Nucleic Acid Aptamers Targeting Epigenetic Regulators: An Innovative Therapeutic Option

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
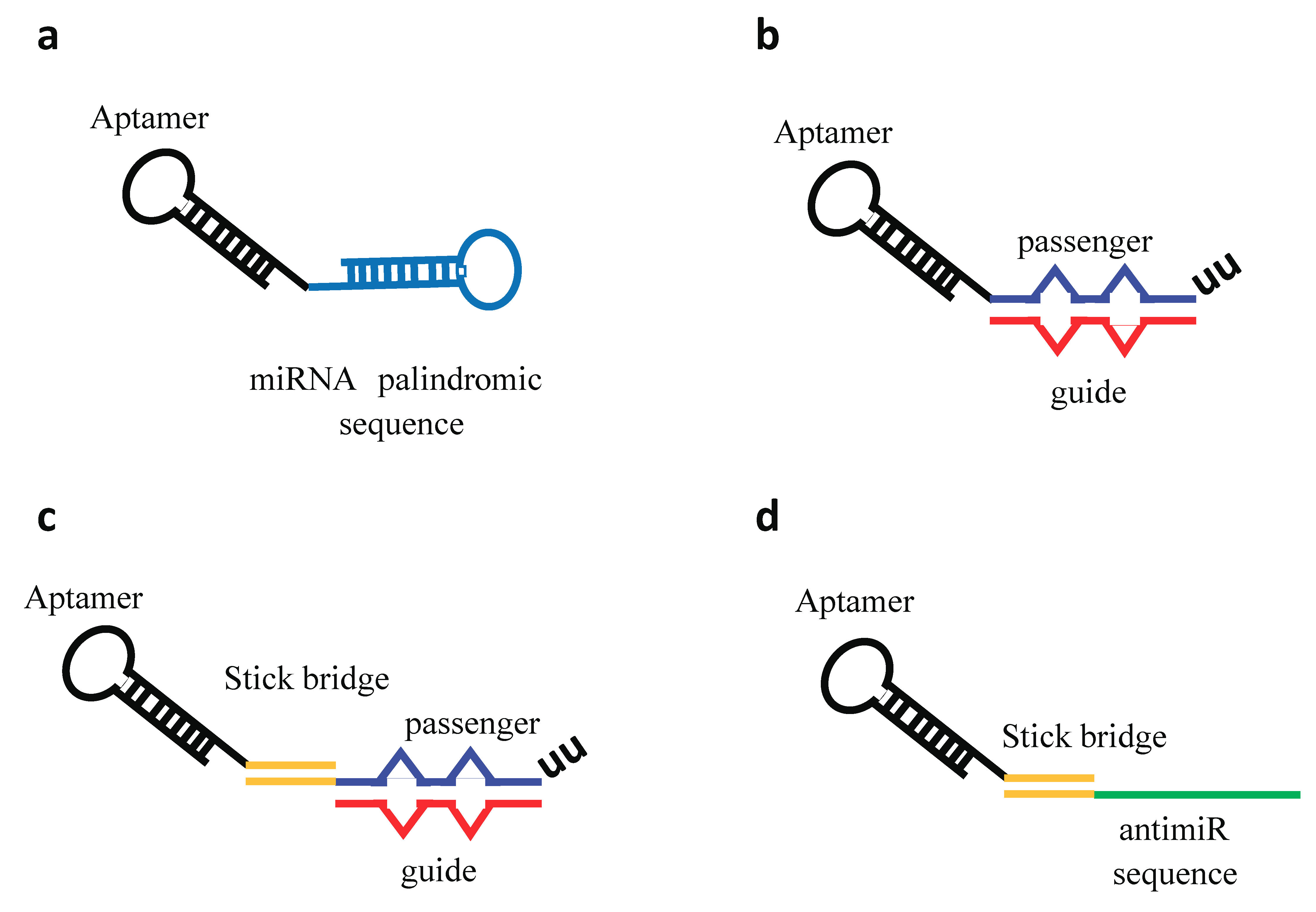
:1. Introduction
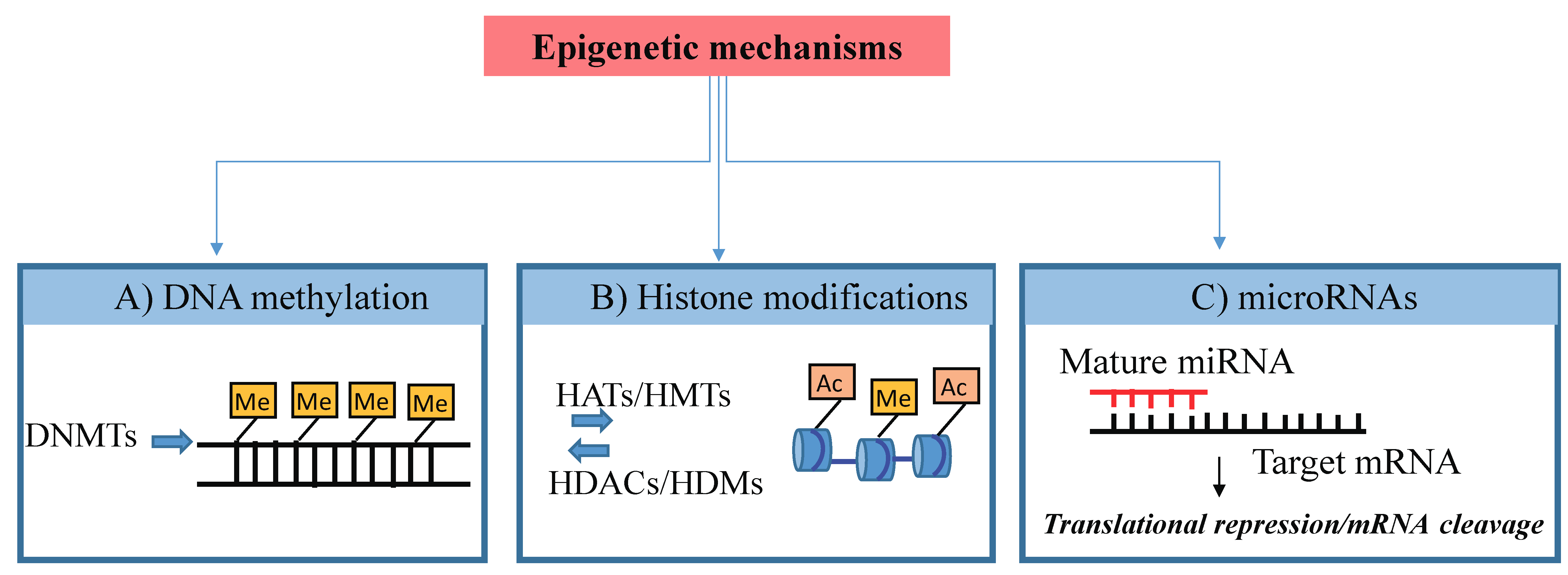
2. Overview of the Epigenetics’ Processes
2.1. DNA Methylation
2.2. Post-Translational Modifications of Histones
2.3. Noncoding RNAs
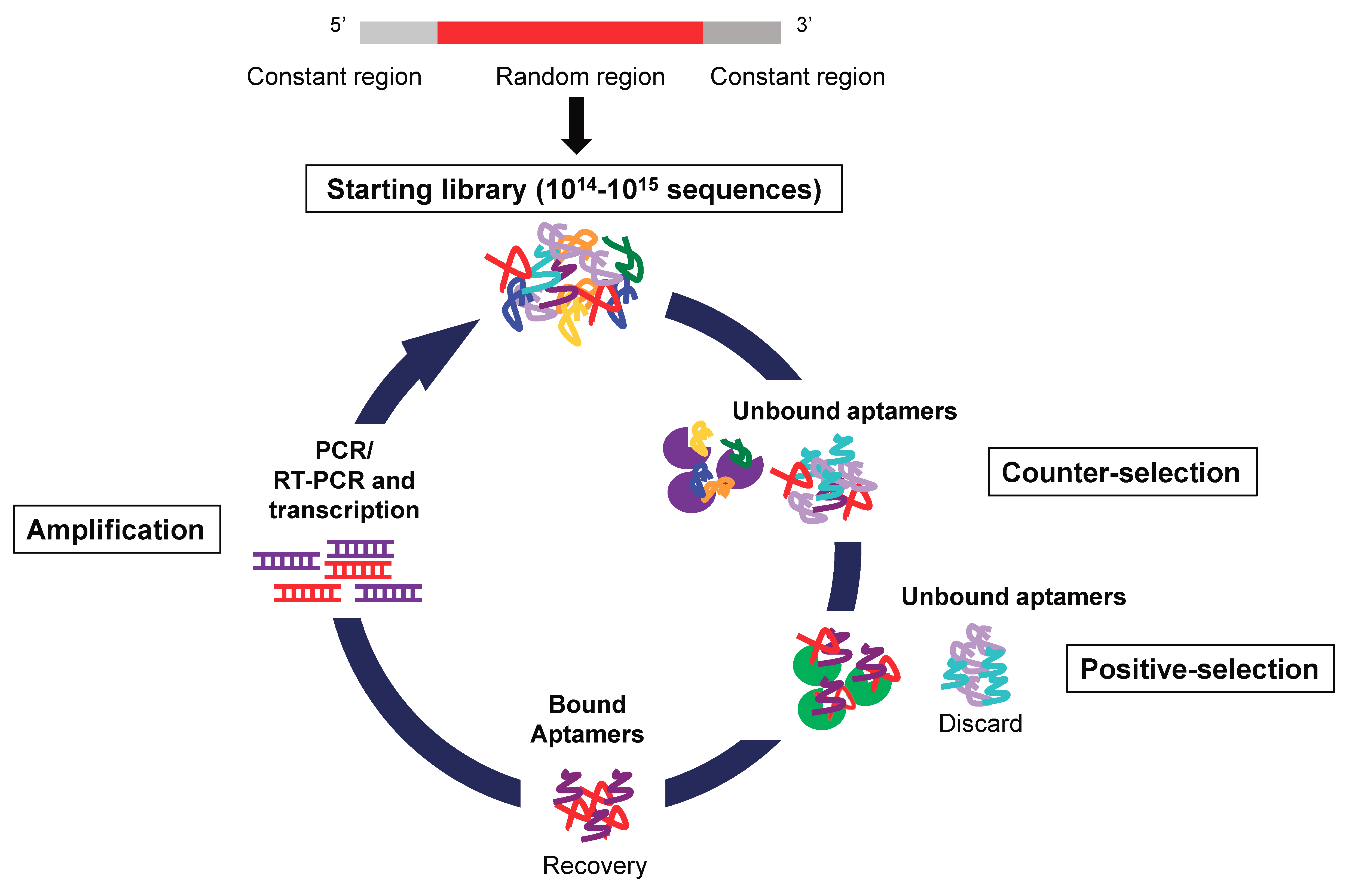
3. Aptamers
3.1. Aptamer Production
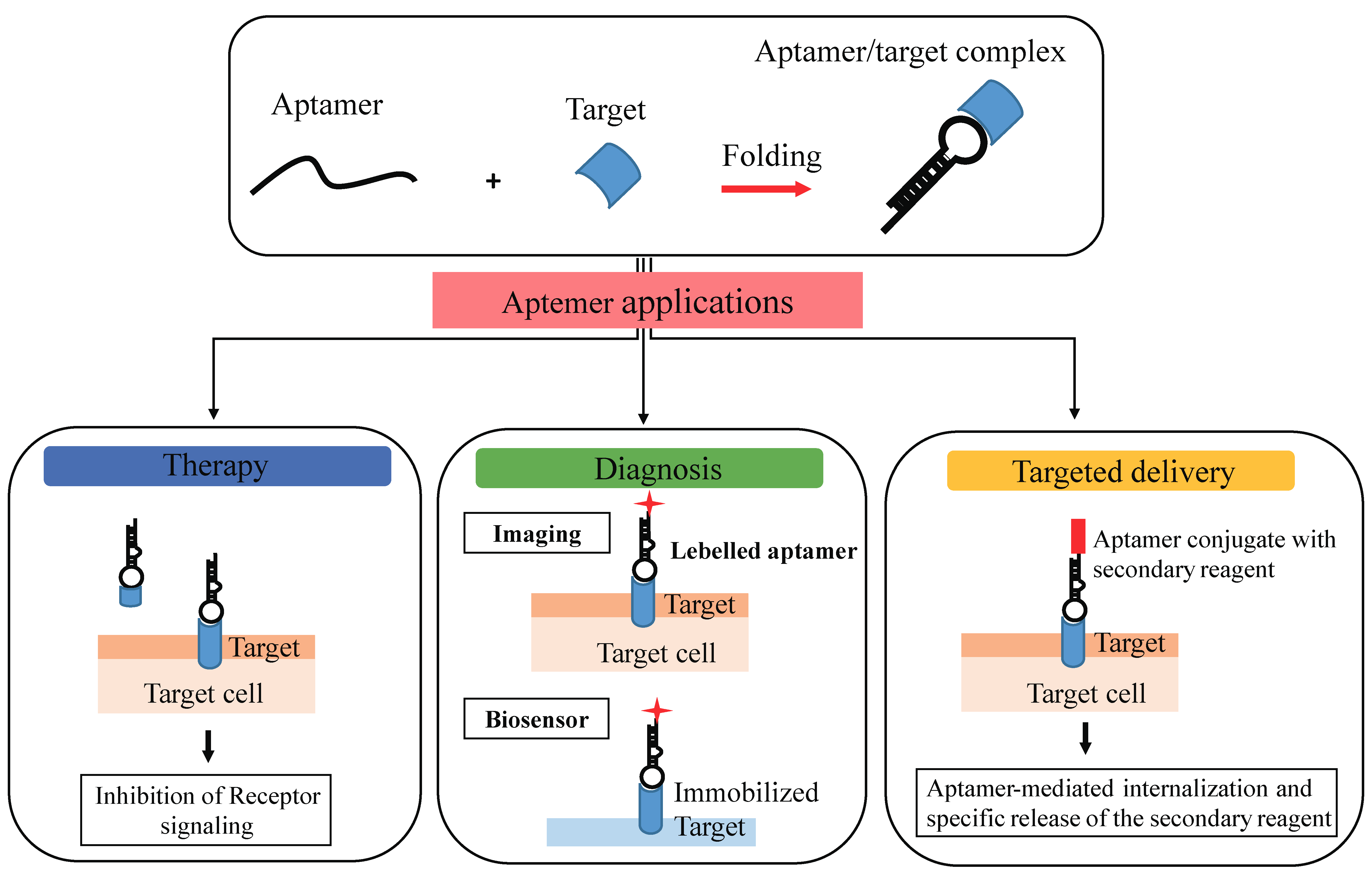
3.2. Aptamer Advantages as Diagnostic and Therapeutic Tools
4. Aptamer-Based Targeting of Epigenetics Marks
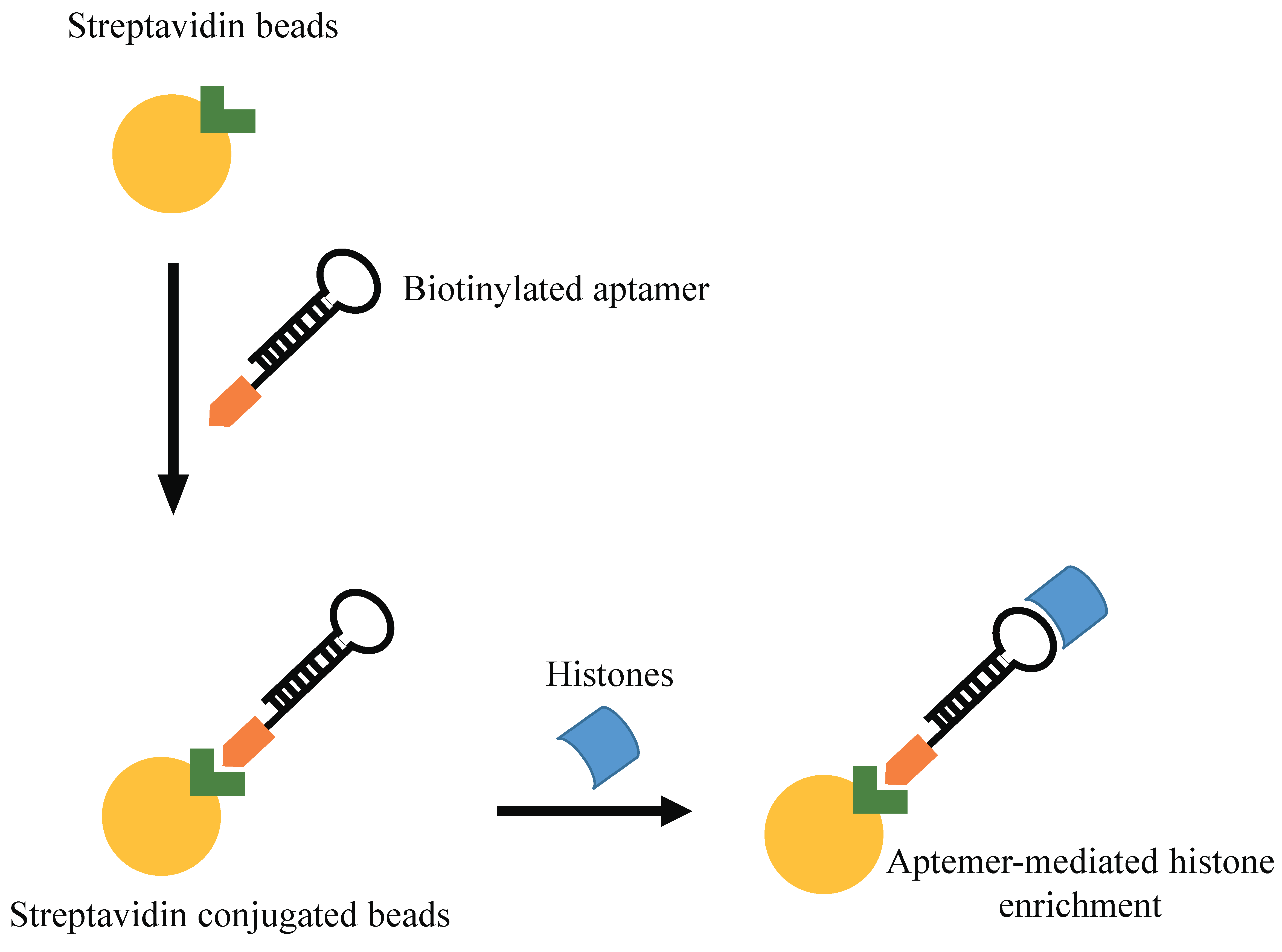
4.1. Aptamer-Mediated Targeting of Epigenetic Players
4.2. Aptamers as Delivery Carriers
4.3. Aptamer-Based Biosensors
5. SELEX-Based Strategies to Investigate Mechanisms of Gene Expression
6. Conclusions
7. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADAR | adenosine deaminase acting on RNA |
| CE | capillary electrophoresis |
| DNMTs | DNA methyltransferases |
| DNMTi | DNA N-methyl transferase inhibitors |
| GBM | glioblastoma |
| HATs | histone acetyltransferases |
| HDACs | histone deacetylases |
| HDACi | histone deacetylase inhibitors |
| HMTs | histone lysine methyltransferases |
| HDMs | lysine demethylases |
| HT-SELEX | high-throughput SELEX |
| Kd | dissociation constant |
| miRNAs | microRNAs |
| MUC1 | Mucin 1 |
| NSs | nanosheets |
| ncRNAs | noncoding RNAs |
| NSCLC | non-small-cell lung carcinoma |
| PDGFR | Platelet-derived growth factor receptor |
| PTEN | phosphatase and tensin homolog deleted on chromosome ten |
| PTMs | post-translational modifications |
| QDs | quantum dots |
| RISC | RNA-induced silencing complex |
