
Living Together, Singing Together: Revealing Similar Patterns of Vocal Activity in Two Tropical Songbirds Applying BirdNET


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Study Area
2.3. Recording Protocol
2.4. Acoustic Data Analysis
2.5. BirdNET Performance Evaluation
2.6. Statistical Analyses
3. Results
3.1. BirdNET Performance
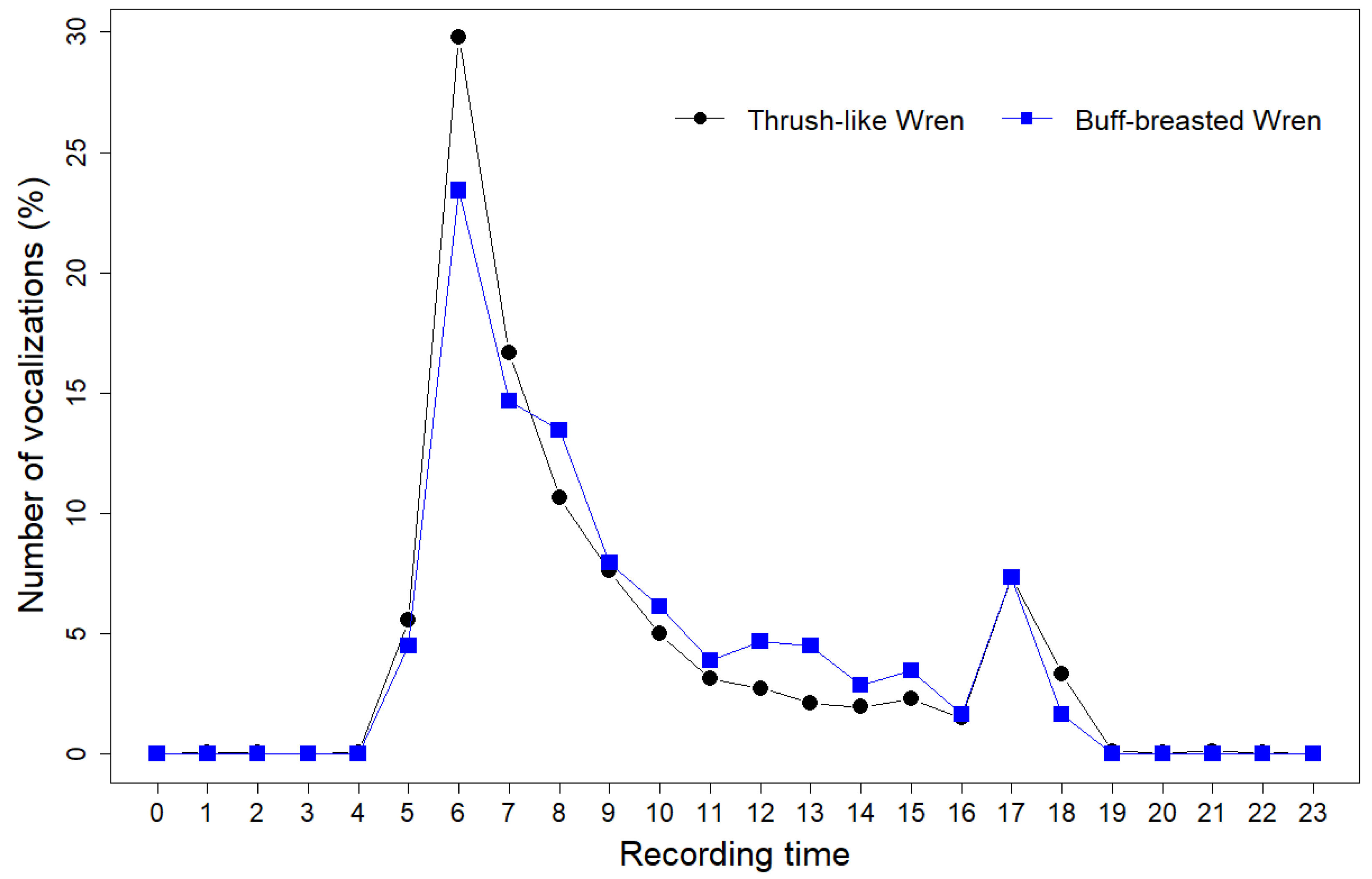
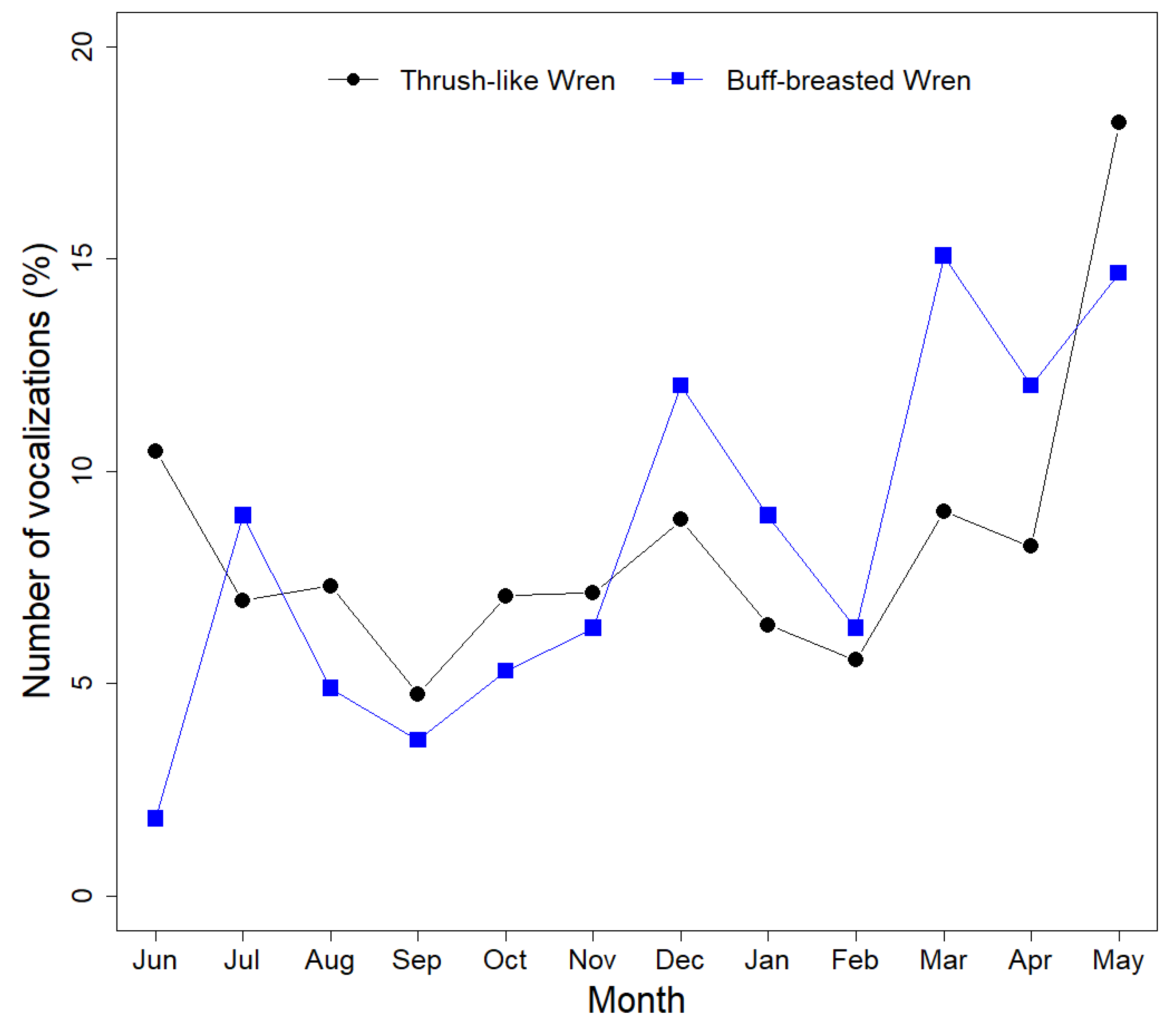
3.2. Vocal Activity Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hour | Station A | Station B | Station C | Station D | Station E | Total | % |
|---|---|---|---|---|---|---|---|
| 00 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 01 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 02 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 03 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 04 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 05 | 0 | 16 | 2 | 1 | 3 | 22 | 4.48 |
| 06 | 11 | 32 | 41 | 8 | 23 | 115 | 23.42 |
| 07 | 4 | 28 | 18 | 2 | 20 | 72 | 14.66 |
| 08 | 7 | 12 | 23 | 13 | 11 | 66 | 13.44 |
| 09 | 1 | 3 | 15 | 14 | 6 | 39 | 7.94 |
| 10 | 2 | 0 | 10 | 11 | 7 | 30 | 6.11 |
| 11 | 2 | 0 | 5 | 4 | 8 | 19 | 3.87 |
| 12 | 1 | 1 | 12 | 9 | 0 | 23 | 4.68 |
| 13 | 2 | 4 | 11 | 5 | 0 | 22 | 4.48 |
| 14 | 1 | 0 | 7 | 6 | 0 | 14 | 2.85 |
| 15 | 2 | 3 | 5 | 4 | 3 | 17 | 3.46 |
| 16 | 0 | 0 | 7 | 0 | 1 | 8 | 1.63 |
| 17 | 2 | 16 | 4 | 5 | 9 | 36 | 7.33 |
| 18 | 0 | 0 | 5 | 2 | 1 | 8 | 1.63 |
| 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 21 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 22 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| TOTAL | 350 | 115 | 165 | 84 | 92 | 491 |
| Hour | Station A | Station B | Station C | Station D | Station E | Total | % |
|---|---|---|---|---|---|---|---|
| 00 | 0 | 0 | 2 | 0 | 1 | 3 | 0.02 |
| 01 | 0 | 0 | 1 | 2 | 1 | 4 | 0.03 |
| 02 | 1 | 0 | 1 | 1 | 1 | 4 | 0.03 |
| 03 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 04 | 0 | 2 | 3 | 1 | 4 | 10 | 0.07 |
| 05 | 6 | 10 | 8 | 466 | 267 | 757 | 5.56 |
| 06 | 221 | 125 | 41 | 2661 | 1005 | 4053 | 29.78 |
| 07 | 215 | 85 | 11 | 1270 | 689 | 2270 | 16.68 |
| 08 | 150 | 32 | 3 | 822 | 443 | 1450 | 10.65 |
| 09 | 97 | 19 | 2 | 665 | 250 | 1033 | 7.59 |
