1. Introduction
There is an increasing need for fast and accurate quantification of bacteria concentrations in multiple fields such as environmental monitoring and hazard analysis in multiple industries, particularly the food industry. The purpose of the quantification is to ensure that the total bacteria concentrations in the production adhere to the regulations and guidelines set by national and international authorities and to ensure safe and high-quality products. Monitoring happens by collecting samples of cleaning water (e.g., CIP-water (clean-in-place) from production equipment) or by collecting samples from production surfaces [
1] (e.g., using a moist swab) and resuspending the collected bacteria in an appropriate buffer liquid. The bacterial concentrations in the collected samples are then analyzed using plate counting [
2]. This method is well established and reliable, but is notoriously slow and only shows the presence of culturable bacteria [
3]. Consequently, a need exists for a fast and reliable method for the quantification of bacteria concentrations in samples that can complement or replace traditional plate counting.
In recent years, a number of alternative technologies for the quantification of bacteria concentrations has emerged to overcome the limitations of traditional plate counting. Some examples are polymerase chain reaction-based (such as real-time PCR or qPCR) or fluorescence-based techniques (such as fluorescent-assisted cell sorting). However these methods are labor-intensive and require complicated sample processing with expensive reagents [
4]. Impedance Flow Cytometry (IFC) is an emerging technology for single-cell analysis and particle enumeration [
5]. This technology has shown its potential for single-cell analysis in samples containing a wide range of cell types. It has been used to characterize the electrical properties of Red Blood Cells (RBCs) [
6], to analyze plant spores [
7], to identify the differentiation state of Mesenchymal Stem Cells (MSCs) [
8], to characterize and classify tumor cells [
9], to study the effect of chemotherapy treatment on the cell membrane of HeLa cells [
10], to monitor the viability of yeast cells [
11] and to analyze pollen samples [
12]. Due to their small size, it can be difficult to achieve sufficient signal-to-noise when measuring bacteria using IFC. Several suggestions have been given on how to improve this [
13,
14,
15]. The spectral response of
E. coli has previously been studied using impedance spectroscopy techniques [
16,
17], and while some research has gone into specifically investigating the use of IFC to detect bacteria [
18,
19], assess the viability of Bacillus megaterium [
20] and to classify bacteria based on their species [
21], a general characterization of bacteria using IFC remains an underdeveloped research area.
In this paper we take a more general approach to the analysis of bacteria viability with IFC by including general inactivation methods such as ethanol and autoclaving. To our knowledge this is also the first time IFC has been used to study the viability of E. coli.
IFC has a number of advantages over competing methods for bacteria analysis, mainly that it is label-free, non-invasive, sensitive and fast. A traditional plate count will underestimate the true number of viable bacteria in non-laboratory cultures and environmental samples since it is not able to detect the presence of unculturable species or bacteria that are intact and metabolically active but unable to divide under ordinary conditions (viable but non-culturable bacteria) [
22,
23,
24,
25]. In IFC, every particle or cell in the samples is counted and characterized no matter what the species is or what viability state the bacteria is in. Consequently, IFC has the potential to provide a more accurate measurement of the total bacteria count by including cells which would previously have been unseen.
However, using IFC as a replacement for traditional plate counting may also introduce new technical challenges. When a bacterium is inactivated it loses its ability to divide and subsequently will not generate a visible colony (colony-forming unit, CFU) on an agar plate. In this case, the cell could retain enough of its cellular structure to still be counted using IFC and lead to a false positive detection of inactivated cells. It is therefore imperative that the change in dielectric response of inactivated bacteria is investigated and characterized, if IFC is going to emerge as a useful and reliable alternative to plate counting.
Despite being frequently used, the term bacteria viability does not have a straightforward definition [
26]. A comprehensive definition of viability considers cell component integrity, metabolic activity and ability to proliferate. However, in this article we will use viability (i.e., “a viable cell”) to mean either a cell that shows an intact cellular structure (specifically the cell membrane) or is able to give progeny on an appropriate agar and generate visible colonies. Similarly, we will use the term “inactivated cell” to mean a cell that has undergone an inactivation process so that it no longer fulfills the viability conditions described above. The inactivation agents used in this study are ethanol, heat (at 90 °C) and autoclaving. These methods are expected to act with rapid and nearly simultaneous oxidation or denaturation of multiple targets, including the cytoplasmic membrane, proteins, ribosomes, and/or DNA [
26].
