
Strategies for Mitigating Commercial Sensor Chip Variability with Experimental Design Controls



Abstract
1. Introduction
2. Materials and Methods
2.1. Recombinant Repeat Expression and Purification
2.2. Characterization of Tandem Repeat Protein–Antibody Binding Affinity
2.3. Comparison of Inter- and Intra-Chip Variability
2.4. Measurement of Maximum Sensor Immobilization
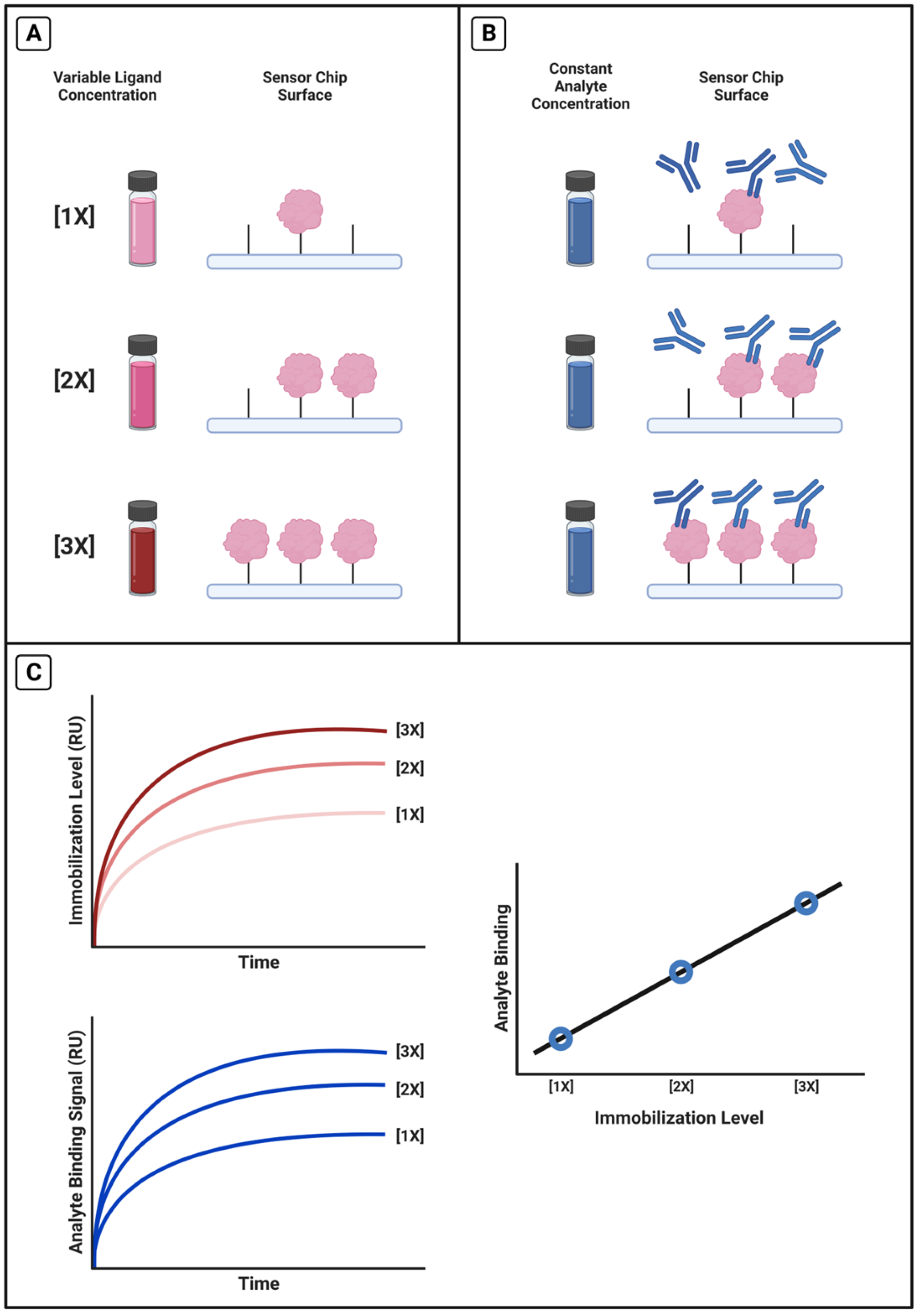
2.5. Calibrating the Relationship between Immobilization Level and Antibody Signal Magnitude
2.6. Normalization of HE4–Antibody Binding Affinity
3. Results and Discussion
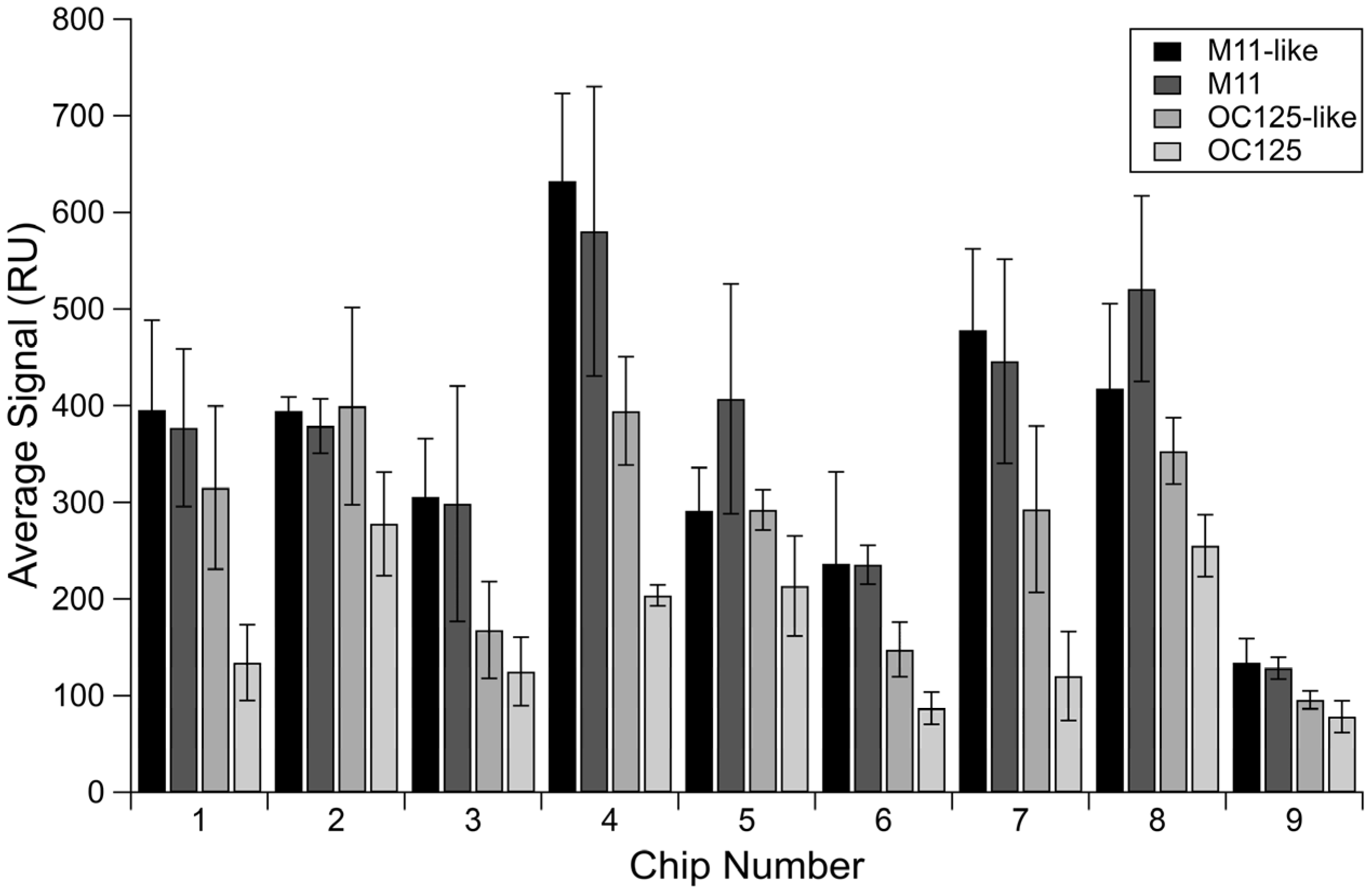
3.1. Non-Specific Binding of Antibodies across a Selection of Chips
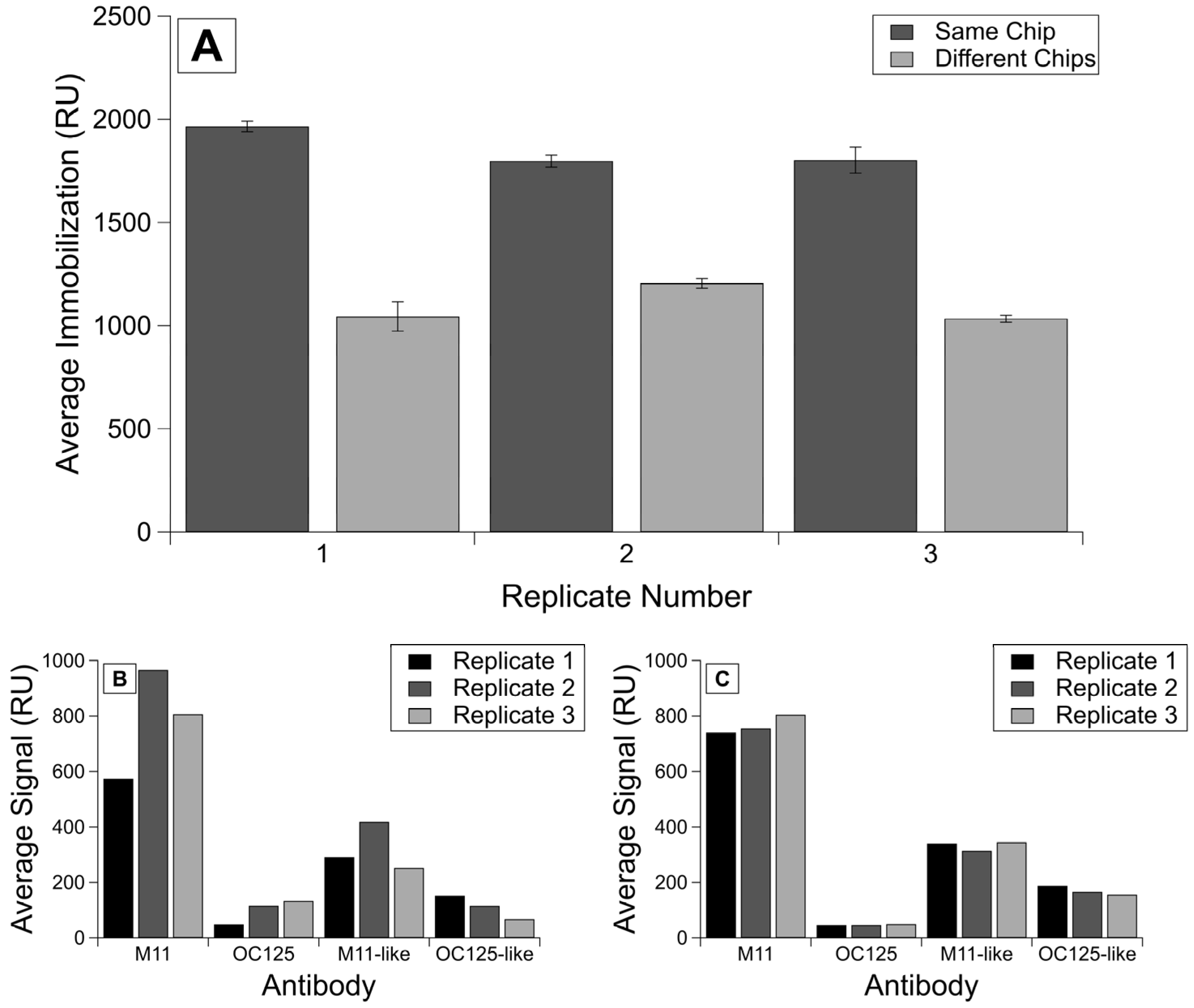
3.2. Comparison of Inter- and Intra-Chip Variability
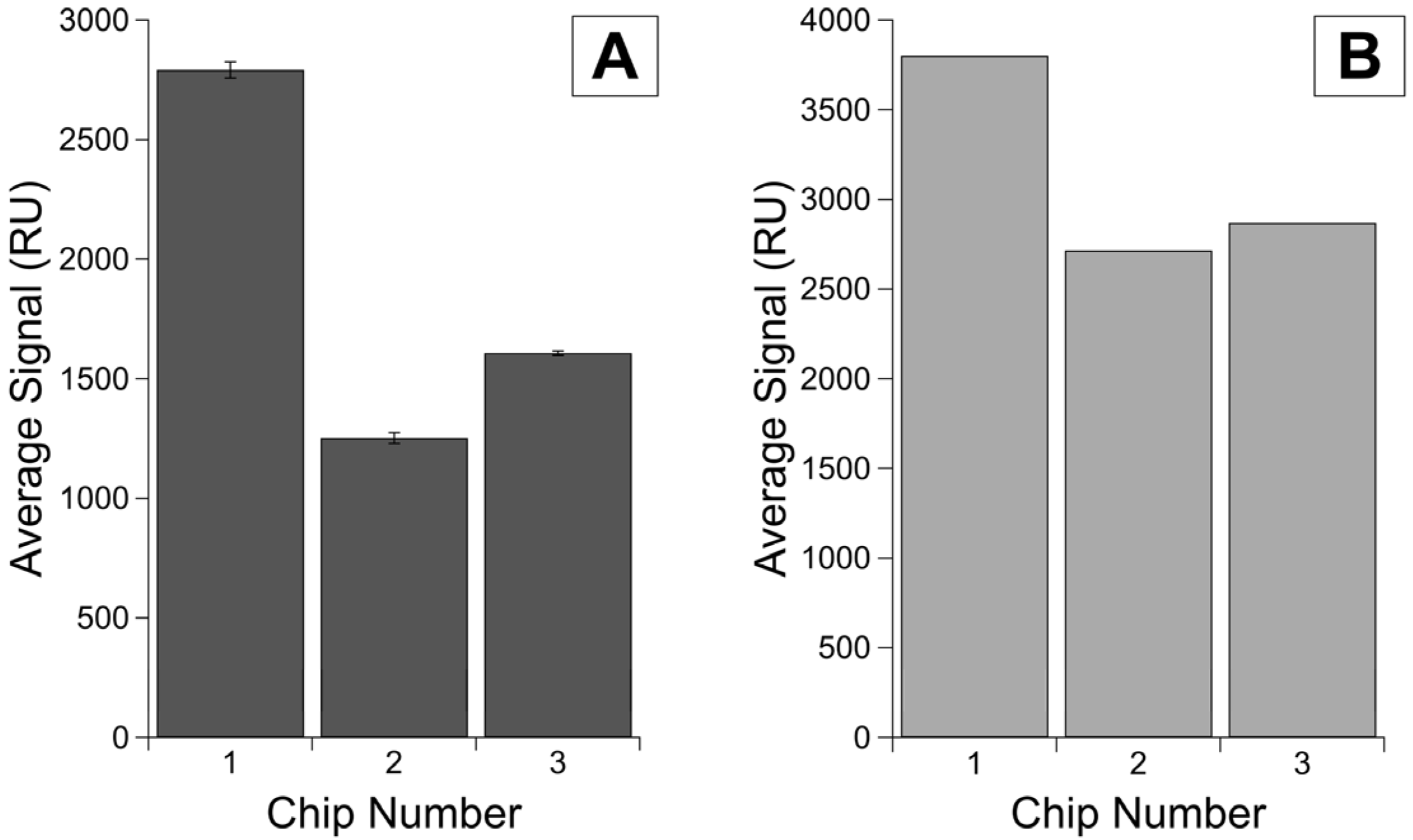
3.3. Establishing a Maximum Immobilization on Different Chips
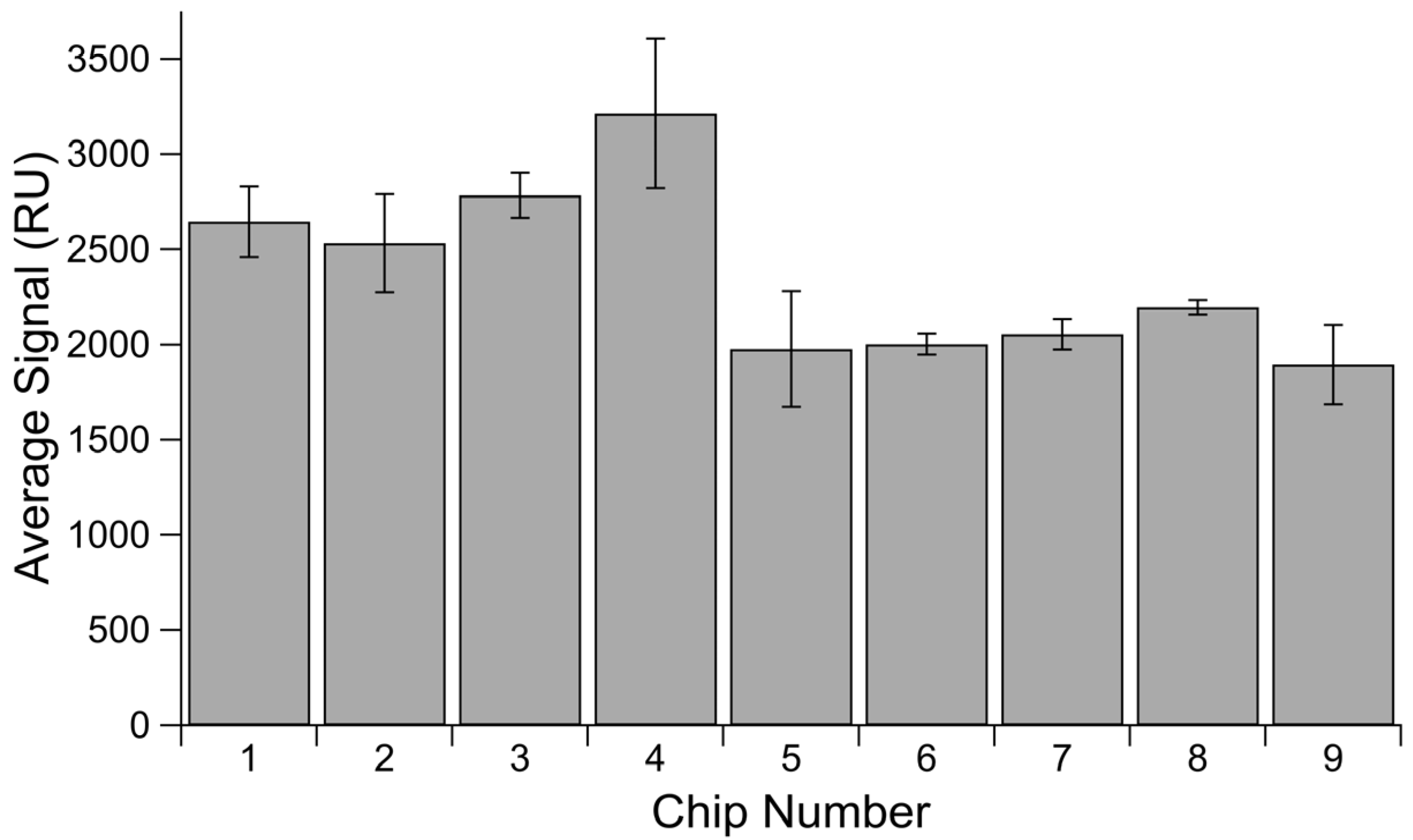
3.4. Chip Thickness
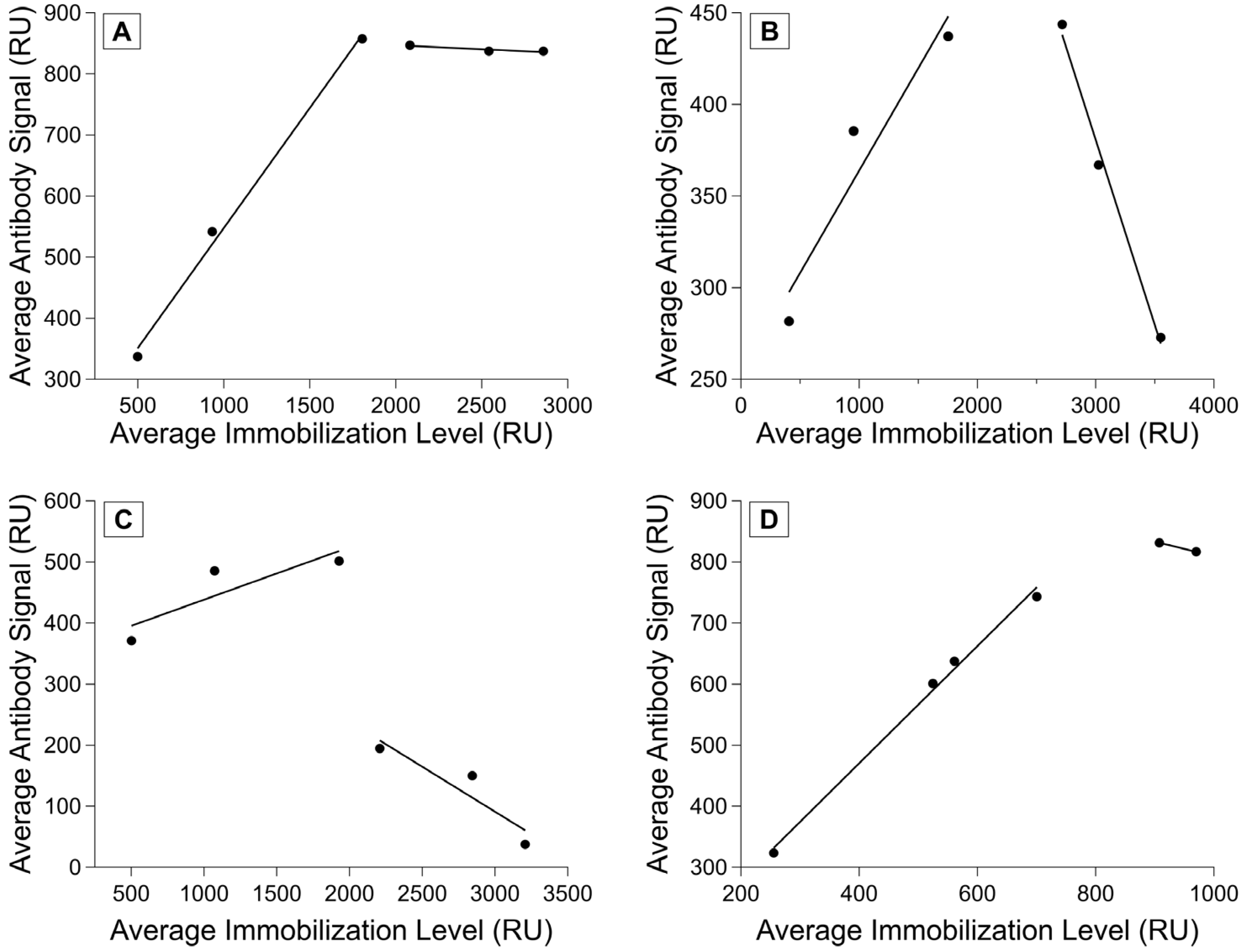
3.5. Calibrating the Relationship between Antibody Signal and Immobilization Level
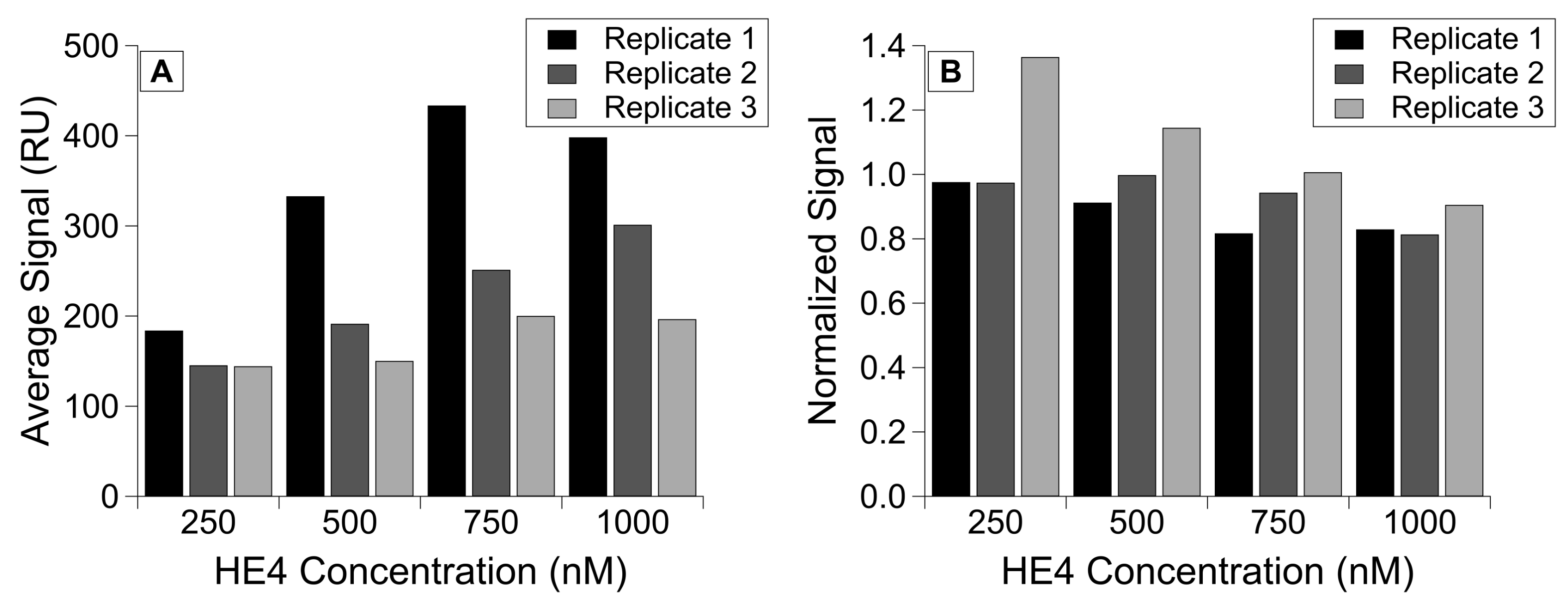
3.6. Normalizing Binding Affinity Data to Account for Immobilization Variability
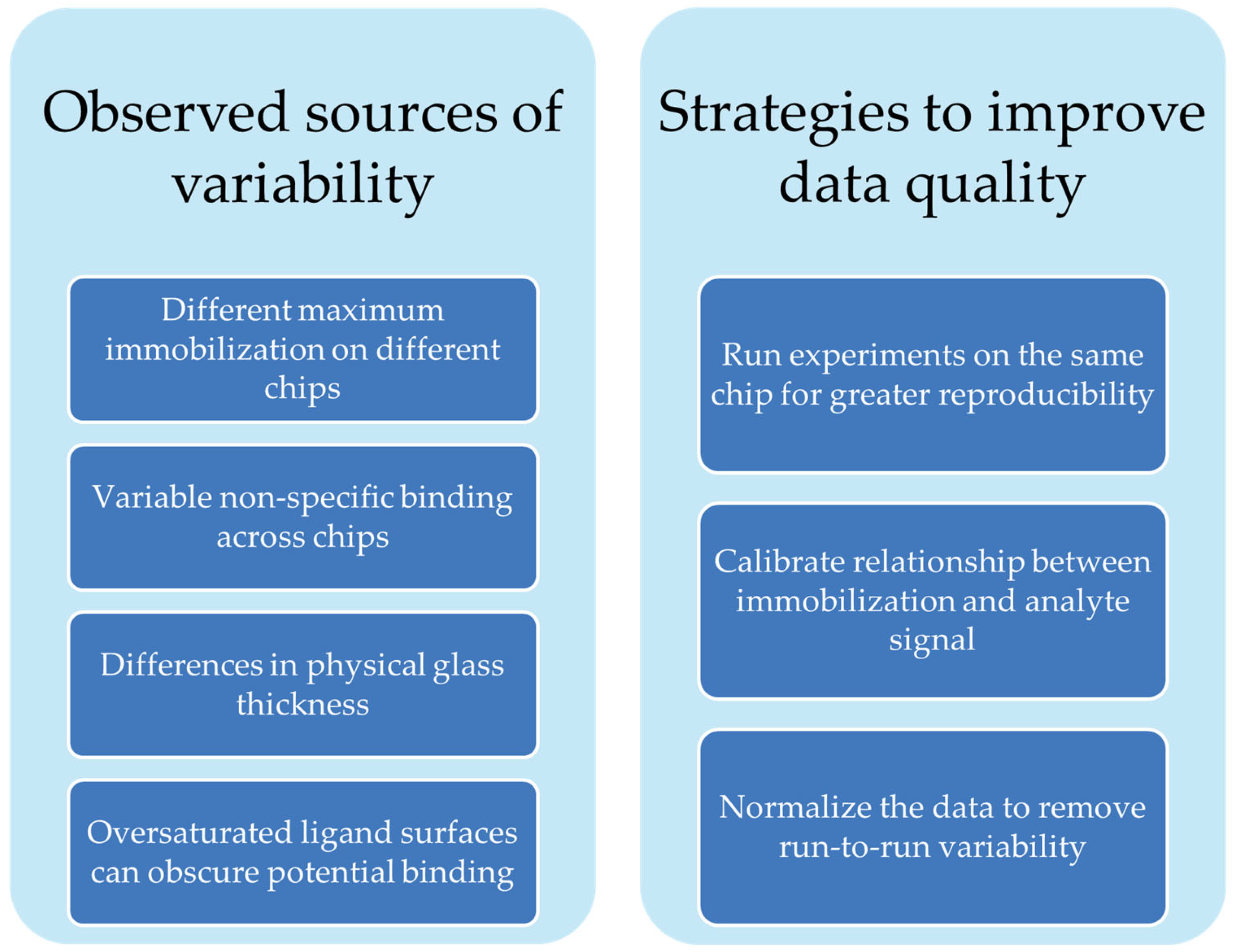