| 10 | 73 | 11 | 5 | 464 | 125 | 678 | 4.98 |
| 11 | 48 | 6 | 1 | 227 | 144 | 426 | 3.13 |
| 12 | 59 | 4 | 0 | 164 | 144 | 371 | 2.73 |
| 13 | 30 | 6 | 1 | 129 | 121 | 587 | 2.11 |
| 14 | 71 | 3 | 2 | 141 | 48 | 265 | 1.95 |
| 15 | 111 | 1 | 0 | 69 | 129 | 310 | 2.28 |
| 16 | 51 | 0 | 3 | 91 | 58 | 203 | 1.49 |
| 17 | 14 | 12 | 0 | 691 | 279 | 996 | 7.32 |
| 18 | 1 | 4 | 67 | 294 | 87 | 453 | 3.33 |
| 19 | 0 | 1 | 0 | 8 | 3 | 12 | 0.09 |
| 20 | 0 | 1 | 5 | 2 | 0 | 8 | 0.06 |
| 21 | 0 | 0 | 4 | 6 | 3 | 13 | 0.1 |
| 22 | 0 | 0 | 5 | 0 | 0 | 5 | 0.04 |
| 23 | 0 | 0 | 1 | 0 | 0 | 1 | 0.01 |
| TOTAL | 1148 | 322 | 166 | 8174 | 3802 | 13,612 |
| Month | Station A | Station B | Station C | Station D | Station E | Total | % |
|---|---|---|---|---|---|---|---|
| June 2015 | 0 | 5 | 0 | 1 | 3 | 9 | 1.83 |
| July 2015 | 0 | 13 | 10 | 11 | 10 | 44 | 8.96 |
| August 2015 | 0 | 13 | 6 | 4 | 1 | 24 | 4.89 |
| September 2015 | 0 | 4 | 12 | 0 | 2 | 18 | 3.67 |
| October 2015 | 0 | 4 | 11 | 8 | 3 | 26 | 5.3 |
| November 2015 | 10 | 8 | 11 | 1 | 1 | 31 | 6.31 |
| December 2015 | 3 | 31 | 19 | 5 | 1 | 59 | 12.02 |
| January 2016 | 3 | 7 | 14 | 20 | 0 | 44 | 8.96 |
| February 2016 | 3 | 5 | 3 | 20 | 3 | 31 | 6.31 |
| March 2016 | 6 | 0 | 53 | 4 | 11 | 74 | 15.07 |
| April 2016 | 7 | 0 | 20 | 3 | 29 | 59 | 12.02 |
| May 2016 | 3 | 28 | 6 | 7 | 28 | 72 | 14.66 |
| TOTAL | 35 | 115 | 165 | 84 | 92 | 491 |
| Month | Station A | Station B | Station C | Station D | Station E | Total | % |
|---|---|---|---|---|---|---|---|
| June 2015 | 0 | 24 | 20 | 1032 | 349 | 1425 | 10.47 |
| July 2015 | 0 | 33 | 334 | 612 | 269 | 948 | 6.96 |
| August 2015 | 0 | 30 | 23 | 642 | 299 | 994 | 7.3 |
| September 2015 | 0 | 68 | 25 | 407 | 148 | 648 | 4.76 |
| October 2015 | 29 | 10 | 35 | 590 | 298 | 962 | 7.07 |
| November 2015 | 46 | 6 | 14 | 442 | 464 | 972 | 7.14 |
| December 2015 | 235 | 9 | 3 | 738 | 221 | 1206 | 8.86 |
| January 2016 | 142 | 11 | 6 | 412 | 299 | 870 | 6.39 |
| February 2016 | 70 | 3 | 1 | 477 | 206 | 757 | 5.56 |
| March 2016 | 53 | 67 | 1 | 726 | 386 | 1233 | 9.06 |
| April 2016 | 79 | 32 | 3 | 796 | 210 | 1120 | 8.23 |
| May 2016 | 494 | 29 | 1 | 1300 | 653 | 2477 | 18.2 |
| TOTAL | 1148 | 322 | 166 | 8174 | 3802 | 13,612 |
References
