
Near-InfraRed Spectroscopy Provides a Reproducible Estimate of Muscle Aerobic Capacity, but Not Whole-Body Aerobic Power

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Organization
2.3. Anthropometric and Body Composition Measurements
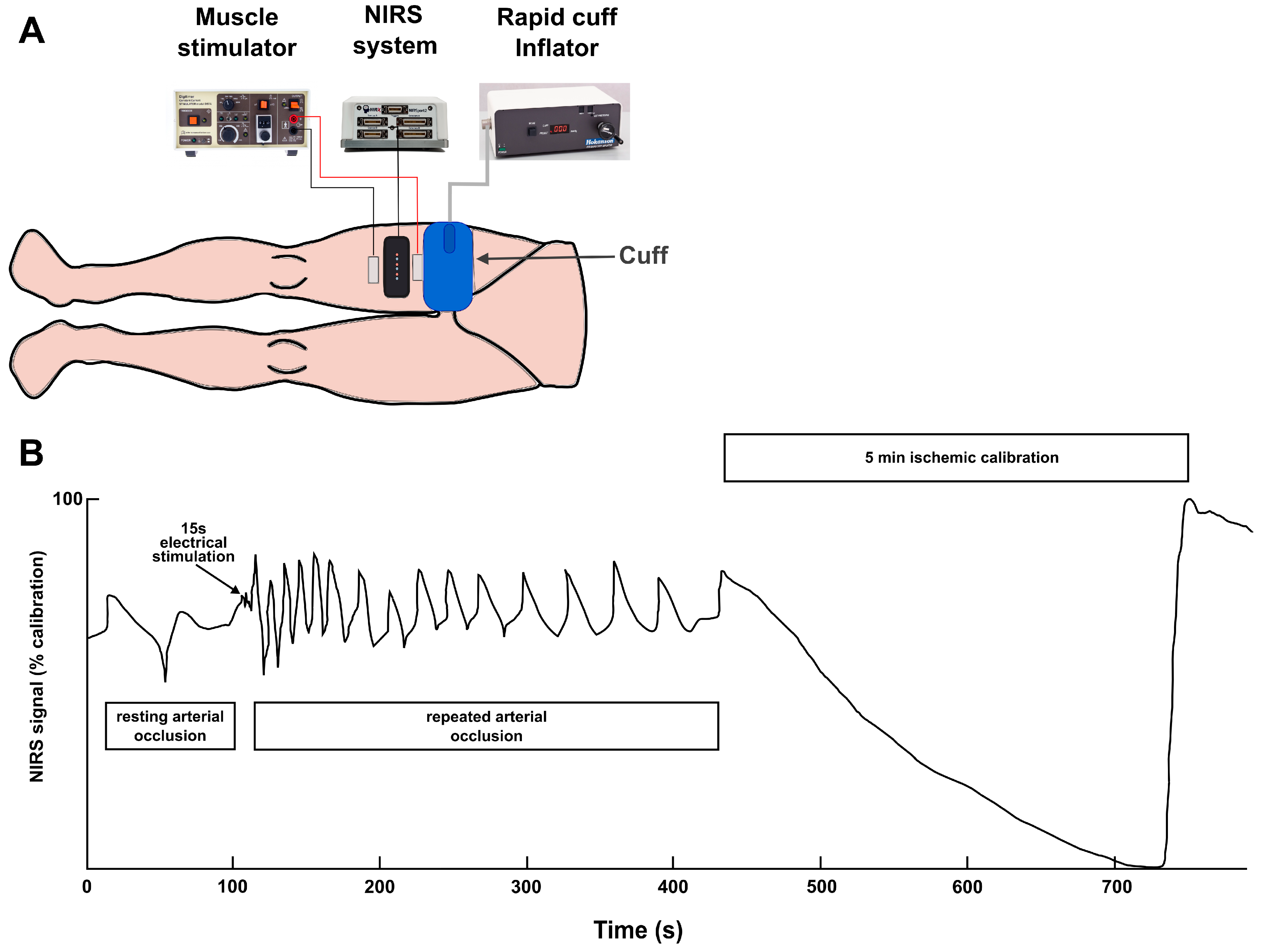
2.4. Measurement of Mitochondrial Respiratory Capacity (NIRS Procedure)
2.5. Whole-Body Aerobic Capacity (O2 Peak) Test
2.6. Statistical Analysis
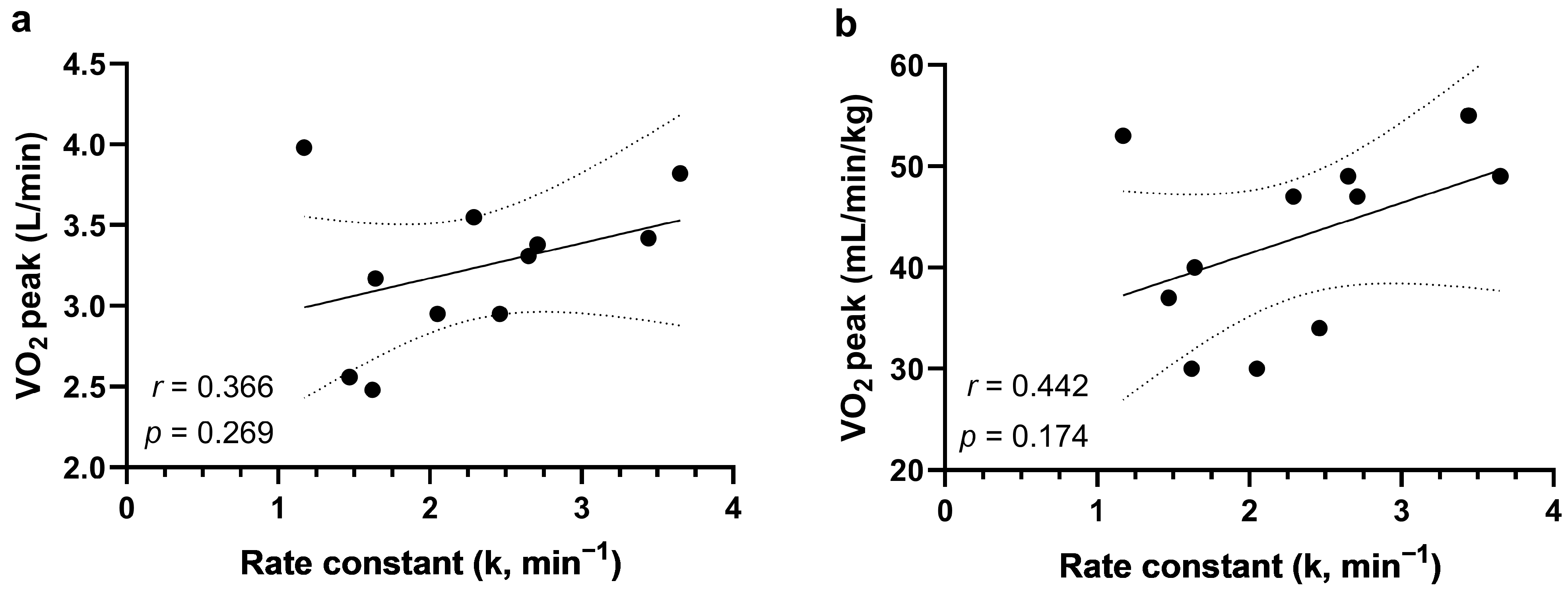
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strasser, B.; Burtscher, M. Survival of the Fittest: O2Max, a Key Predictor of Longevity. Front. Biosci. 2018, 23, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Solomon, A.; Borodulin, K.; Ngandu, T.; Kivipelto, M.; Laatikainen, T.; Kulmala, J. Self-Rated Physical Fitness and Estimated Maximal Oxygen Uptake in Relation to All-Cause and Cause-Specific Mortality. Scand. J. Med. Sci. Sports 2018, 28, 532–540. [Google Scholar] [CrossRef]
- Joyner, M.J.; Dominelli, P.B. Central Cardiovascular System Limits to Aerobic Capacity. Exp. Physiol. 2021, 106, 2299–2303. [Google Scholar] [CrossRef]
- van der Zwaard, S.; Brocherie, F.; Jaspers, R.T. Under the Hood: Skeletal Muscle Determinants of Endurance Performance. Front. Sports Act. Living 2021, 3, 719434. [Google Scholar] [CrossRef]
- Skattebo, Ø.; Calbet, J.A.L.; Rud, B.; Capelli, C.; Hallén, J. Contribution of Oxygen Extraction Fraction to Maximal Oxygen Uptake in Healthy Young Men. Acta Physiol. 2020, 230, e13486. [Google Scholar] [CrossRef]
- Wagner, P.D. Determinants of Maximal Oxygen Consumption. J. Muscle Res. Cell Motil. 2023, 44, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Cartee, G.D.; Hepple, R.T.; Bamman, M.M.; Zierath, J.R. Exercise Promotes Healthy Aging of Skeletal Muscle. Cell Metab. 2016, 23, 1034–1047. [Google Scholar] [CrossRef] [PubMed]
- Hamaoka, T.; Iwane, H.; Shimomitsu, T.; Katsumura, T.; Murase, N.; Nishio, S.; Osada, T.; Kurosawa, Y.; Chance, B. Noninvasive Measures of Oxidative Metabolism on Working Human Muscles by Near-Infrared Spectroscopy. J. Appl. Physiol. 1996, 81, 1410–1417. [Google Scholar] [CrossRef]
- Ryan, T.E.; Brophy, P.; Lin, C.-T.; Hickner, R.C.; Neufer, P.D. Assessment of in Vivo Skeletal Muscle Mitochondrial Respiratory Capacity in Humans by Near-Infrared Spectroscopy: A Comparison with in Situ Measurements. J. Physiol. 2014, 592, 3231–3241. [Google Scholar] [CrossRef]
