Neurochemical Changes and c-Fos Mapping in the Brain after Carisbamate Treatment of Rats Subjected to Lithium–Pilocarpine-Induced Status Epilepticus

 ,
,
Abstract
:1. Introduction
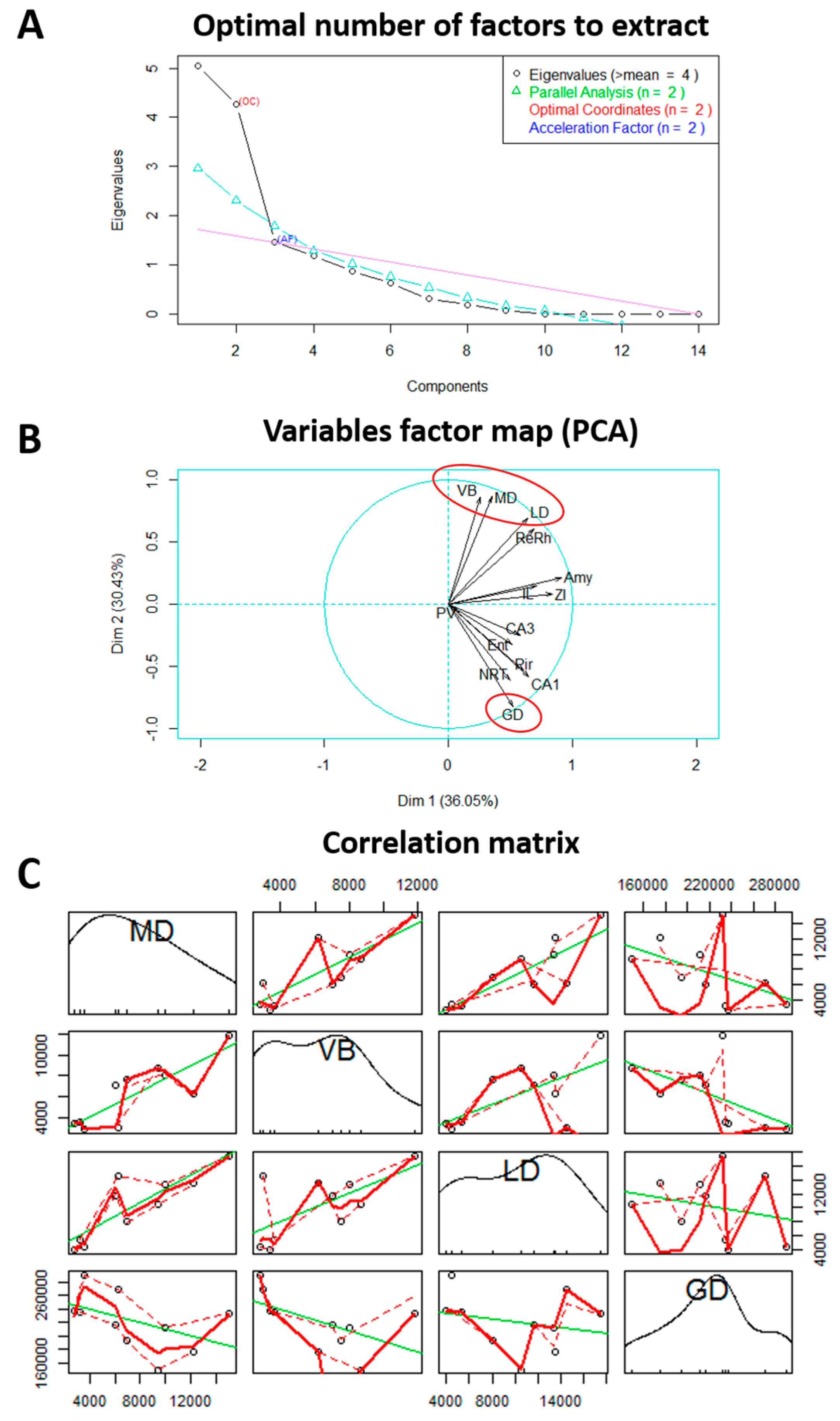
2. Results
2.1. Monoamine and Amino Acid Quantification
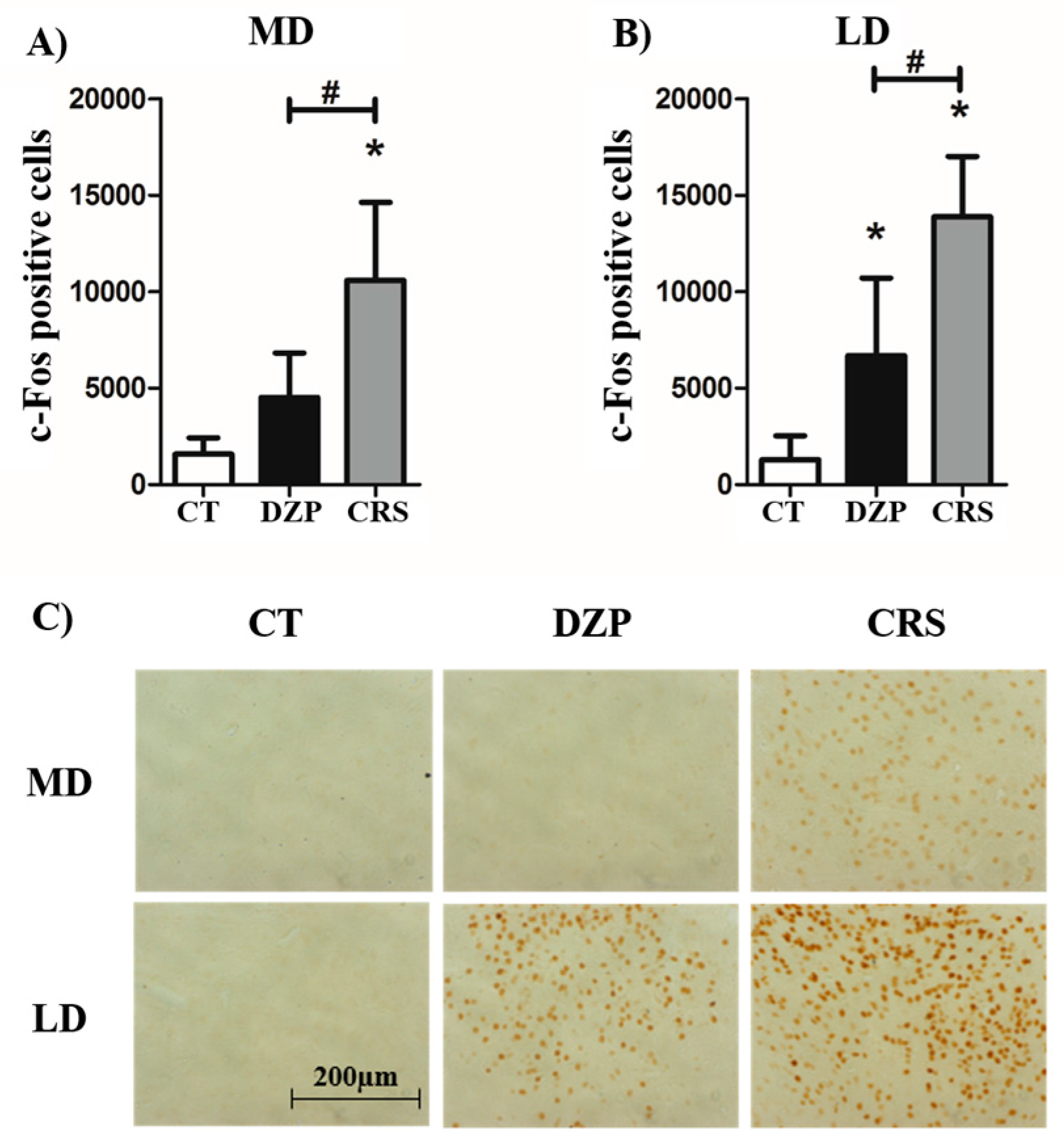
2.2. c-Fos Immunostaining
3. Discussion
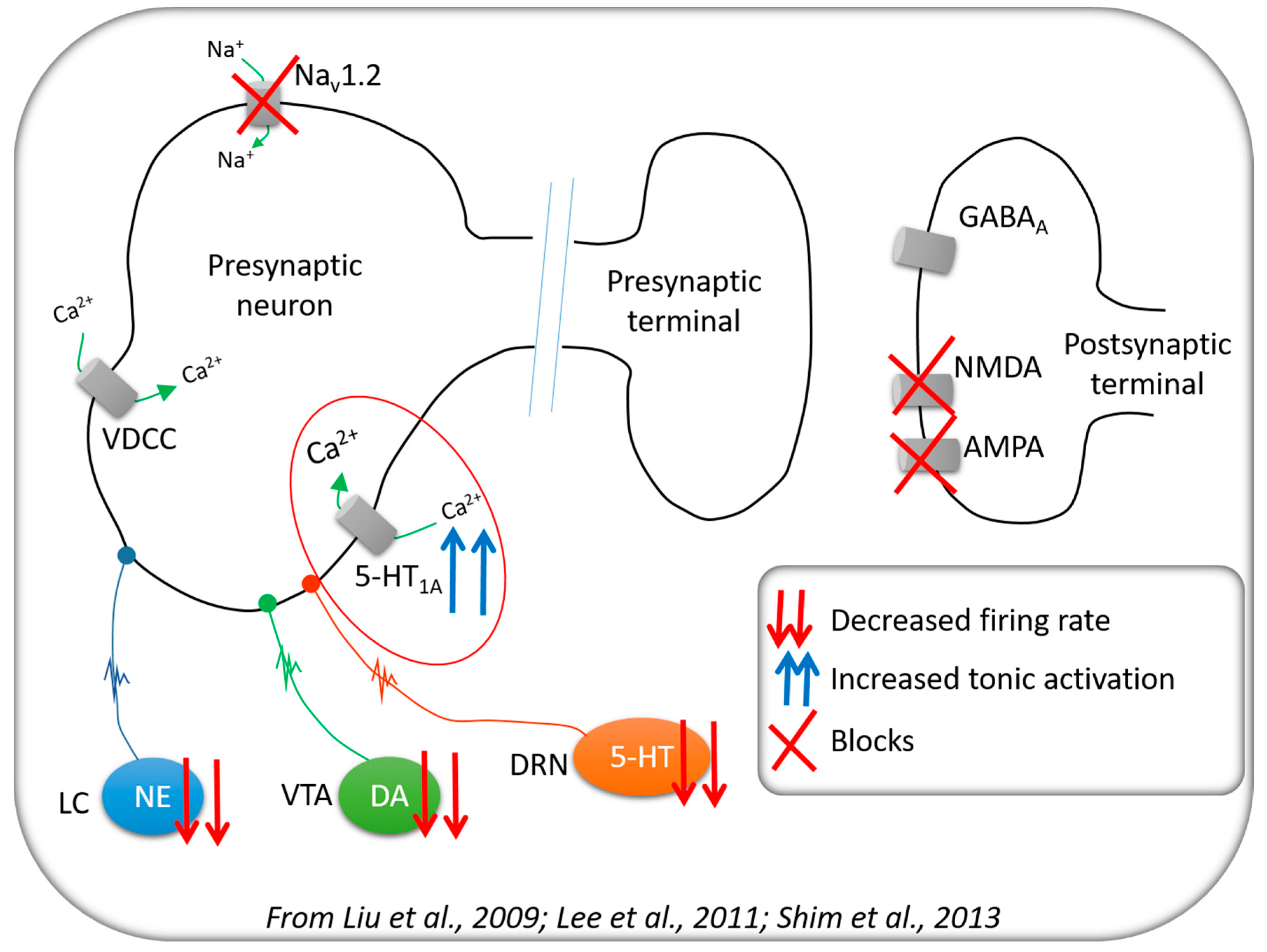
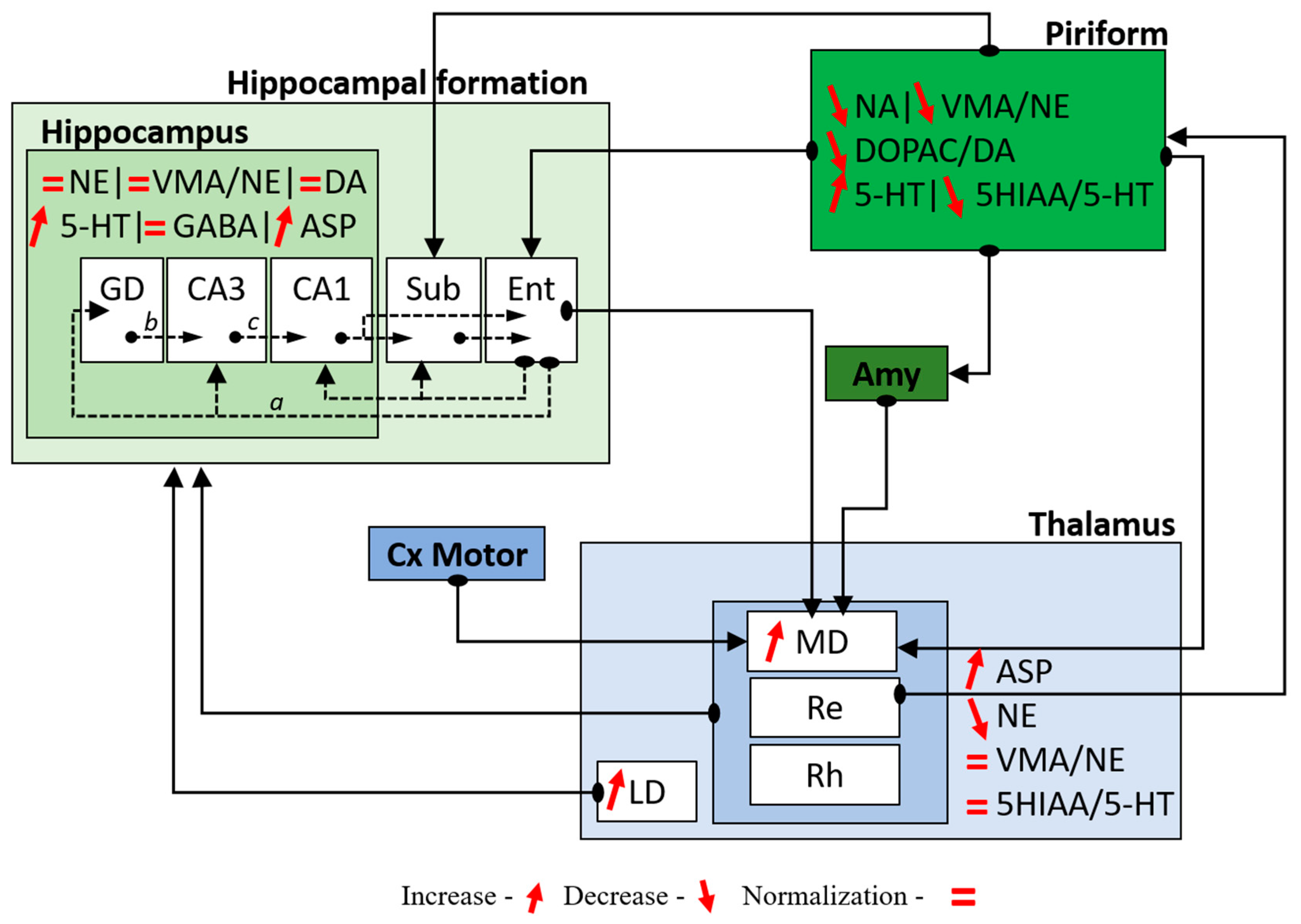
3.1. Neurochemical Changes
3.2. Cerebral Activity
4. Materials and Methods
4.1. General Procedure
4.2. Measurement of Amino Acid and Monoamine Concentrations
4.3. c-Fos Immunolabelling
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Engel, J. Excitation and inhibition in epilepsy. Can. J. Neurol. Sci. 1996, 23, 167–174. [Google Scholar] [CrossRef] [PubMed]
- French, J.A.; Williamson, P.D.; Thadani, V.M.; Darcey, T.M.; Mattson, R.H.; Spencer, S.S.; Spencer, D.D. Characteristics of medial temporal lobe epilepsy: I. Results of history and physical examination. Ann. Neurol. 1993, 34, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, N.; Bernasconi, A.; Caramanos, Z.; Antel, S.B.; Andermann, F.; Arnold, D.L. Mesial temporal damage in temporal lobe epilepsy: A volumetric MRI study of the hippocampus, amygdala and parahippocampal region. Brain J. Neurol. 2003, 126, 462–469. [Google Scholar] [CrossRef]
- Leite, J.P.; Bortolotto, Z.A.; Cavalheiro, E.A. Spontaneous recurrent seizures in rats: An experimental model of partial epilepsy. Neurosci. Biobehav. Rev. 1990, 14, 511–517. [Google Scholar] [CrossRef]
- Curia, G.; Longo, D.; Biagini, G.; Jones, R.S.G.; Avoli, M. The pilocarpine model of temporal lobe epilepsy. J. Neurosci. Methods 2008, 172, 143–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandratavicius, L.; Balista, P.A.; Lopes-Aguiar, C.; Ruggiero, R.N.; Umeoka, E.H.; Garcia-Cairasco, N.; Bueno-Junior, L.S.; Leite, J.P. Animal models of epilepsy: Use and limitations. Neuropsychiatr. Dis. Treat. 2014, 10, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, M.M. Overview of Drugs Used For Epilepsy and Seizures. Pharm. Ther. 2010, 35, 392–415. [Google Scholar]