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liedberg, B.; Nylander, C.; Lundstrom, I. Surface Plasmon Resonance for Gas Detection and Biosensing. Sens. Actuators 1983, 4, 299–304. [Google Scholar] [CrossRef]
- Liedberg, B.; Nylander, C.; Lundström, I. Biosensing with surface plasmon resonance—How it all started. Biosens. Bioelectron. 1995, 10, i–ix. [Google Scholar] [CrossRef]
- Hou, Y.; Che, D.; Wei, D.; Wang, C.; Xie, Y.; Zhang, K.; Cao, J.; Fu, J.; Zhou, N.; He, H. Phenothiazine antipsychotics exhibit dual properties in pseudo-allergic reactions: Activating MRGPRX2 and inhibiting the H(1) receptor. Mol. Immunol. 2019, 111, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Rich, R.L.; Hoth, L.R.; Geoghegan, K.F.; Brown, T.A.; LeMotte, P.K.; Simons, S.P.; Hensley, P.; Myszka, D.G. Kinetic analysis of estrogen receptor/ligand interactions. Proc. Natl. Acad. Sci. USA 2002, 99, 8562–8567. [Google Scholar] [CrossRef]
- Duellman, T.; Chen, X.; Wakamiya, R.; Yang, J. Nucleic acid-induced potentiation of matrix metalloproteinase-9 enzymatic activity. Biochem. J. 2018, 475, 1597–1610. [Google Scholar] [CrossRef] [PubMed]