In this paper we use a prototype multi-frequency impedance flow cytometer developed by the Danish company SBT Instruments A/S (Herlev, Denmark) to test the impedance response of Escherichia coli (E. coli). We inactivate the E. coli cells using three different methods (ethanol, 90 °C heat or autoclaving) and compare it to the impedance response of untreated E. coli cells. We also discuss the expected impact of the inactivation on the electrical properties of the bacteria. Furthermore, we compare the experimental impedance measurements with CFU counts and fluorescence imaging to determine whether the change in impedance response can be correlated to the cells’ ability to grow and the membrane integrity of the cells, respectively. Finally, we evaluate the sensitivity and specificity of IFC as a tool for characterizing inactivated and viable cells.
3. Results
Three different inactivation experiments were carried out using inactivation treatments with ethanol, heat and autoclavation, respectively. Shortly after each inactivation was completed, the untreated and treated samples were analyzed using the impedance flow cytometer and the concentration and electrical responses were obtained from the recorded data. The same samples were also analyzed using plate counting to investigate viability and fluorescent microscopy in order to investigate membrane integrity.
3.1. Concentrations of Bacteria Samples
Bacteria concentrations were studied after the inactivation treatment using drop plating and IFC in parallel. The concentrations measured with drop plating and IFC for the untreated and treated samples from the three inactivation experiments can be found in
Table 1. The concentration of bacteria in the untreated samples was roughly 10
8 CFU/mL in all three experiments. No standardization of the concentration (e.g., using optical density) was performed with the different untreated samples, which could reduce the variation in future experiments. When compared to the counts per milliliter provided by IFC, we see that IFC consistently measures lower concentrations. This difference is attributed to a lack of calibration in the prototype impedance flow cytometer and has been observed before [
21]. However, the standard deviation is significantly lower using IFC compared to the plate counts.
No growth was detected in the inactivated samples on the agar plates (inactivation efficiency of 100%). However, with IFC it was still possible to detect events after inactivation for all three inactivation methods. This suggests that some of the inactivated cells retain enough cell integrity after inactivation to be detected by the impedance system. In general, the lower concentration in the treated samples can be attributed to complete cell disintegration during the inactivation. Such a process is more likely to happen in
E. coli when heat is applied, especially for a prolonged time, compared to cells exposed to ethanol [
36], which corresponds well with the reduction in cell counts that we see for the three methods.
3.2. Characterization by Impedance
The measured argument and modulus of detected bacteria from each of the three inactivation experiments are shown in
Figure 3, together with the measured response of a reference sample of polystyrene beads. It is evident that the beads and untreated bacteria populations, individually, appear in the same position for each of the three experiments, demonstrating that the observed changes in impedance response of the treated cells is related to the inactivation process and not the general sample handling.
Figure 3 shows the differentiation of 1.5 µm beads, untreated
E. coli and treated
E. coli based on the low- and high-frequency arguments. For the ethanol and autoclave inactivation experiment we observed a significant shift between untreated and treated bacteria, except in different directions. However, no such shift was observed in the heat inactivation experiment.
The measured argument from the ethanol inactivation experiment (
Figure 3a), allows us to distinguish a population for each sample with only minor overlap. The untreated bacteria and the beads show a separation between sample populations experiencing the high frequency due to the short-circuiting of the membrane capacitance, while the responses in the low-frequency condition are more similar. The untreated population of cells shows a greater variation compared to the beads as can be expected in biological samples. Comparing the ethanol-treated population to the untreated population, we observe a shift primarily in the low-frequency argument. This corresponds well with the expected response, where disruptions of the membrane lower the effective membrane resistivity and allow the electric field to penetrate the cells even at lower frequencies.
Unexpectedly, it is not possible to differentiate bacteria treated at 90 °C for 5 min from untreated bacteria (
Figure 3b) in either the low- or high-frequency argument. This suggests that the treated bacteria have similar electrical properties to untreated bacteria and still retain a high degree of membrane integrity.
The population from the autoclaved sample shows a shift towards the beads in both the high-frequency and low-frequency argument, differentiating the majority of the autoclaved bacteria from the untreated bacteria (
Figure 3c). Due to the high pressure and temperature of the autoclave, it is likely that the bacteria shattered and that we observed cells with highly damaged cell walls. If the cell wall is damaged to a degree in which the majority of the cytoplasm has been replaced by electrolyte, the electrical response would be similar to what we observed.