- Gibb, R.; Browning, E.; Glover-Kapfer, P.; Jones, K.E. Emerging opportunities and challenges for passive acoustics in ecological assessment and monitoring. Methods Ecol. Evol. 2018, 10, 169–185. [Google Scholar] [CrossRef]
- Wang, Q.; Song, Y.; Du, Y.; Yang, Z.; Cui, P.; Luo, B. Hierarchical-taxonomy-aware and attentional convolutional neural networks for acoustic identification of bird species: A phylogenetic perspective. Ecol. Inform. 2024, 80, 102538. [Google Scholar] [CrossRef]
- Zeng, Y.; Wang, X.; Liu, J.; Cao, J.; Sun, Y.; Zhao, S.; Chen, Z.; Kim, J.K.; Zhang, J.; He, P. Harnessing the power of eDNA technology for macroalgal ecological studies: Recent advances, challenges, and future perspectives. Algal Res. 2024, 77, 103340. [Google Scholar] [CrossRef]
- Mata, A.; Moffat, D.; Almeida, S.; Radeta, M.; Jay, W.; Mortimer, N.; Awty-Carroll, K.; Thomas, O.R.; Brotas, V.; Groom, S. Drone imagery and deep learning for mapping the density of wild Pacific oysters to manage their expansion into protected areas. Ecol. Inform. 2024, 82, 102708. [Google Scholar] [CrossRef]
- Wang, B.; Li, R.; Lan, X.; Kong, D.; Liu, X.; Xie, S. Benthic diatom eDNA metabarcoding for ecological assessment of an urban river: A comparison with morphological method. Ecol. Indic. 2024, 166, 112302. [Google Scholar] [CrossRef]
- Digby, A.; Towsey, M.; Bell, B.D.; Teal, P.D. A practical comparison of manual and autonomous methods for acoustic monitoring. Methods Ecol. Evol. 2013, 4, 675–683. [Google Scholar] [CrossRef]
- Sugai, L.S.M.; Silva, T.S.F.; Ribeiro, J.W.; Llusia, D. Terrestrial Passive Acoustic Monitoring: Review and Perspectives. BioScience 2018, 69, 15–25. [Google Scholar] [CrossRef]
- Desjonquères, C.; Gifford, T.; Linke, S. Passive acoustic monitoring as a potential tool to survey animal and ecosystem processes in freshwater environments. Freshw. Biol. 2019, 65, 7–19. [Google Scholar] [CrossRef]
- Hill, A.P.; Prince, P.; Snaddon, J.L.; Doncaster, C.P.; Rogers, A. AudioMoth: A low-cost acoustic device for monitoring biodiversity and the environment. HardwareX 2019, 6, e00073. [Google Scholar] [CrossRef]
- Manzano-Rubio, R.; Bota, G.; Brotons, L.; Soto-Largo, E.; Pérez-Granados, C. Low-cost open-source recorders and ready-to-use machine learning approaches provide effective monitoring of threatened species. Ecol. Inform. 2022, 72, 101910. [Google Scholar] [CrossRef]