- Fernandes, M.J.; Carneiro, J.E.M.; Amorim, R.P.; Araujo, M.G.L.; Nehlig, A. Neuroprotective agents and modulation of temporal lobe epilepsy. Front. Biosci. Elite Ed. 2015, 7, 79–93. [Google Scholar] [CrossRef] [PubMed]
- François, J.; Boehrer, A.; Nehlig, A. Effects of carisbamate (RWJ-333369) in two models of genetically determined generalized epilepsy, the GAERS and the audiogenic Wistar AS. Epilepsia 2008, 49, 393–399. [Google Scholar] [CrossRef] [PubMed]
- François, J.; Germe, K.; Ferrandon, A.; Koning, E.; Nehlig, A. Carisbamate has powerful disease-modifying effects in the lithium-pilocarpine model of temporal lobe epilepsy. Neuropharmacology 2011, 61, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Makinson, C.D.; Tanaka, B.S.; Sorokin, J.M.; Wong, J.C.; Christian, C.A.; Goldin, A.L.; Escayg, A.; Huguenard, J.R. Regulation of Thalamic and Cortical Network Synchrony by Scn8a. Neuron 2017, 93, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Danober, L.; Deransart, C.; Depaulis, A.; Vergnes, M.; Marescaux, C. Pathophysiological mechanisms of genetic absence epilepsy in the rat. Prog. Neurobiol. 1998, 55, 27–57. [Google Scholar] [CrossRef]
- Liu, Y.; Yohrling, G.J.; Wang, Y.; Hutchinson, T.L.; Brenneman, D.E.; Flores, C.M.; Zhao, B. Carisbamate, a novel neuromodulator, inhibits voltage-gated sodium channels and action potential firing of rat hippocampal neurons. Epilepsy Res. 2009, 83, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Lee, M.L.; Shih, C.C.; Liou, H.H. Carisbamate (RWJ-333369) inhibits glutamate transmission in the granule cell of the dentate gyrus. Neuropharmacology 2011, 61, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.; El Mansari, M.; Blier, P. Modulation of the