The modulus of the differential current is different for the three treated samples. The population of ethanol-treated bacteria (
Figure 3d) is narrow with low variation and a slope close to 1, aligning well with the idea of a disrupted membrane, where current can flow through equally well in both low and high frequencies.
The heat-treated sample (
Figure 3e) shows a variation in modulus similar to that of the untreated bacteria but with a smaller magnitude overall.
Similar to the heat-treated sample, the modulus for the autoclaved population (
Figure 3f) is smaller compared to the untreated sample. However, for this sample we see a relatively small high frequency (HF) modulus compared to the low frequency (LF) modulus. This could support the idea that the majority of the cytoplasm has been replaced by electrolyte. At low frequencies, the disrupted membrane lets the current flow through the cell, but the intact part of the membrane still provides some resistance to the current flow. However, at higher frequencies the capacitance of the lipid membrane is short-circuited leading to cell walls that are more “transparent” and, due to the exchange of cytoplasm, a smaller differential current.
3.3. Fluorescence Imaging
To check whether it is indeed a loss of membrane integrity that causes the changes in impedance response, we investigated the bacteria using fluorescent staining.
Images of untreated and treated fluorescent bacteria and their corresponding membrane integrity percentages are shown in
Figure 4. As expected, untreated samples showed a high degree of membrane integrity across all three experiments: 92.3% for ethanol, 98.7% for heat and 94.8% for autoclave. The deviation from a 100% membrane integrity is expected to be due to a small percentage of fast-growing cells with compromised membranes [
37] or bacteria that were structurally damaged during sample preparation.
The bacteria in all three treated samples exhibited low membrane integrity percentages: 7.3% for ethanol, 6.9% for heat and 1.7% for autoclave. However, these percentages are still higher than we would expect considering the plate counts (0% viability for all treatments). This could mean that a small population of bacteria keep membrane integrity but are unable to divide, or that there is an unknown intrinsic error related to the staining protocol.
These results show that all three inactivation methods used in this study—ethanol, heat and autoclave—disrupt the membrane of the bacteria. It is worth mentioning that the total number of bacteria was substantially lower in the treated samples with respect to the untreated samples. This evidence aligns with the results obtained from the plate counts for each experiment.
3.4. Characterization of Treated and Untreated Bacteria
So far, the measured current response has been investigated and correlated to the biological state of the samples. However, in order to use IFC as an alternative to plate counts (e.g., as a risk assessment tool in the food industry) it is also important to consider the effectiveness of the viability classification of the technology—i.e., investigate how well the technology can characterize viable cells as viable and how well it can characterize inactivated cells as inactivated. We therefore investigated the classification of untreated and treated bacteria for each of the three inactivation methods and determined the optimal classification threshold for each of them. We achieved this by using ROC curves as seen in
Figure 5. Based on the individual thresholds for each of the inactivation methods, we found two thresholds that together return the best classification between untreated and treated bacteria across all samples. The thresholds were set in the high-frequency argument since this yielded the overall best sensitivity and selectivity. A similar analysis for the low-frequency argument can be found in the
Section 2.1 of the Supplementary Materials.
3.4.1. ROC with Single HF Argument Threshold
Figure 5 shows the ROC curves obtained for the HF argument from the three inactivation experiments. Optimal threshold values that classify untreated and treated
E. coli were identified as 2.22, 1.78 and 0.63 for the ethanol, heat, and autoclavation inactivation experiments, respectively. The optimal thresholds were chosen in order to maximize sensitivity and selectivity.
The classification of untreated bacteria and bacteria treated with either ethanol or autoclavation is strong, with a sensitivity of 99.5% and 95.5%, and a selectivity of 96.7% and 88.8%, respectively. The area under the curve (AUC) values calculated for each ROC curve can be found in
Table 2 and show an AUC of 0.97 and 0.93 for the ethanol experiment and autoclave experiment. However, for the heat-treated cells, the classification is quite poor with and AUC of 0.62, a sensitivity of 60.7% and a selectivity of 60.0%. This corresponds well with the observed differences in dielectric responses seen for the three experiments (
Figure 3).