- Kahl, S.; Wood, C.M.; Eibl, M.; Klinck, H. BirdNET: A deep learning solution for avian diversity monitoring. Ecol. Inform. 2021, 61, 101236. [Google Scholar] [CrossRef]
- Bota, G.; Manzano-Rubio, R.; Catalán, L.; Gómez-Catasús, J.; Pérez-Granados, C. Hearing to the Unseen: AudioMoth and BirdNET as a Cheap and Easy Method for Monitoring Cryptic Bird Species. Sensors 2023, 23, 7176. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Granados, C.; Schuchmann, K.; Marques, M.I. Vocal behavior of the Undulated Tinamou (Crypturellus undulatus) over an annual cycle in the Brazilian Pantanal: New ecological information. Biotropica 2019, 52, 165–171. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.-L.; Ramoni-Perazzi, P.; Marques, M.I. Calling behaviour of Elachistocleis matogrosso (Anura, Microhylidae) is associated with habitat temperature and rainfall. Bioacoustics 2019, 29, 670–683. [Google Scholar] [CrossRef]
- Pérez-Granados, C. BirdNET: Applications, performance, pitfalls and future opportunities. Ibis 2023, 165, 1068–1075. [Google Scholar] [CrossRef]
- Wood, C.M.; Cruz, A.B.; Kahl, S. Pairing a user-friendly machine-learning animal sound detector with passive acoustic surveys for occupancy modeling of an endangered primate. Am. J. Primatol. 2023, 85, e23507. [Google Scholar] [CrossRef]
- Wood, C.M.; Kahl, S.; Barnes, S.; Van Horne, R.; Brown, C. Passive acoustic surveys and the BirdNET algorithm reveal detailed spatiotemporal variation in the vocal activity of two anurans. Bioacoustics 2023, 32, 532–543. [Google Scholar] [CrossRef]
- Bota, G.; Manzano-Rubio, R.; Fanlo, H.; Franch, N.; Brotons, L.; Villero, D.; Devisscher, S.; Pavesi, A.; Cavaletti, E.; Pérez-Granados, C. Passive acoustic monitoring and automated detection of the American bullfrog. Biol. Invasions 2024, 26, 1269–1279. [Google Scholar] [CrossRef]
- Znidersic, E.; Towsey, M.; Roy, W.; Darling, S.E.; Truskinger, A.; Roe, P.; Watson, D.M. Using visualization and machine learning methods to monitor low detectability species—The least bittern as a case study. Ecol. Inform. 2019, 55, 101014. [Google Scholar] [CrossRef]