- Teh, H.F.; Peh, W.Y.X.; Su, X.; Thomsen, J.S. Characterization of Protein–DNA Interactions Using Surface Plasmon Resonance Spectroscopy with Various Assay Schemes. Biochemistry 2007, 46, 2127–2135. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, K.; Jeong, E.-J.; Shin, Y.-B.; Chung, B.H. Surface plasmon resonance imaging analysis of protein–protein interactions using on-chip-expressed capture protein. Anal. Biochem. 2006, 351, 298–304. [Google Scholar] [CrossRef]
- Sandoval, D.R.; Gomez Toledo, A.; Painter, C.D.; Tota, E.M.; Sheikh, M.O.; West, A.M.V.; Frank, M.M.; Wells, L.; Xu, D.; Bicknell, R.; et al. Proteomics-based screening of the endothelial heparan sulfate interactome reveals that C-type lectin 14a (CLEC14A) is a heparin-binding protein. J. Biol. Chem. 2020, 295, 2804–2821. [Google Scholar] [CrossRef]
- Chen, W.; Qiang, X.; Wang, Y.; Zhu, S.; Li, J.; Babaev, A.; Yang, H.; Gong, J.; Becker, L.; Wang, P.; et al. Identification of tetranectin-targeting monoclonal antibodies to treat potentially lethal sepsis. Sci. Transl. Med. 2020, 12, eaaz3833. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, L.; Zhou, B.; Wang, X.; Liu, G.; Liu, W.; Wang, P. Investigation of biological cell–protein interactions using SPR sensor through laser scanning confocal imaging–surface plasmon resonance system. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 121, 381–386. [Google Scholar] [CrossRef]