antidepressant-like effects of sustained administration of carisbamate and lamotrigine on monoaminergic systems: Electrophysiological studies in the rat brain. J. Pharmacol. Exp. Ther. 2013, 347, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Marques-Carneiro, J.E.; Persike, D.S.; Litzahn, J.J.; Cassel, J.C.; Nehlig, A.; Fernandes, M.J. Hippocampal Proteome of Rats Subjected to the Li-Pilocarpine Epilepsy Model and the Effect of Carisbamate Treatment. Pharmaceuticals 2017, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Surguchov, A.; Surgucheva, I.; Sharma, M.; Sharma, R.; Singh, V. Pore-Forming Proteins as Mediators of Novel Epigenetic Mechanism of Epilepsy. Front. Neurol. 2017, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.G.; Laxer, K.D.; Barakos, J.; Cheong, I.; Finlay, D.; Garcia, P.; Cardenas-Nicolson, V.; Weiner, M.W. Involvement of the thalamocortical network in TLE with and without mesiotemporal sclerosis. Epilepsia 2010, 51, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Vergnes, M.; Marescaux, C.; Depaulis, A. Mapping of spontaneous spike and wave discharges in Wistar rats with genetic generalized non-convulsive epilepsy. Brain Res. 1990, 523, 87–91. [Google Scholar] [CrossRef]
- Cavalheiro, E.A.; Fernandes, M.J.; Turski, L.; Naffah-Mazzacoratti, M.G. Spontaneous recurrent seizures in rats: Amino acid and monoamine determination in the hippocampus. Epilepsia 1994, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Theodore, W.H. Does Serotonin Play a Role in Epilepsy? Epilepsy Curr. 2003, 3, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Bertram, E.H.; Mangan, P.S.; Zhang, D.; Scott, C.A.; Williamson, J.M. The midline thalamus: Alterations and a potential role in limbic epilepsy. Epilepsia 2001, 42, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Bertram, E.H.; Zhang, D.; Williamson, J.M. Multiple roles of midline dorsal thalamic nuclei in induction and spread of limbic