- Gill, S.A.; Vonhof, M.J.; Stutchbury, B.J.M.; Morton, E.S.; Quinn, J.S. No evidence for acoustic mate-guarding in duetting buff-breasted wrens (Thryothorus leucotis). Behav. Ecol. Sociobiol. 2005, 57, 557–565. [Google Scholar] [CrossRef]
- De Nóbrega, P.F.A.; De Pinho, J.B. Biologia reprodutiva e uso de habitat por Cantorchilus leucotis (Lafresnaye, 1845) (Aves, Troglodytidae) no Pantanal, Mato Grosso, Brasil. Papéis Avulsos Zool. 2010, 50, 511–516. [Google Scholar] [CrossRef]
- Gill, S.A. Buff-breasted Wren (Cantorchilus leucotis). In Birds of the World; Schulenberg, T.S., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Kroodsma, D.E.; Brewer, D. Moustached Wren (Pheugopedius genibarbis). In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Mann, N.I.; Dingess, K.A.; Barker, F.K.; Graves, J.A.; Slater, P.J. A comparative study of song form and duetting in Neotropical Thryothorus wrens. Behaviour 2009, 146, 1–43. [Google Scholar]
- Heinonen Fortabat, S.; Gil, G.; Marino, G. Sobre las aves del Parque Nacional Río Pilcomayo con la adición de Basileuterus flaveolus a la avifauna argentina. El Hornero 1995, 14. Available online: https://bibliotecadigital.exactas.uba.ar/download/hornero/hornero_v014_n01y02_p069.pdf (accessed on 12 August 2024). [CrossRef]
- Kroodsma, D.; Brewer, D.; Kirwan, G.M. Thrush-like Wren (Campylorhynchus turdinus). In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Junk, W.J.; da Cunha, C.N.; Wantzen, K.M.; Petermann, P.; Strüssmann, C.; Marques, M.I.; Adis, J. Biodiversity and its conservation in the Pantanal of Mato Grosso, Brazil. Aquat. Sci. 2006, 68, 278–309. [Google Scholar] [CrossRef]
- Pott, A.; Pott, V.J. Features and conservation of the Brazilian Pantanal wetland. Wetl. Ecol. Manag. 2004, 12, 547–552. [Google Scholar] [CrossRef]
- Orfeo, O.; Stevaux, J. Hydraulic and morphological characteristics of middle and upper reaches of the Paraná River (Argentina and Brazil). Geomorphology 2002, 44, 309–322. [Google Scholar] [CrossRef]
- Ferreira, V.G. Contribution to the Taxonomy and Ecology of Strandesia sl (Crustacea, Ostracoda, Cypricercinae) from Brazilian Floodplains. Master Thesis, Maringá State University, Maringá, Brazil, 2019. Available online: http://hdl.handle.net/1834/14992 (accessed on 12 August 2024).