- Ryan, T.E.; Erickson, M.L.; Brizendine, J.T.; Young, H.-J.; McCully, K.K. Noninvasive Evaluation of Skeletal Muscle Mitochondrial Capacity with Near-Infrared Spectroscopy: Correcting for Blood Volume Changes. J. Appl. Physiol. 2012, 113, 175–183. [Google Scholar] [CrossRef]
- Ryan, T.E.; Southern, W.M.; Reynolds, M.A.; McCully, K.K. A Cross-Validation of near-Infrared Spectroscopy Measurements of Skeletal Muscle Oxidative Capacity with Phosphorus Magnetic Resonance Spectroscopy. J. Appl. Physiol. 2013, 115, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- de Aguiar, R.A.; Turnes, T.; Borszcz, F.K.; Raimundo, J.A.G.; Caputo, F. Near-Infrared Spectroscopy-Derived Muscle o2 Kinetics after Moderate Running Exercise in Healthy Males: Reliability and Associations with Parameters of Aerobic Fitness. Exp. Physiol. 2022, 107, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Lagerwaard, B.; Keijer, J.; McCully, K.K.; de Boer, V.C.J.; Nieuwenhuizen, A.G. In Vivo Assessment of Muscle Mitochondrial Function in Healthy, Young Males in Relation to Parameters of Aerobic Fitness. Eur. J. Appl. Physiol. 2019, 119, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Beever, A.T.; Tripp, T.R.; Zhang, J.; MacInnis, M.J. NIRS-Derived Skeletal Muscle Oxidative Capacity Is Correlated with Aerobic Fitness and Independent of Sex. J. Appl. Physiol. 2020, 129, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Guzman, S.; Ramirez, J.; Keslacy, S.; de Leon, R.; Yamazaki, K.; Dy, C. Association between Muscle Aerobic Capacity and Whole-Body Peak Oxygen Uptake. Eur. J. Appl. Physiol. 2020, 120, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Lagerwaard, B.; Nieuwenhuizen, A.G.; de Boer, V.C.J.; Keijer, J. In Vivo Assessment of Mitochondrial Capacity Using NIRS in Locomotor Muscles of Young and Elderly Males with Similar Physical Activity Levels. GeroScience 2020, 42, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Lagerwaard, B.; Janssen, J.J.E.; Cuijpers, I.; Keijer, J.; de Boer, V.C.J.; Nieuwenhuizen, A.G. Muscle Mitochondrial Capacity in High- and Low-Fitness Females Using near-Infrared Spectroscopy. Physiol. Rep. 2021, 9, e14838. [Google Scholar] [CrossRef]
- Pelka, E.Z.; Davis, B.R.; Tomko, P.M.; McDaniel, J. Analysis of Electrical Stimulation and Voluntary Muscle Contraction on Skeletal Muscle Oxygen Uptake and Mitochondrial Recovery Using Near-Infrared Spectroscopy. Eur. J. Appl. Physiol. 2023, 123, 2053–2061. [Google Scholar] [CrossRef] [PubMed]