| SAM | S-adenosylmethionine |
| SELEX | Systematic Evolution of Ligands by Exponential enrichment |
| TFs | transcription factors |
| UTR | untranslated region |
References
- Waterland, R.A.; Michels, K.B. Epigenetic epidemiology of the developmental origins hypothesis. Annu. Rev. Nutr. 2007, 27, 363–388. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Samaranayake, M.; Pradhan, S. Epigenetic mechanisms in mammals. Cell. Mol. Life Sci. 2009, 66, 596–612. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. Epigenetic mechanisms for nutrition deterrminants of later health outcomes. Am. J. Clin. Nutr. 2009, 89, 1488S–1493S. [Google Scholar] [CrossRef] [PubMed]
- Sabatti, C.; Service, S.K.; Hartikainen, A.L.; Pouta, A.; Ripatti, S.; Brodsky, J.; Jones, C.G.; Zaitlen, N.A.; Varilo, T.; Kaakinen, M.; et al. Genome-wide association analysis of metabolic traits in a birth cohort from a founder population. Nat. Genet. 2009, 41, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, W. Epigenetic impact of dietary polyphenols in cancer chemoprevention: Lifelong remodeling of our epigenomes. Pharmacol. Res. 2012, 65, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Petronis, A. Epigenetics as a unifying principle in the aetiology of complex traits and diseases. Nature 2010, 465, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.X.; Riggs, A.D. DNA methylation and demethylation in mammals. J. Biol. Chem. 2011, 286, 18347–18353. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; D’Alessio, A.C.; Taranova, O.V.; Hong, K.; Sowers, L.C.; Zhang, Y. Role of tet proteins in 5mc to 5hmc conversion, es-cell self-renewal and inner cell mass specification. Nature 2010, 466, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.; Thornton, J. Understanding the molecular machinery of genetics through 3d structures. Nat. Rev. Genet. 