- Alho, C.J.R.; Silva, J.S.V. Effects of Severe Floods and Droughts on Wildlife of the Pantanal Wetland (Brazil)—A Review. Animals 2012, 2, 591–610. [Google Scholar] [CrossRef]
- Hamilton, S.K.; Sippel, S.J.; Melack, J.M. Inundation patterns in the Pantanal wetland of South America determined from passive microwave remote sensing. Arch. Für Hydrobiol. 1996, 137, 1–23. [Google Scholar] [CrossRef]
- Fraser, L.H.; Keddy, P.A. The World’s Largest Wetlands: Ecology and Conservation; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar] [CrossRef]
- Wood, C.M.; Kahl, S. Guidelines for appropriate use of BirdNET scores and other detector outputs. J. Ornithol. 2024, 165, 777–782. [Google Scholar] [CrossRef]
- Knight, E.C.; Hannah, K.C.; Foley, G.J.; Scott, C.D.; Brigham, R.M.; Bayne, E. Recommendations for acoustic recognizer performance assessment with application to five common automated signal recognition programs. Avian Conserv. Ecol. 2017, 12, 14. [Google Scholar] [CrossRef]
- Thompson, D.E. An Overview of Audacity. Gen. Music. Today 2014, 27, 40–43. [Google Scholar] [CrossRef]
- Wood, C.M.; Günther, F.; Rex, A.; Hofstadter, D.F.; Reers, H.; Kahl, S.; Peery, M.Z.; Klinck, H. Real-time acoustic monitoring facilitates the proactive management of biological invasions. Biol. Invasions 2024, 1–8. [Google Scholar] [CrossRef]
- Wood, C.M.; Kahl, S.; Chaon, P.; Peery, M.Z.; Klinck, H. Survey coverage, recording duration and community composition affect observed species richness in passive acoustic surveys. Methods Ecol. Evol. 2021, 12, 885–896. [Google Scholar] [CrossRef]
- Sethi, S.S.; Fossøy, F.; Cretois, B.; Rosten, C.M. Management Relevant Applications of Acoustic Monitoring for Norwegian Nature–The Sound of Norway; (Report 2064); Norwegian Institute for Nature Research: Trondheim, Sweden, 2021; Available online: https://hdl.handle.net/11250/2832294 (accessed on 12 August 2024).
- Kahl, S. Identifying Birds by Sound: Large-Scale Acoustic Event Recognition for Avian Activity Monitoring. Master Thesis, Chemnitz University of Technology, Chemnitz, Germany, 2020. Available online: https://nbn-resolving.org/urn:nbn:de:bsz:ch1-qucosa2-369869 (accessed on 12 August 2024).
- Cole, J.S.; Michel, N.L.; A Emerson, S.; Siegel, R.B. Automated bird sound classifications of long-duration recordings produce occupancy model outputs similar to manually annotated data. Ornithol. Appl. 2022, 124, duac003. [Google Scholar] [CrossRef]
- Ventura, T.M.; Ganchev, T.D.; Pérez-Granados, C.; de Oliveira, A.G.; Pedroso, G.d.S.G.; Marques, M.I.; Schuchmann, K.-L. The importance of acoustic background modelling in CNN-based detection of the neotropical White-lored Spinetail (Aves, Passeriformes, Furnaridae). Bioacoustics 2024, 33, 103–121. [Google Scholar] [CrossRef]
- Gil, D.; Llusia, D. The Bird Dawn Chorus Revisited. In Coding Strategies in Vertebrate Acoustic Communication; Aubin, T., Mathevon, N., Eds.; Springer: Cham, Switzerland, 2020; Volume 7, pp. 45–90. [Google Scholar] [CrossRef]
- Bradley, D.W.; Mennill, D.J. Solos, duets and choruses: Vocal behaviour of the Rufous-naped Wren (Campylorhynchus rufinucha), a cooperatively breeding neotropical songbird. J. Ornithol. 2009, 150, 743–753. [Google Scholar] [CrossRef]
- Baldo, S.; Mennill, D.J. Vocal behavior of Great Curassows, a vulnerable Neotropical bird. J. Field Ornithol. 2011, 82, 249–258. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.-L. Diel and Seasonal Variations of Vocal Behavior of the Neotropical White-Tipped Dove (Leptotila verreauxi). Diversity 2020, 12, 402. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K. Nocturnal vocal behaviour of the diurnal Undulated Tinamou (Crypturellus undulatus) is associated with temperature and moon phase. Ibis 2021, 163, 684–694. [Google Scholar] [CrossRef]
- Winiarska, D.; Pérez-Granados, C.; Budka, M.; Osiejuk, T.S. Year-round vocal activity of two African barbet species. Emu-Austral Ornithol. 2024, 1–11. [Google Scholar] [CrossRef]
- Winiarska, D.; Pérez-Granados, C.; Budka, M.; Osiejuk, T.S. Passive acoustic monitoring of endangered endemic Afromontane tropical species: A case study with two turacos. Afr. J. Ecol. 2024, 62, 213280. [Google Scholar] [CrossRef]
- BirdLife International. Campylorhynchus turdinus. The IUCN Red List of Threatened Species. 2018. Available online: https://www.iucnredlist.org/species/22711306/131962920 (accessed on 12 August 2024).