seizures. Epilepsia 2008, 49, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Sloan, D.M.; Bertram, E.H. Changes in midline thalamic recruiting responses in the prefrontal cortex of the rat during the development of chronic limbic seizures. Epilepsia 2009, 50, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Sloan, D.M.; Zhang, D.; Bertram, E.H. Increased GABAergic inhibition in the midline thalamus affects signaling and seizure spread in the hippocampus-prefrontal cortex pathway. Epilepsia 2011, 52, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Sloan, D.M.; Zhang, D.; Bertram, E.H. Excitatory amplification through divergent-convergent circuits: The role of the midline thalamus in limbic seizures. Neurobiol. Dis. 2011, 43, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Bertram, E.H.; Zhang, D.X.; Mangan, P.; Fountain, N.; Rempe, D. Functional anatomy of limbic epilepsy: A proposal for central synchronization of a diffusely hyperexcitable network. Epilepsy Res. 1998, 32, 194–205. [Google Scholar] [CrossRef]
- Cassidy, R.M.; Gale, K. Mediodorsal thalamus plays a critical role in the development of limbic motor seizures. J. Neurosci. 1998, 18, 9002–9009. [Google Scholar] [PubMed]
- Patel, S.; Millan, M.H.; Meldrum, B.S. Decrease in excitatory transmission within the lateral habenula and the mediodorsal thalamus protects against limbic seizures in rats. Exp. Neurol. 1988, 101, 63–74. [Google Scholar] [CrossRef]
- Banerjee, P.K.; Snead, O.C. Thalamic mediodorsal and intralaminar nuclear lesions disrupt the generation of experimentally induced generalized absence-like seizures in rats. Epilepsy Res. 1994, 17, 193–205. [Google Scholar] [CrossRef]
