr 1989, 61, 531–543. [Google Scholar]
- Madani, S; Prost, J; Belleville, J. Dietary protein level and origin (casein and highly purified soybean protein) affect hepatic storage, plasma lipid transport, and antioxidative defense status in the rat. Nutrition 2000, 16, 368–375. [Google Scholar]
- Kanazawa, T; Osanak, T; Zhang, XS; Umehara, T; Yin, XZ; Onodera, K; Oike, Y; Ohkubo, K. Protective effects of soy protein on the peroxidizability of lipoprotein in cerebrovascular disease. J Nutr 1995, 125, 639S–646S. [Google Scholar]
- Aoki, H; Otaka, Y; Igarashi, K; Takenaka, A. Soy Protein Reduces Paraquat-Induced Oxidative Stress in Rats. J Nutr 2002, 132, 2258–2262. [Google Scholar]
- Pick, E. Microassays for superoxide and hydrogen peroxide production and nitroblue tetrazolium reduction using an enzyme immunoassay microplate reader. Methods Enzymol 1986, 132, 407–421. [Google Scholar]
- Ohkawa, H; Ohishi, N; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biolchem 1979, 95, 351–358. [Google Scholar]
- Hafeman, DG; Sunde, RA; Hoekstra, WG. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J Nutr 1974, 104, 580–587. [Google Scholar]
- Spitz, DR; Oberley, LW. An assay for superoxide dismutase activity in mammalian tissue homogenates. Anal Biochem 1989, 179, 8–18. [Google Scholar]
- Moron, MS; Depierre, JW; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim Biophys Acta 1979, 582, 67–78. [Google Scholar]
- Reinila, M; MacDonald, E; Salem, N, Jr; Linnoila, M; Trams, EG. Standardized method for the detemination of human erythroyte membrane adenosine triphosphatases. Anal Biochem 1982, 124, 19–26. [Google Scholar]
- Aebi, H. Catalase. Bergmeyer, HU, Ed.; In Methods of Enzymatic Analysis, 2nd ed; Verlag Chemie Weinheim: Germany, 1974; pp. 673–684. [Google Scholar]
- Lowry, OH; Rosebrough, HJ; Farr, AL; Randall, RJ. Protein measurement with the Folinphenol reagent. J Biol Chem 1951, 193, 265–275. [Google Scholar]
- Klein, M; Hoffmann, L. The effect of the protein level of food on the energy maintenance requirement of growing rats on different feeding regimens. Arch Tierernahr 1993, 44, 95–109. [Google Scholar]
- Miller, DS; Mumford, P. Gluttony. 1. An experimental study of overeating low-or high-protein diets. Am J Clin Nutr 1967, 20, 1212–1222. [Google Scholar]
- Toyomizu, M; Tanaka, M; Kojima, M; Ishibashi, T. The substrate-specific impairment of oxidative phosphorylation in liver mitochondria from high-protein-fed chickens. Brit J Nutr 1995, 74, 797–806. [Google Scholar]
- Young, IS; McEneny, J. Lipoprotein oxidation and atherosclerosis. Biochem Soc Trans 2001, 29, 358–362. [Google Scholar]
- Zhu, JJ; Le, GW; Shi, YH. Biological function and application of cysteamine. China Feed 2005, 24, 22–23. (in Chinese). [Google Scholar]
- Matos, DG de; Nogueira, D; Cortvrindt, R; Herrera, C; Adriaenssens, T; Pasqualini, RS; Smitz, J. Capacity of adult and prepubertal mouse oocytes to undergo embryo development in the presence of cysteamine. Mol Reprod Dev 2003, 64, 214–218. [Google Scholar]