Included in the figure are three categorical scatter plots including the used thresholds for the HF argument. The categorical scatter plots show that the identified thresholds work well for separating untreated and treated cells in the ethanol and autoclavation experiments but not in the heat experiment.
A summary of the AUC, sensitivity and selectivity found for each optimal threshold can be found in
Table 2.
In the
Section 2.2 of the supplementary materials, three additional repetitions of the ethanol inactivation experiment are shown. The additional experiments displayed good repeatability of the results with sensitivities of 99.1%, 98.3% and 99.1%, and selectivities of 97.0%, 86.5% and 98.8%.
3.4.2. ROC with Multiple HF Argument Thresholds
We have identified two thresholds (one from the ethanol inactivation experiment and one from the autoclavation experiment) that work well for classifying untreated bacteria from ethanol-treated and autoclaved bacteria, respectively. In a real-world situation, we cannot be sure how the bacteria are inactivated and therefore want to establish a general threshold that is as effective as possible for most situations. We therefore combine the two thresholds and classify every event between the thresholds as untreated bacteria and every event outside the thresholds as non-bacteria.
The thresholds are illustrated in
Figure 6a and the sensitivity and selectivity for each of the three treated samples are summarized in
Figure 6b. We see that the combined thresholds are generally good at classifying untreated cells, ethanol-treated cells and autoclaved cells with sensitivities and selectivities >90%. Similarly, the selectivity of the system towards polystyrene beads was >99.7% in all three experiments. However, the combined threshold is still very poor at classifying the heat-treated cells with a selectivity of only 18%.
Even so, if the reduction in cell count from cell disruption during inactivation is combined with the improved threshold limits from the ROC analysis (
), the total reduction in detected viable bacteria szs found to be 100% after ethanol treatment, 85% after heat treatment and 99% after autoclavation (see
Figure 6c).
Future experiments should focus on investigating the impedance response of the inactivated bacteria at additional frequencies in order to optimize the sensitivity and selectivity of the system. The full spectral sweep could be obtained by measurements using a chip with interdigitated electrodes similar to what is carried out in [
16] and determining the frequencies with the highest differentiation between intact and inactivated cells. This is, of course, of particular interest when investigating the heat-inactivated samples, where improved sensitivity is crucial for the application of the technology. Special focus should be put on trying to understand why the impedance response of a heat-treated bacteria appears similar to untreated cells, even though they do not grow. It would be interesting to understand whether the number and/or size of the disruptions in the membrane play a role in this. Additionally, it would be interesting to monitor the cellular degradation of the cells over time (1–24 h) after inactivation with all three methods.
4. Discussion and Conclusions
In this paper we have shown how an impedance flow cytometer prototype can be used to detect and characterize E. coli bacteria that have been inactivated with ethanol, heat and autoclaving.
We compared the bacteria counts from IFC with those obtained from plate counts for treated and untreated bacteria and saw that the inactivation processes yielded zero CFU counts, but that the impedance flow cytometer still detected cells regardless of the viability state showcasing the increased sensitivity of the technology.
We have shown that the impedance response of bacteria changes significantly upon ethanol inactivation. This observation reinforces the belief that ethanol inactivates without necessarily dissolving the entire cell but strongly affecting the integrity of the membrane. By analyzing the impedance response of the ethanol-treated cells we were able to classify the treated cells from untreated cells with a selectivity of 99.6%.
Similarly, the impedance response of bacteria that remained after autoclaving also changed, although differently from the ethanol-inactivated cells. Here the impedance change indicates a broken cell membrane in combination with a replacement of cytoplasm with the surrounding electrolyte. By analyzing the impedance response of the autoclaved treated cells, we were able to classify the treated cells from untreated cells with a selectivity of 90.6%.
However, no significant differences were observed in the impedance response of heat-inactivated cells. This suggests that no measurable changes occurred in the cell bacteria properties, or that the system we used is not fully optimized to detect those changes. The selectivity towards treated cells was only 18.0%.
Nevertheless, investigations with fluorescent dyes showed that the cell membrane was indeed disrupted after inactivation with all three methods, proving that the structure of the cell membrane changes during inactivation. These results demonstrate that IFC can be used to detect and characterize the membrane integrity of E. coli bacteria, but with the current flow cytometer it is not possible for all inactivation methods. However, further investigation using different frequencies could improve this.