- Yoshiko, A.; Shiozawa, K.; Niwa, S.; Takahashi, H.; Koike, T.; Watanabe, K.; Katayama, K.; Akima, H. Association of Skeletal Muscle Oxidative Capacity with Muscle Function, Sarcopenia-Related Exercise Performance, and Intramuscular Adipose Tissue in Older Adults. Geroscience 2023, 46, 2715–2727. [Google Scholar] [CrossRef]
- Hanna, R.; Gosalia, J.; Demalis, A.; Hobson, Z.; McCully, K.K.; Irving, B.A.; Mookerjee, S.; Vairo, G.L.; Proctor, D.N. Bilateral NIRS Measurements of Muscle Mitochondrial Capacity: Feasibility and Repeatability. Physiol. Rep. 2021, 9, e14826. [Google Scholar] [CrossRef]
- Lai, R.E.; Holman, M.E.; Chen, Q.; Rivers, J.; Lesnefsky, E.J.; Gorgey, A.S. Assessment of Mitochondrial Respiratory Capacity Using Minimally Invasive and Noninvasive Techniques in Persons with Spinal Cord Injury. PLoS ONE 2022, 17, e0265141. [Google Scholar] [CrossRef] [PubMed]
- Batterson, P.M.; Norton, M.R.; Hetz, S.E.; Rohilla, S.; Lindsay, K.G.; Subudhi, A.W.; Jacobs, R.A. Improving Biologic Predictors of Cycling Endurance Performance with Near-Infrared Spectroscopy Derived Measures of Skeletal Muscle Respiration: E Pluribus Unum. Physiol. Rep. 2020, 8, e14342. [Google Scholar] [CrossRef] [PubMed]
- Fennell, C.R.J.; Mauger, A.R.; Hopker, J.G. Reproducibility of NIRS-Derived Mitochondrial Oxidative Capacity in Highly Active Older Adults. Exp. Gerontol. 2023, 175, 112156. [Google Scholar] [CrossRef] [PubMed]
- Adami, A.; Cao, R.; Porszasz, J.; Casaburi, R.; Rossiter, H.B. Reproducibility of NIRS Assessment of Muscle Oxidative Capacity in Smokers with and without COPD. Respir. Physiol. Neurobiol. 2017, 235, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Adami, A.; Rossiter, H.B. Principles, Insights, and Potential Pitfalls of the Noninvasive Determination of Muscle Oxidative Capacity by near-Infrared Spectroscopy. J. Appl. Physiol. 2018, 124, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Zuccarelli, L.; do Nascimento Salvador, P.C.; Del Torto, A.; Fiorentino, R.; Grassi, B. Skeletal Muscle o2 Kinetics by the NIRS Repeated Occlusions Method during the Recovery from Cycle Ergometer Exercise. J. Appl. Physiol. 2020, 128, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Koutlas, A.; Smilios, I.; Kokkinou, E.M.; Myrkos, A.; Kounoupis, A.; Dipla, K.; Zafeiridis, A. NIRS-Derived Muscle-Deoxygenation and Microvascular Reactivity During Occlusion–Reperfusion at Rest Are Associated With Whole-Body Aerobic Fitness. Res. Q. Exerc. Sport 2023, 95, 127–139. [Google Scholar] [CrossRef]