- BirdLife International. Cantorchilus leucotis. The IUCN Red List of Threatened Species. 2021. Available online: https://www.iucnredlist.org/species/22711467/166912898 (accessed on 12 August 2024).
- Alho, C. Biodiversity of the Pantanal: Response to seasonal flooding regime and to environmental degradation. Braz. J. Biol. 2008, 68, 957–966. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.-L. Illuminating the nightlife of two Neotropical nightjars: Vocal behavior over a year and monitoring recommendations. Ethol. Ecol. Evol. 2020, 32, 466–480. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.-L. Passive Acoustic Monitoring of Chaco Chachalaca (Ortalis canicollis) Over a Year: Vocal Activity Pattern and Monitoring Recommendations. Trop. Conserv. Sci. 2021, 14, 19400829211058295. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K. Diel and seasonal variation of Striped Cuckoo (Tapera naevia) vocalizations revealed using automated signal recognition. Ibis 2022, 165, 179–189. [Google Scholar] [CrossRef]
- De Deus, F.F.; Schuchmann, K.-L.; Marques, M.I. Seasonality in the Brazilian Pantanal influences avian functional diversity. Stud. Neotropical Fauna Environ. 2020, 57, 187–197. [Google Scholar] [CrossRef]
- Koller, W.W.; de Barros, A.T.M.; Corrêa, E.C. Abundance and seasonality of Cochliomyia macellaria (Diptera: Calliphoridae) in Southern Pantanal, Brazil. Rev. Bras. De Parasitol. Veter. 2011, 20, 27–30. [Google Scholar] [CrossRef]
- Soares, S.d.A.; Suarez, Y.R.; Fernandes, W.D.; Tenório, P.M.S.; Delabie, J.H.C.; Antonialli-Junior, W.F. Temporal variation in the composition of ant assemblages (Hymenoptera, Formicidae) on trees in the Pantanal floodplain, Mato Grosso do Sul, Brazil. Rev. Bras. de Èntomol. 2013, 57, 84–90. [Google Scholar] [CrossRef]
- Lopes, L.E.; Fernandes, A.M.; Marini, M.Â. Diet of some Atlantic Forest birds. Ararajuba 2005, 13, 95–103. [Google Scholar]
- De Pinho, J.B.; Aragona, M.; Hakamada, K.Y.P.; Marini, M. Migration patterns and seasonal forest use by birds in the Brazilian Pantanal. Bird Conserv. Int. 2017, 27, 371–387. [Google Scholar] [CrossRef]