The need for a fast and accurate quantification of bacteria concentrations in the food industry, and other fields, is still commercially unfulfilled. This work has shown that IFC could be a promising candidate to fulfil this need, as the technology shows potential in differentiating between bacteria samples that are treated and untreated using a number of different inactivation methods.

 ,
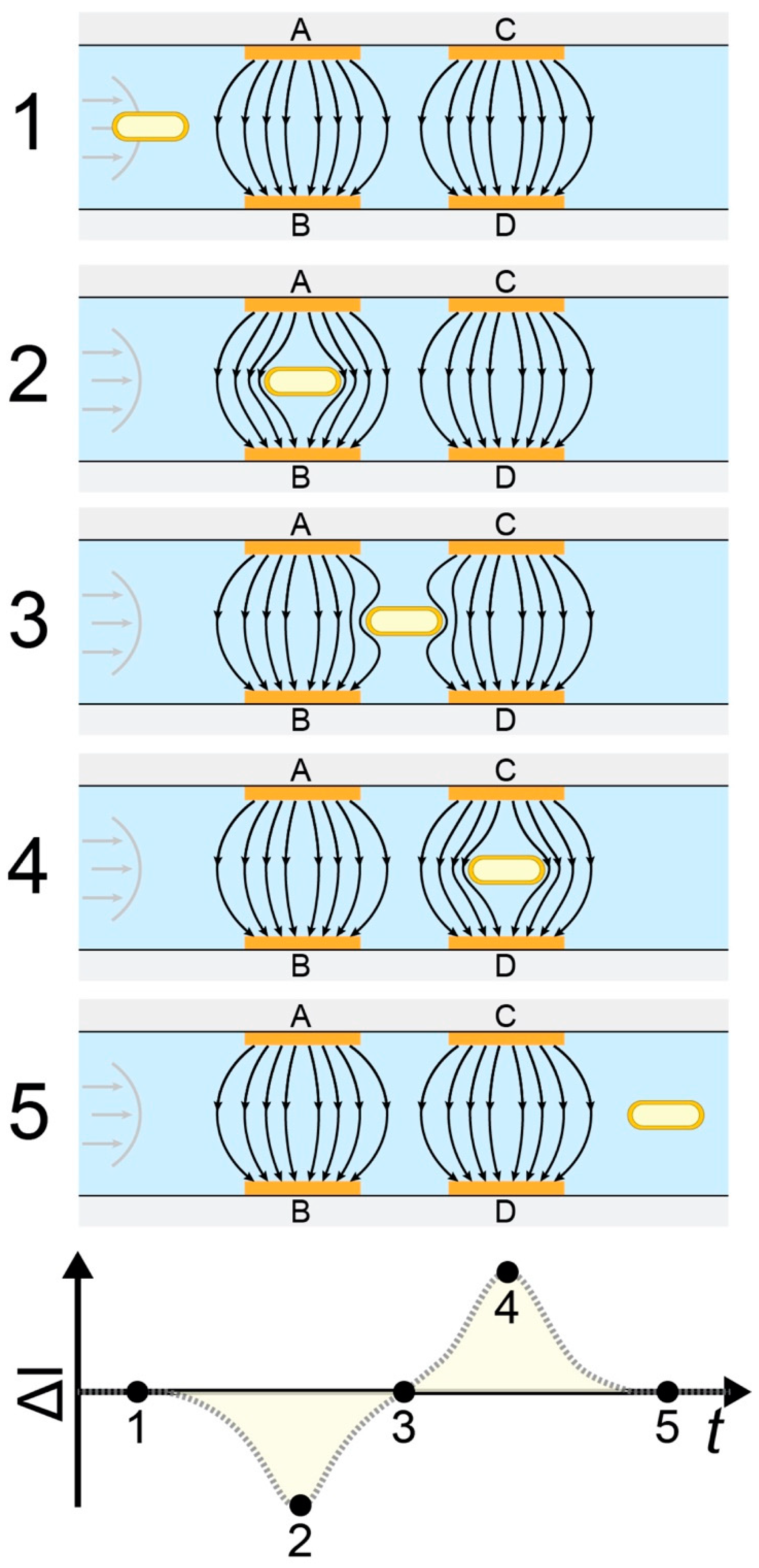
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}