- Pilotto, A.M.; Adami, A.; Mazzolari, R.; Brocca, L.; Crea, E.; Zuccarelli, L.; Pellegrino, M.A.; Bottinelli, R.; Grassi, B.; Rossiter, H.B.; et al. Near-Infrared Spectroscopy Estimation of Combined Skeletal Muscle Oxidative Capacity and O2 Diffusion Capacity in Humans. J. Physiol. 2022, 600, 4153–4168. [Google Scholar] [CrossRef]
- Juškevičiūtė, E.; Neuberger, E.; Eimantas, N.; Venckunas, T.; Kamandulis, S.; Simon, P.; Brazaitis, M. Three-Week Sprint Interval Training (SIT) Reduces Cell-Free DNA and Low-Frequency Fatigue but Does Not Induce VO2max Improvement in Older Men. Eur. J. Appl. Physiol. 2023, 124, 1297–1309. [Google Scholar] [CrossRef]
- Ballesteros-Pomar, M.D.; Calleja-Fernández, A.; Diez-Rodríguez, R.; Vidal-Casariego, A.; Blanco-Suárez, M.D.; Cano-Rodríguez, I. Comparison of Different Body Composition Measurements in Severely Obese Patients in the Clinical Setting. Nutr. Hosp. 2012, 27, 1626–1630. [Google Scholar] [CrossRef]
- Brizendine, J.T.; Ryan, T.E.; Larson, R.D.; McCully, K.K. Skeletal Muscle Metabolism in Endurance Athletes with Near-Infrared Spectroscopy. Med. Sci. Sports Exerc. 2013, 45, 869–875. [Google Scholar] [CrossRef]
- McCully, K.K.; Iotti, S.; Kendrick, K.; Wang, Z.; Posner, J.D.; Leigh, J.; Chance, B. Simultaneous in Vivo Measurements of HbO2 Saturation and PCr Kinetics after Exercise in Normal Humans. J. Appl. Physiol. 1994, 77, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.G.; Marshall, S.W.; Batterham, A.M.; Hanin, J. Progressive Statistics for Studies in Sports Medicine and Exercise Science. Med. Sci. Sports Exerc. 2009, 41, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, P.M.; Sabapathy, S.; Craven, J.; Arnold, B.; Minahan, C. Overreaching Attenuates Training-Induced Improvements in Muscle Oxidative Capacity. Med. Sci. Sports Exerc. 2020, 52, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, R.d.A.; Forot, J.; Millet, G.Y.; Murias, J.M. Comparing Muscle o2 from Near-Infrared Spectroscopy Desaturation Rate to Pulmonary o2 during Cycling below, at, and above the Maximal Lactate Steady State. J. Appl. Physiol. 2022, 132, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Katona, P.; Pilissy, T.; Tihanyi, A.; Laczkó, J. The Combined Effect of Cycling Cadence and Crank Resistance on Hamstrings and Quadriceps Muscle Activities during Cycling. Acta Physiol. Hung. 2014, 101, 505–516. [Google Scholar] [CrossRef]