- Bhandari, D.; Chen, F.-C.; Hamal, S.; Bridgman, R.C. Kinetic Analysis and Epitope Mapping of Monoclonal Antibodies to Salmonella Typhimurium Flagellin Using a Surface Plasmon Resonance Biosensor. Antibodies 2019, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Sibille, P.; Strosberg, A.D. FIV epitope defined by a phage peptide library screened with a monoclonal anti-FIV antibody. Immunol. Lett. 1997, 59, 133–137. [Google Scholar] [CrossRef]
- Chang, C.-C.; Chiu, N.-F.; Lin, D.S.; Chu-Su, Y.; Liang, Y.-H.; Lin, C.-W. High-Sensitivity Detection of Carbohydrate Antigen 15-3 Using a Gold/Zinc Oxide Thin Film Surface Plasmon Resonance-Based Biosensor. Anal. Chem. 2010, 82, 1207–1212. [Google Scholar] [CrossRef] [PubMed]
- Uludag, Y.; Tothill, I.E. Cancer Biomarker Detection in Serum Samples Using Surface Plasmon Resonance and Quartz Crystal Microbalance Sensors with Nanoparticle Signal Amplification. Anal. Chem. 2012, 84, 5898–5904. [Google Scholar] [CrossRef] [PubMed]
- Adikaram, P.R.; Zhang, J.H.; Kittock, C.M.; Pandey, M.; Hassan, S.A.; Lue, N.G.; Wang, G.; Gucek, M.; Simonds, W.F. Development of R7BP inhibitors through cross-linking coupled mass spectrometry and integrated modeling. Commun. Biol. 2019, 2, 338. [Google Scholar] [CrossRef] [PubMed]