2008, 9, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Bulger, M.; Muramatsu, M.; Ito, T. Multistep chromatin assembly of supercoiled plasmid DNA by nucleosome assembly protein-1 and atp-utilizing chromatin assembly and remodeling factor. J. Biol. Chem. 2001, 276, 27384–27391. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- The ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, V.W.; Goren, A.; Bernstein, B.E. Charting histone modifications and the functional organization of mammalian genomes. Nat. Rev. Genet. 2011, 12, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Li, Y.; Ren, B.; Natarajan, R. Enhancers: Multidimensional signal integrators. Transcription 2011, 2, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Vastolo, V.; Nettore, I.C.; Ciccarelli, M.; Albano, L.; Raciti, G.A.; Longo, M.; Beguinot, F.; Ungaro, P. High-fat diet unveils an enhancer element at the ped/pea-15 gene responsible for epigenetic memory in skeletal muscle. Metab. Clin. Exp. 2018. [Google Scholar] [CrossRef] [PubMed]
- Sassone-Corsi, P. When metabolism and epigenetics converge. Science 2013, 339, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Rouhi, A.; Mager, D.L.; Humphries, R.K.; Kuchenbauer, F. MiRNAs, epigenetics, and cancer. Mamm. Genome 2008, 19, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Morlando, M.; Fatica, A. Alteration of epigenetic regulation by long noncoding RNAs in cancer. Int. J. Mol. Sci. 2018, 19, 570. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farth, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. Causes and consequences of micro-RNA dysregulation. Cancer J. 2012, 18, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Serra, P.; Esteller, M. DNA methylation-associated silencing of tumor-suppressor microRNAs in cancer. Oncogene 2012, 31, 1609–1622. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. DNA methylation and microRNA dysregulation in cancer. Mol. Oncol. 2012, 6, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, Y.; Suzuki, H.; Taya, T.; Nishizawa, M.; Tsugawa, H.; Matsuzaki, J.; Hirata, K.; Saito, H.; Hibi, T. Development of a novel microRNA promoter microarray for chip-on-chip assay to identify epigenetically regulated microRNAs. Biochem. Biophys. Res. Commun. 2012, 426, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liang, H. When microRNAs meet RNA editing in cancer: A nucleotide change can make a difference. Bioessays 2018, 40. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage t4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.S.; Matthews, C.S.; Missailidis, S. DNA aptamers that bind to muc1 tumour marker: Design and characterization of muc1-binding single-stranded DNA aptamers. Tumour Biol. 2006, 27, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Lupold, S.E.; Hicke, B.J.; Lin, Y.; Coffey, D.S. Identification and characterization of nuclease-stabilized RNA molecules that bind human prostate cancer cells via the prostate-specific membrane antigen. Cancer Res. 2012, 62, 4029–4033. [Google Scholar]
- Chen, C.H.; Chernis, G.A.; Hoang, V.Q.; Landgraf, R. Inhibition of heregulin signaling by an aptamer that preferentially binds to the oligomeric form of human epidermal growth factor receptor-3. Proc. Natl. Acad. Sci. USA 2003, 100, 9226–9231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Larson, T.; Nguyen, H.H.; Sokolov, K.V.; Ellington, A.D. Directed evolution of gold nanoparticle delivery to cells. Chem. Commun. 2010, 46, 392–394. [Google Scholar] [CrossRef] [PubMed]