- da Silva, J.C.L.; Tarassova, O.; Ekblom, M.M.; Andersson, E.; Rönquist, G.; Arndt, A. Quadriceps and Hamstring Muscle Activity during Cycling as Measured with Intramuscular Electromyography. Eur. J. Appl. Physiol. 2016, 116, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- McPhee, J.S.; Williams, A.G.; Perez-Schindler, J.; Degens, H.; Baar, K.; Jones, D.A. Variability in the Magnitude of Response of Metabolic Enzymes Reveals Patterns of Co-Ordinated Expression Following Endurance Training in Women. Exp. Physiol. 2011, 96, 699–707. [Google Scholar] [CrossRef]
- Johnson, M.A.; Polgar, J.; Weightman, D.; Appleton, D. Data on the Distribution of Fibre Types in Thirty-Six Human Muscles: An Autopsy Study. J. Neurol. Sci. 1973, 18, 111–129. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| PPO, W | 305.4 (32.7) |
| O2 peak, mL/kg/min | 44.7 (9.2) |
| HR peak, bpm | 180 (12) |
| VE, L/min | 125 (21) |
| RER | 1.19 (0.04) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venckunas, T.; Satas, A.; Brazaitis, M.; Eimantas, N.; Sipaviciene, S.; Kamandulis, S. Near-InfraRed Spectroscopy Provides a Reproducible Estimate of Muscle Aerobic Capacity, but Not Whole-Body Aerobic Power. Sensors 2024, 24, 2277. https://doi.org/10.3390/s24072277
Venckunas T, Satas A, Brazaitis M, Eimantas N, Sipaviciene S, Kamandulis S. Near-InfraRed Spectroscopy Provides a Reproducible Estimate of Muscle Aerobic Capacity, but Not Whole-Body Aerobic Power. Sensors. 2024; 24(7):2277. https://doi.org/10.3390/s24072277
Chicago/Turabian StyleVenckunas, Tomas, Andrius Satas, Marius Brazaitis, Nerijus Eimantas, Saule Sipaviciene, and Sigitas Kamandulis. 2024. "Near-InfraRed Spectroscopy Provides a Reproducible Estimate of Muscle Aerobic Capacity, but Not Whole-Body Aerobic Power" Sensors 24, no. 7: 2277. https://doi.org/10.3390/s24072277
APA StyleVenckunas, T., Satas, A., Brazaitis, M., Eimantas, N., Sipaviciene, S., & Kamandulis, S. (2024). Near-InfraRed Spectroscopy Provides a Reproducible Estimate of Muscle Aerobic Capacity, but Not Whole-Body Aerobic Power. Sensors, 24(7), 2277. https://doi.org/10.3390/s24072277