- Raether, H. Surface Plasma Oscillations as a Tool for Surface Examinations. Surf. Sci. 1967, 8, 233–246. [Google Scholar] [CrossRef]
- Cooper, M.A. Label-free screening of bio-molecular interactions. Anal. Bioanal. Chem. 2003, 377, 834–842. [Google Scholar] [CrossRef]
- Šípová, H.; Homola, J. Surface plasmon resonance sensing of nucleic acids: A review. Anal. Chim. Acta 2013, 773, 9–23. [Google Scholar] [CrossRef]
- Lee, K.-S.; El-Sayed, M.A. Gold and Silver Nanoparticles in Sensing and Imaging: Sensitivity of Plasmon Response to Size, Shape, and Metal Composition. J. Phys. Chem. B 2006, 110, 19220–19225. [Google Scholar] [CrossRef]
- Rani, M.; Sharma, N.K.; Sajal, V. Localized surface plasmon resonance based fiber optic sensor with nanoparticles. Opt. Commun. 2013, 292, 92–100. [Google Scholar] [CrossRef]
- Chauhan, M.; Singh, V.K. Review on recent experimental SPR/LSPR based fiber optic analyte sensors. Opt. Fiber Technol. 2021, 64, 102580. [Google Scholar] [CrossRef]
- Hanson, E.K.; Whelan, R.J. Application of the Nicoya OpenSPR to Studies of Biomolecular Binding: A Review of the Literature from 2016 to 2022. Sensors 2023, 23, 4831. [Google Scholar] [CrossRef]