- Kato, K.; Urino, T.; Hori, T.; Tsuda, H.; Yoshida, K.; Hashizume, K.; Tanaka, T. Experimental petit mal-like seizure induced by microinjection of kainic acid into the unilateral mediodorsal nucleus of the thalamus. Neurol. Med. Chir. (Tokyo) 2008, 48, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Scholl, E.A.; Dudek, F.E.; Ekstrand, J.J. Neuronal degeneration is observed in multiple regions outside the hippocampus after lithium pilocarpine-induced status epilepticus in the immature rat. Neuroscience 2013, 252, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.L.; Morrow, D.; Liu, S.; Liu, K.J. Hydroethidine detection of superoxide production during the lithium–pilocarpine model of status epilepticus. Epilepsy Res. 2002, 49, 226–238. [Google Scholar] [CrossRef]
- Jafarian, M.; Karimzadeh, F.; Alipour, F.; Attari, F.; Lotfinia, A.A.; Speckmann, E.J.; Zarrindast, M.R.; Gorji, A. Cognitive impairments and neuronal injury in different brain regions of a genetic rat model of absence epilepsy. Neuroscience 2015, 298, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Pinault, D. The thalamic reticular nucleus: Structure, function and concept. Brain Res. Brain Res. Rev. 2004, 46, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Morrisett, R.A.; Jope, R.S.; Snead, O.C. Effects of drugs on the initiation and maintenance of status epilepticus induced by administration of pilocarpine to lithium-pretreated rats. Exp. Neurol. 1987, 97, 193–200. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Australian Academic Press: Samford Valley, Australia, 2007; ISBN 978-0-12-547612-6. [Google Scholar]
- Gundersen, H.J.; Jensen, E.B.; Kiêu, K.; Nielsen, J. The efficiency of systematic sampling in stereology—Reconsidered. J. Microsc. 1999, 193, 199–211. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hippocampus | Thalamus | Piriform Cortex | ||||||||||||||
| CT | DZP | CRS | ANOVA | BS | CT | DZP | CRS | ANOVA | BS | CT | DZP | CRS | ANOVA | BS | ||
| Monoamines (ng/g) | NE | 0.39 | 0.12 * | 0.28 # | F(2,15) = 20.73; p < 0.001 | p < 0.001 | 0.71 | 0.33 * | 0.51 * | F(2,15) = 19.76; p < 0.001 | p < 0.001 | 0.58 | 0.23 * | 0.32 * | F(2,15) = 35.88; p < 0.001 | p < 0.001 |