- Catuogno, S.; Esposito, C.L. Aptamer cell-based selection: Overview and advances. Biomedicines 2017, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Catuogno, S.; Esposito, C.L.; de Franciscis, V. Developing aptamers by cell-based selex. Methods Mol. Biol. 2016, 1380, 33–46. [Google Scholar] [PubMed]
- Chen, M.; Yu, Y.; Jiang, F.; Zhou, J.; Li, Y.; Liang, C.; Dang, L.; Lu, A.; Zhang, G. Development of cell-selex technology and its application in cancer diagnosis and therapy. Int. J. Mol. Sci. 2016, 17, 79. [Google Scholar] [CrossRef] [PubMed]
- Darmostuk, M.; Rimpelova, S.; Gbelcova, H.; Ruml, T. Current approaches in selex: An update to aptamer selection technology. Biotechnol. Adv. 2015, 33, 1141–1161. [Google Scholar] [CrossRef] [PubMed]
- Keefe, A.D.; Cload, S.T. Selex with modified nucleotides. Curr. Opin. Chem. Biol. 2008, 12, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.S.; Borkowski, S.; Kurreck, J.; Stephens, A.W.; Bald, R.; Hecht, M.; Friebe, M.; Dinkelborg, L.; Erdmann, V.A. Application of locked nucleic acids to improve aptamer in vivo stability and targeting function. Nucleic Acids Res. 2004, 32, 5757–5765. [Google Scholar] [CrossRef] [PubMed]
- Healy, J.M.; Lewis, S.D.; Kurz, M.; Boomer, R.M.; Thompson, K.M.; Wilson, C.; McCauley, T.G. Pharmacokinetics and biodistribution of novel aptamer compositions. Pharm. Res. 2004, 21, 2234–2246. [Google Scholar] [CrossRef] [PubMed]