- Nguyen, H.H.; Park, J.; Kang, S.; Kim, M. Surface Plasmon Resonance: A Versatile Technique for Biosensor Applications. Sensors 2015, 15, 10481–10510. [Google Scholar] [CrossRef]
- Wang, C.W.; Hanson, E.K.; Minkoff, L.; Whelan, R.J. Individual recombinant repeats of MUC16 display variable binding to CA125 antibodies. Cancer Biomark. 2023, 37, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Yoshitani, N.; Saito, K.; Saikawa, W.; Asanuma, M.; Yokoyama, S.; Hirota, H. NTA-mediated protein capturing strategy in screening experiments for small organic molecules by surface plasmon resonance. Proteomics 2007, 7, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Han, C.; Jiang, M.; Zhang, T.; Kang, Q.; Wang, X.; Wang, P.; Zhou, F. Rapid and regenerable surface plasmon resonance determinations of biomarker concentration and biomolecular interaction based on tris-nitrilotriacetic acid chips. Anal. Chim. Acta 2021, 1170, 338625. [Google Scholar] [CrossRef]
- Cai, J.; Wang, Z.; Wang, M.; Zhang, D. Au nanoparticle-grafted hierarchical pillars array replicated from diatom as reliable SERS substrates. Appl. Surf. Sci. 2021, 541, 148374. [Google Scholar] [CrossRef]
- Ma, Y.; Song, M.; Li, L.; Lao, X.; Wong, M.-C.; Hao, J. Advances in upconversion luminescence nanomaterial-based biosensor for virus diagnosis. Exploration 2022, 2, 20210216. [Google Scholar] [CrossRef] [PubMed]