| VMA | 1.64 | 1.43 | 1.41 | F(2,15) = 1.64; p = 0.226 | p = 0.269 | 1.32 | 1.27 | 1.42 | F(2,15) = 0.99; p = 0.392 | p = 0.307 | 7.02 | 6.08 | 6.03 | F(2,15) = 2.67; p = 0.101 | p = 0.160 | |
| VMA/NE | 4.27 | 14.58 * | 5.16 # | F(2,15) = 9.33; p = 0.002 | p = 0.002 | 1.90 | 4.14 * | 2.84 # | F(2,15) = 18.07; p < 0.001 | p < 0.001 | 12.40 | 26.82 * | 19.67 *# | F(2,15) = 21.03; p < 0.001 | p < 0.001 | |
| DA | 0.07 | 0.12 | 0.07 | F(2,15) = 4.31; p = 0.033 | p = 0.087 | 0.05 | 0.09 | 0.06 | F(2,15) =2.35; p = 0.129 | p = 0.172 | 0.15 | 0.13 | 0.23 | F(2,15) = 3.73; p = 0.048 | p = 0.068 | |
| DOPAC | 0.01 | 0.02 | 0.02 | F(2,15) = 0.67; p = 0.521 | p = 0.383 | 0.01 | 0.03 | 0.01 | F(2,15) = 3.19; p = 0.069 | p = 0.128 | 0.07 | 0.07 | 0.03 *# | F(2,15) = 15.12; p < 0.001 | p = 0.011 | |
| DOPAC/DA | 0.23 | 0.17 | 0.24 | F(2,15) = 0.74; p = 0.490 | p = 0.311 | 0.06 | 0.29 | 0.15 | F(2,15) = 2.59; p = 0.107 | p = 0.137 | 0.50 | 0.57 * | 0.12 * | F(2,15) = 23.35; p <0 .001 | p < 0.001 | |
| HVA | 0.07 | 0.08 | 0.08 | F(2,15) = 0.06; p = 0.941 | p = 0.492 | 0.05 | 0.09 | 0.06 | F(2,15) = 1.44; p = 0.267 | p = 0.263 | 0.05 | 0.04 | 0.16 | F(2,15) = 0.71; p = 0.504 | p = 0.340 | |
| HVA/DA | 1.04 | 0.66 | 1.06 | F(2,15) = 3.78; p = 0.046 | p = 0.051 | 1.53 | 0.88 | 1.23 | F(2,15) = 1.45; p = 0.264 | p = 0.227 | 0.73 | 0.55 | 1.51 # | F(2,15) = 7.08; p < 0.007 | p < 0.01 | |
| 5-HT | 0.68 | 0.65 | 0.95 *# | F(2,15) = 8.50; p = 0.003 | p = 0.007 | 1.13 | 1.12 | 1.29 | F(2,15) = 1.94; p = 0.177 | p = 0.172 | 0.65 | 0.43 | 1.01 # | F(2,15) = 8.58; p = 0.003 | p = 0.011 | |
| 5HIAA | 0.30 | 0.51 * | 0.48 * | F(2,15) = 11.55; p < 0.001 | p < 0.001 | 0.64 | 1.04 * | 0.84 | F(2,15) = 13.16; p < 0.001 | p < 0.001 | 0.23 | 0.26 | 0.18 | F(2,15) = 4.28; p = 0.033 | p = 0.077 | |
| 5HIAA/5-HT | 4.76 | 7.36 | 6.54 | F(2,15) = 3.09; p = 0.075 | p = 0.136 | 0.57 | 0.92 * | 0.67 # | F(2,15) = 13.83; p < 0.001 | p < 0.001 | 0.38 | 0.49 | 0.18 *# | F(2,15) = 18.61; p < 0.001 | p < 0.001 | |