- Catuogno, S.; Esposito, C.L.; Condorelli, G.; De Franciscis, V. Nucleic acids delivering nucleic acids. Adv. Drug Deliv. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ausiò, J.; Moore, S.C. Reconstitution of chromatin complexes from high-performance liquid chromatography-purified histones. Methods 1998, 15, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Gurley, L.R.; Prentice, D.A.; Valdez, J.G.; Spall, W.D. High-performance liquid chromatography of chromatin histones. J. Chromatogr. 1983, 266, 609–627. [Google Scholar] [CrossRef]
- Lindner, H.H. Analysis of histones, histone variants, and their post-translationally modified forms. Electrophoresis 2008, 29, 2516–2532. [Google Scholar] [CrossRef] [PubMed]
- Chambers, R.S. High-throughput antibody production. Curr. Opin. Chem. Biol. 2005, 1, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Bunka, D.H.; Stockley, P.G. Aptamers come of age—At last. Nat. Rev. Microbiol. 2006, 8, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Hom, D.; Lindsay, S.M.; Chaput, J.C. In vitro selection of histone h4 aptamers for recognition imaging microscopy. J. Am. Chem. Soc. 2007, 129, 14568–14569. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.A.; Lin, L.; Lindsay, S.M.; Chaput, J.C. Evolution of a histone h4-k16 acetyl-specific DNA aptamer. J. Am. Chem. Soc. 2009, 131, 6330–6331. [Google Scholar] [CrossRef] [PubMed]
- Shao, N.; Zhang, K.; Chen, Y.; He, X.; Zhang, Y. Preparation and characterization of DNA aptamer based spin column for enrichment and separation of histones. Chem. Commun. 2012, 48, 6684–6686. [Google Scholar] [CrossRef] [PubMed]