- Zhao, Sh,M,; He, XJ; Huang, LJ; Jin, L; Wang, CB; Liang, ZR; Huang, YY; Huang, YF. Effect of acute hypoxia on SOD, MDA, NO and Na, K-ATPase in rabbits. J Clin Exp Med 2003, 2, 155–157. (in Chinese). [Google Scholar]
- Glitsch, HG. Electrophysiology of the sodium-potassium-ATPase in cardiac cells. Physiol Rev 2001, 81, 1791–1826. [Google Scholar]
- Jorgensen, PL; Pedersen, PA. Structure-function relationships of Na+, K+, ATP, or Mg2+ binding and energy transduction in Na, K-ATPase. Biochim Biophys Acta 2001, 1505, 57–74. [Google Scholar]
- Guo, JR; Cui, JJ; Ding, JQ; Ren, LY; Yu, LT. Effects of propofol on energy metabolism, Na, K-ATPase activity and lipid peroxidation of hippocampus mitochondrial following global ischemia-reperfusion in rats. Chin J Clin Pharmacol Ther 2004, 9, 893–896. (in Chinese). [Google Scholar]
- He, LY; Huang, CX. Effects of Dall on ATPase and free radicals metabolism of brain following ischemia-reperfusion in rats. J Hubei Preclin Med 2003, 14, 1–3. (in Chinese). [Google Scholar]
- Xie, Z. Molecular mechanisms of Na/K-ATPase-mediated signal transduction. Ann NY Acad Sci 2003, 986, 497–503. [Google Scholar]
- Chiang, MT; Kimura, S. Effect of dietary protein on the peroxidation of eicosapentaenoic acid in stroke-prone spontaneously hypertensive rats. Int J Vitam Nutr Res 1991, 61, 239–243. [Google Scholar]
- Bergeron, N; Jacques, H. Influence of fish protein as compared to casein and soy protein on serum and liver lipids, and serum lipoprotein cholesterol levels in the rabbit. Atherosclerosis 1989, 78, 113–121. [Google Scholar]
- Wallner, S; Hermetter, A; Mayer, B; Wascher, TC. The alpha-amino group of L-arginine mediates its antioxidant effect. Eur J Clin Invest 2001, 31, 98–102. [Google Scholar]
- Suetsuna, K. Separation and identification of peptides derived from soybean protein, and their active oxygen scavenging activity. J Jpn Soc Nutr Food Sci 1999, 52, 225–228. [Google Scholar]
- Chen, H-M; Muramoto, K; Yamauchi, F; Fujimoto, K; Nokihara, K. Antioxidative properties of histidine-containing peptides fragments found in the digests of a soybean protein. J Agric Food Chem 1998, 46, 49–53. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Casein (g/kg diet)
| Soybean protein (g/kg diet)
| ||||
|---|---|---|---|---|---|---|
| Group1 | Group2 | Group3 | Group4 | Group5 | Group6 | |
| Caseinb | 200 | 600 | 600 | |||
| soybean proteinb | 200 | 600 | 600 | |||
| Corn starchc | 580 | 220 | 220 | 580 | 220 | 220 |
| Sucrose | 60 | 20 | 20 | 60 | 20 | 20 |
| Soybean oild | 50 | 50 | 50 | 50 | 50 | 50 |
| Cellulose powdera | 50 | 50 | 50 | 50 | 50 | 50 |
| Mineral mixturee | 40 | 40 | 40 | 40 | 40 | 40 |
| Vitamin mixturef | 20 | 20 | 20 | 20 | 20 | 20 |
| Cysteaminec | 0.06 | 0.06 | ||||
| Groups | MDA(nmol/mg prot)
| SOD(U/mg prot) | ||||
|---|---|---|---|---|---|---|
| Duodenum | Liver | Pancreas | Duodenum | Liver | Pancreas | |