- Awiaz, G.; Lin, J.; Wu, A. Recent advances of Au@Ag core-shell SERS-based biosensors. Exploration 2023, 3, 20220072. [Google Scholar] [CrossRef]
- O’Brien, T.J.; Beard, J.B.; Underwood, L.J.; Dennis, R.A.; Santin, A.D.; York, L. The CA 125 Gene: An Extracellular Superstructure Dominated by Repeat Sequences. Tumor Biol. 2001, 22, 348–366. [Google Scholar]
- Good, D.; (Nicoya Lifesciences, Kitchener, ON, Canada). Personal communication.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M11 | OC125 | M11-like | OC125-like | |
|---|---|---|---|---|
| Maximum | 580.79 | 277.76 | 632.56 | 399.58 |
| Minimum | 128.49 | 78.42 | 134.24 | 95.77 |
| Range | 452.30 | 199.34 | 498.32 | 303.81 |
| Chip Condition | Immobilization | M11 Binding | OC125 Binding | M11-like Binding | OC125-like Binding |
|---|---|---|---|---|---|
| R58 Same | 132.1 | 126.2 | 39.9 | 38.3 | 18,522.0 |
| R58 Different | 386.3 | 170.1 | 133.4 | 199.0 | 4983.9 |
| R9 Same | 60.8 | 44.4 | 42.8 | 48.7 | 95.6 |
| R9 Different | 105.1 | 252.2 | 449.2 | 271.1 | 379.3 |
| Chip Lot Number | Chip Thickness | Count |
|---|---|---|
| Unknown | 0.92 | 3 |
| SNE0111 | 0.93 | 4 |
| Unknown | 0.96 | 8 |