- Hyun, S.; Lee, K.H.; Han, A.; Yu, J. An RNA aptamer that selectively recognizes symmetric dimethylation of arginine 8 in the histone h3 n-terminal peptide. Nucleic Acid Ther. 2011, 21, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Garzon, R.; Cimmino, A.; Liu, Z.; Zanesi, N.; Callegari, E.; Liu, S.; Alder, H.; Costinean, S.; Fernandez-Cymering, C.; et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3a and 3b. Proc. Natl. Acad. Sci. USA 2007, 104, 15805–15810. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.H.; Lee, H.S.; Kim, W.H. Promoter methylation and silencing of pten in gastric carcinoma. Lab. Investig. 2002, 82, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Marsit, C.J.; Zheng, S.; Aldape, K.; Hinds, P.W.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. Pten expression in non-small-cell lung cancer: Evaluating its relation to tumor characteristics, allelic loss, and epigenetic alterations. Hum. Pathol. 2005, 36, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Tsunoda, H.; Nishida, M.; Morishita, Y.; Takimoto, Y.; Kubo, T.; Noguchi, M. Loss of heterozygosity on 10q23.3 and mutation of the tumor suppressor gene pten in benign endometrial cyst of the ovary: Possible sequence progression from benign endomtrial cyst to endometrioid carcinoma and clear cell carcinoma of the ovary. Cancer Res. 2000, 60, 7052–7056. [Google Scholar] [PubMed]
- Ho, C.M.; Lin, M.C.; Huang, S.H.; Huang, C.J.; lai, H.C.; Chien, T.Y.; Chang, S.F. Pten promoter methylation and loh of 10q22-23 locus in pten expression of ovarian clear cell adenocarcinomas. Gynecol. Oncol. 2009, 112, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Zhang, Y.; Zhu, X.; Shan, N.; Chen, Y. Anticancer role of muc1 aptamer-mir-29b chimera in epithelial ovarian carcinoma cells through regulation of pten methylation. Targt. Oncol. 