| Hippocampus | Thalamus | |||||||||||||||
| CT | DZP | CRS | ANOVA | BS | CT | DZP | CRS | ANOVA | BS | |||||||
| Amino acids (nmol/mg) | ASP | 1.77 | 1.51 | 2.23 # | F(2,15) = 8.59; p = 0.003 | p = 0.005 | 2.18 | 1.80 | 2.26 # | F(2,15) = 6.93; p = 0.007 | p = 0.013 | |||||
| GLU | 8.42 | 8.37 | 9.11 | F(2,15) = 0.51; p = 0.608 | p = 0.382 | 8.89 | 8.57 | 8.22 | F(2,15) = 0.35; p = 0.705 | p = 0.398 | ||||||
| GLN | 3.63 | 4.65 * | 4.67 * | F(2,15) = 9.94; p < 0.001 | p = 0.002 | 4.04 | 5.37 * | 5.06 * | F(2,15) = 17.57; p < 0.001 | p < 0.001 | ||||||
| GLY | 0.38 | 0.44 | 0.44 | F(2,15) = 1.00; p = 0.388 | p = 0.288 | 0.48 | 0.50 | 0.49 | F(2,15) = 0.07; p = 0.932 | p = 0.932 | ||||||
| TAU | 5.39 | 4.98 | 5.55 | F(2,15) = 0.78; p = 0.471 | p = 0.311 | 2.36 | 2.15 | 2.38 | F(2,15) = 1.11; p = 0.354 | p = 0.292 | ||||||
| GABA | 2.00 | 2.70 * | 2.24 | F(2,15) = 4.99; p = 0.021 | p = 0.045 | 2.55 | 2.51 | 2.23 | F(2,15) = 1.90; p = 0.182 | p = 0.156 | ||||||
| Structure | Number of Cells | ANOVA | Bootstrap | ||
|---|---|---|---|---|---|
| CT | DZP | CRS | |||
| PV | 6471 | 37,435 * | 43,927 * | F(2,12) = 77.17; p < 0.001 | p < 0.001 |
| IL | 1275 | 26,498 * | 28,314 * | F(2,12) = 119.88; p < 0.001 | p < 0.001 |
| ReRh | 995 | 21,466 * | 27,867 * | F(2,12) = 73.00; p < 0.001 | p < 0.001 |
| MD | 1571 | 4517 | 10,588 *# | F(2,12) = 21.85; p < 0.001 | p < 0.001 |
| RTN | 880 | 29,169 * | 25,824 * | F(2,12) = 52.09; p < 0.001 | p < 0.001 |
| ZI | 1537 | 11,858 * | 13,409 * | F(2,12) = 6.08; p = 0.015 | p = 0.009 |
| VB | 852 | 4866 * | 7562 * | F(2,12) = 10.59; p = 0.002 | p = 0.003 |
| LD | 1291 | 6698 * | 13,880 *# | F(2,12) = 33.66; p < 0.001 | p < 0.001 |
| GD | 2865 | 234,694 * | 208,044 * | F(2,12) = 68.35; p < 0.001 | p < 0.001 |
| CA3 | 5832 | 79,664 * | 80,145 * | F(2,12) = 48.48; p < 0.001 | p < 0.001 |