| Group1 | 2.67±0.12ab | 3.91±0.26a | 2.09±0.10a | 95.04±3.57de | 218.55±8.25c | 98.92±2.51c |
| Group2 | 4.76±0.26d | 7.92±0.16c | 4.99±0.12c | 61.27±4.23a | 158.28±4.73a | 68.74±0.50a |
| Group3 | 3.69± 0.15c | 5.84±0.16b | 3.36±0.14b | 79.66±2.36bc | 185.09±4.82b | 82.60±1.69b |
| Group4 | 2.28±0.13a | 4.44±0.41a | 1.82±0.09a | 98.35±3.46e | 204.33±8.83c | 101.44±1.78c |
| Group5 | 4.66±0.35d | 7.77±0.25c | 4.55±0.22c | 69.50±3.02ab | 166.96±3.48ab | 70.92±4.05a |
| Group6 | 3.17±0.26bc | 5.85±0.07b | 3.09±0.34b | 84.21±5.41cd | 182.67±2.61b | 85.77±4.34b |
| Group | GSH-Px(U/mg prot) | GSH(mg/g prot) | ||||
|---|---|---|---|---|---|---|
| Duodenum | Liver | Pancreas | Duodenum | Liver | Pancreas | |
| Group1 | 80.828±0.925e | 59.963±0.206c | 102.196±3.630b | 99.22±0.32d | 191.29±4.25c | 153.02±4.69c |
| Group2 | 58.748±1.073a | 50.688±2.038a | 88.169±4.244a | 38.91±2.02a | 78.23±1.93a | 123.38±2.19a |
| Group3 | 70.476±1.497c | 53.818±0.306ab | 97.560±5.194ab | 65.59±1.65b | 100.84±1.47b | 138.97±2.23b |
| Group4 | 82.610±0.324e | 61.197±2.009c | 100.362±1.224b | 116.27±3.98e | 184.34±6.07c | 165.78±3.09d |
| Group5 | 67.190±0.573b | 52.718±0.297ab | 91.768±2.533ab | 44.31±1.58a | 84.61±3.23a | 130.29±1.73a |
| Group6 | 76.610±0.566d | 54.930±0.248b | 97.689±1.692ab | 78.72±6.80c | 97.55±4.62b | 146.11±1.33bc |
| Groups | CAT(U/mg prot) | Na+ K+-ATPase(μmolPi/mg prot/hour) | ||||
|---|---|---|---|---|---|---|
| Duodenum | Liver | Pancreas | Duodenum | Liver | Pancreas | |
| Group1 | 56.17±0.75e | 82.16±0.21d | 78.65±0.51c | 21.53±0.32d | 10.67±0.07c | 3.14±0.29c |
| Group2 | 25.08±1.10a | 54.00±0.58a | 56.54±1.65a | 13.92±0.21a | 1.95±0.23a | 0.95±0.01a |
| Group3 | 42.44±1.33c | 73.13±1.46c | 68.32±0.66b | 15.97±0.23bc | 5.47±0.14b | 1.93±0.02b |
| Group4 | 57.77±0.85e | 82.13±1.48d | 78.88±0.35c | 22.53±1.02d | 10.40±0.18c | 3.72±0.44c |
| Group5 | 37.54±1.91b | 58.98±0.69b | 57.16±2.94a | 14.79±0.64ab | 2.59±0.51a | 0.99±0.06a |
| Group6 | 46.24±0.77d | 72.52±1.14c | 70.22±0.45b | 17.71±0.68c | 5.76±0.61b | 2.06±0.05b |
Share and Cite
Gu, C.; Shi, Y.; Le, G. Effect of Dietary Protein Level and Origin on the Redox Status in the Digestive Tract of Mice. Int. J. Mol. Sci. 2008, 9, 464-475. https://doi.org/10.3390/ijms9040464
Gu C, Shi Y, Le G. Effect of Dietary Protein Level and Origin on the Redox Status in the Digestive Tract of Mice. International Journal of Molecular Sciences. 2008; 9(4):464-475. https://doi.org/10.3390/ijms9040464
Chicago/Turabian StyleGu, Chunmei, Yonghui Shi, and Guowei Le. 2008. "Effect of Dietary Protein Level and Origin on the Redox Status in the Digestive Tract of Mice" International Journal of Molecular Sciences 9, no. 4: 464-475. https://doi.org/10.3390/ijms9040464
APA StyleGu, C., Shi, Y., & Le, G. (2008). Effect of Dietary Protein Level and Origin on the Redox Status in the Digestive Tract of Mice. International Journal of Molecular Sciences, 9(4), 464-475. https://doi.org/10.3390/ijms9040464