- De Deus, F.F.; Schuchmann, K.-L.; Arieira, J.; Tissiani, A.S.d.O.; Marques, M.I. Avian Beta Diversity in a Neotropical Wetland: The Effects of Flooding and Vegetation Structure. Wetlands 2020, 40, 1513–1527. [Google Scholar] [CrossRef]
- Brumm, H.; Slabbekoorn, H. Acoustic communication in noise. Adv. Study Behav. 2005, 35, 151–209. [Google Scholar] [CrossRef]
- Mennill, D.J. Variation in the Vocal Behavior of Common Loons (Gavia immer): Insights from Landscape-level Recordings. Waterbirds 2014, 37, 26–36. [Google Scholar] [CrossRef]
- Robbins, C.S. Effect of time of day on bird activity. Stud. Avian Biol. 1981, 6, 275–286. [Google Scholar]
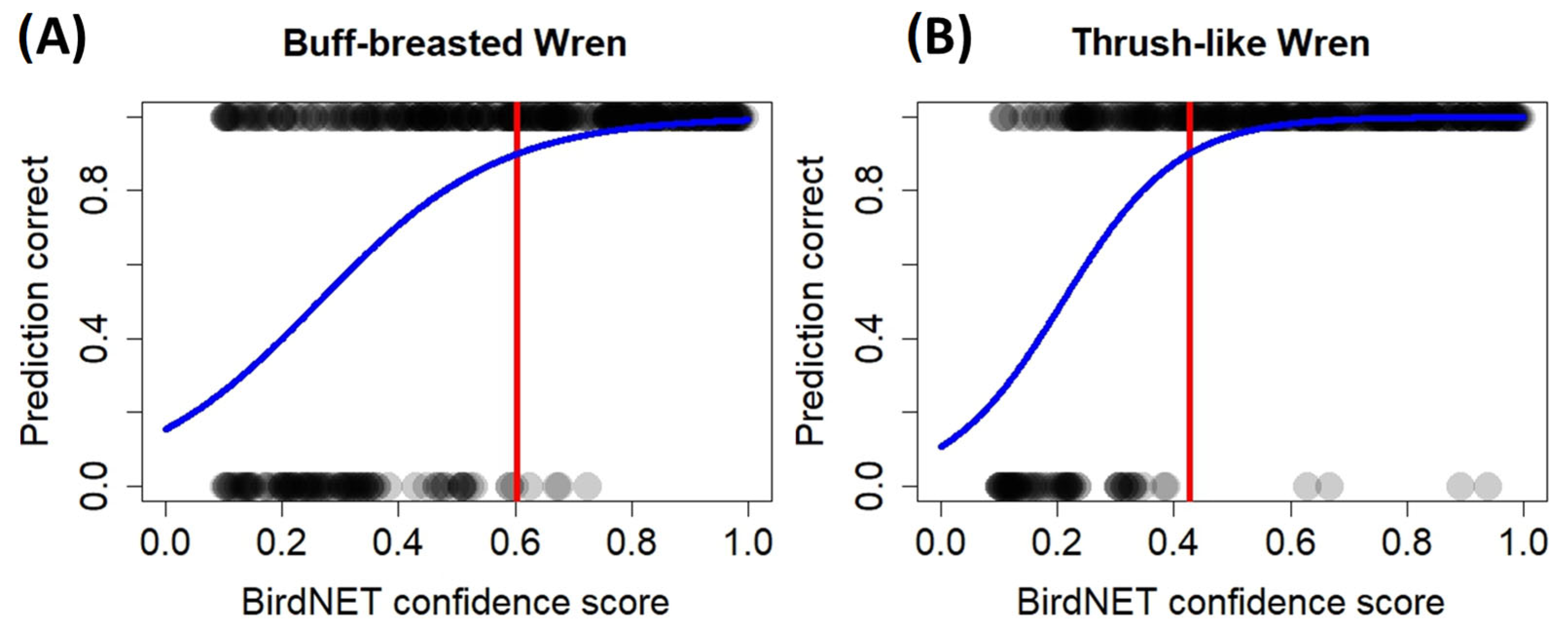
| Species | Predictions | 0.1–0.3 | 0.3–0.5 | 0.5–0.7 | >0.7 | 0.1–1 |
|---|---|---|---|---|---|---|
| Moustached Wren | Correct | 60 | 66 | 87 | 113 | 326 |
| Precision | 60% | 66% | 87% | 75.3% | 72.4% | |
| Buff-breasted Wren | Correct | 42 | 66 | 87 | 149 | 344 |
| Precision | 42% | 66% | 87% | 99.3% | 76.4% | |
| Thrush-like Wren | Correct | 47 | 85 | 98 | 148 | 378 |
| Precision | 47% | 85% | 98% | 98.7% | 84% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amorós-Ausina, D.; Schuchmann, K.-L.; Marques, M.I.; Pérez-Granados, C. Living Together, Singing Together: Revealing Similar Patterns of Vocal Activity in Two Tropical Songbirds Applying BirdNET. Sensors 2024, 24, 5780. https://doi.org/10.3390/s24175780
Amorós-Ausina D, Schuchmann K-L, Marques MI, Pérez-Granados C. Living Together, Singing Together: Revealing Similar Patterns of Vocal Activity in Two Tropical Songbirds Applying BirdNET. Sensors. 2024; 24(17):5780. https://doi.org/10.3390/s24175780
Chicago/Turabian StyleAmorós-Ausina, David, Karl-L. Schuchmann, Marinez I. Marques, and Cristian Pérez-Granados. 2024. "Living Together, Singing Together: Revealing Similar Patterns of Vocal Activity in Two Tropical Songbirds Applying BirdNET" Sensors 24, no. 17: 5780. https://doi.org/10.3390/s24175780