| CA1 | 2724 | 130,109 * | 123,864 * | F(2,12) = 31.19; p < 0.001 | p < 0.001 |
| Pir | 3778 | 213,143 * | 203,401 * | F(2,12) = 70.09; p < 0.001 | p < 0.001 |
| Ent | 5117 | 153,586 * | 136,315 * | F(2,12) = 47.08; p < 0.001 | p < 0.001 |
| Amy | 4097 | 74,388 * | 93,834 * | F(2,12) = 18.78; p < 0.001 | p < 0.001 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques-Carneiro, J.E.; Nehlig, A.; Cassel, J.-C.; Castro-Neto, E.F.; Litzahn, J.J.; Pereira de Vasconcelos, A.; Naffah-Mazacoratti, M.D.G.; Fernandes, M.J.d.S. Neurochemical Changes and c-Fos Mapping in the Brain after Carisbamate Treatment of Rats Subjected to Lithium–Pilocarpine-Induced Status Epilepticus. Pharmaceuticals 2017, 10, 85. https://doi.org/10.3390/ph10040085
Marques-Carneiro JE, Nehlig A, Cassel J-C, Castro-Neto EF, Litzahn JJ, Pereira de Vasconcelos A, Naffah-Mazacoratti MDG, Fernandes MJdS. Neurochemical Changes and c-Fos Mapping in the Brain after Carisbamate Treatment of Rats Subjected to Lithium–Pilocarpine-Induced Status Epilepticus. Pharmaceuticals. 2017; 10(4):85. https://doi.org/10.3390/ph10040085
Chicago/Turabian StyleMarques-Carneiro, José Eduardo, Astrid Nehlig, Jean-Christophe Cassel, Eduardo Ferreira Castro-Neto, Julia Julie Litzahn, Anne Pereira de Vasconcelos, Maria Da Graça Naffah-Mazacoratti, and Maria José da Silva Fernandes. 2017. "Neurochemical Changes and c-Fos Mapping in the Brain after Carisbamate Treatment of Rats Subjected to Lithium–Pilocarpine-Induced Status Epilepticus" Pharmaceuticals 10, no. 4: 85. https://doi.org/10.3390/ph10040085
APA StyleMarques-Carneiro, J. E., Nehlig, A., Cassel, J.-C., Castro-Neto, E. F., Litzahn, J. J., Pereira de Vasconcelos, A., Naffah-Mazacoratti, M. D. G., & Fernandes, M. J. d. S. (2017). Neurochemical Changes and c-Fos Mapping in the Brain after Carisbamate Treatment of Rats Subjected to Lithium–Pilocarpine-Induced Status Epilepticus. Pharmaceuticals, 10(4), 85. https://doi.org/10.3390/ph10040085