| SHE1101 | 0.97 | 4 |
| SCD1105 | 0.97 | 3 |
| Unknown | 0.97 | 8 |
| SCE0428 | 0.98 | 3 |
| Unknown | 0.98 | 10 |
| SND0928 | 0.99 | 1 |
| Unknown | 0.99 | 9 |
| SHE1129 | 1.00 | 2 |
| SND0927 | 1.00 | 1 |
| Unknown | 1.00 | 13 |
| SPD0907 | 1.01 | 1 |
| SQD0322 | 1.01 | 1 |
| Unknown | 1.01 | 14 |
| Unknown | 1.02 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanson, E.K.; Wang, C.-W.; Minkoff, L.; Whelan, R.J. Strategies for Mitigating Commercial Sensor Chip Variability with Experimental Design Controls. Sensors 2023, 23, 6703. https://doi.org/10.3390/s23156703
Hanson EK, Wang C-W, Minkoff L, Whelan RJ. Strategies for Mitigating Commercial Sensor Chip Variability with Experimental Design Controls. Sensors. 2023; 23(15):6703. https://doi.org/10.3390/s23156703
Chicago/Turabian StyleHanson, Eliza K., Chien-Wei Wang, Lisa Minkoff, and Rebecca J. Whelan. 2023. "Strategies for Mitigating Commercial Sensor Chip Variability with Experimental Design Controls" Sensors 23, no. 15: 6703. https://doi.org/10.3390/s23156703
APA StyleHanson, E. K., Wang, C.-W., Minkoff, L., & Whelan, R. J. (2023). Strategies for Mitigating Commercial Sensor Chip Variability with Experimental Design Controls. Sensors, 23(15), 6703. https://doi.org/10.3390/s23156703