2012, 7, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Constantinou, P.F.; Danysh, B.P.; Dharmaraj, N.; Carson, D.D. Transmembrane mucins as novel therapeutic targets. Expert Rev. Endocrinol. Metab. 2011, 6, 835–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Zhou, C.; Zhao, J.; Chen, Y. Reversal of paclitaxel resistance in epithelial ovarian carcinoma cells by a muc1 aptamer-let-7i chimera. Cancer Investig 2012, 30, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Iaboni, M.; Russo, V.; Fontanella, R.; Roscigno, G.; Fiore, D.; Donnarumma, E.; Esposito, C.L.; Quintavalle, C.; Giangrande, P.H.; de Franciscis, V.; et al. Aptamer-miRNA-212 conjugate sensitizes nsclc cells to trail. Mol. Ther. Nucleic Acids 2016, 5, e289. [Google Scholar] [CrossRef] [PubMed]
- Esposito, C.L.; Cerchia, L.; Catuogno, S.; De Vita, G.; Dassie, J.P.; Santamaria, G.; Swiderski, P.; Condorelli, G.; Giangrande, P.H.; de Franciscis, V. Multifunctional aptamer-miRNA conjugates for targeted cancer therapy. Mol. Ther. 2104, 22, 1151–1163. [Google Scholar] [CrossRef] [PubMed]
- Catuogno, S.; Rienzo, A.; Di Vito, A.; Esposito, C.L.; de Franciscis, V. Selective delivery of therapeutic single strand antimirs by aptamer-based conjugates. J. Control. Release 2015, 210, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Esposito, C.L.; Nuzzo, S.; Kumar, S.A.; Rienzo, A.; Lawrence, C.L.; Pallini, R.; Shaw, L.; Alder, J.E.; Ricci-Vitiani, L.; Catuogno, S.; et al. A combined microRNA-based targeted therapeutic approach to eradicate glioblastoma stem-like cells. J. Control. Release 2016, 238, 43–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, B.N.; Mutharasan, R. Biosensor-based microRNA detection: Techniques, design, performance, and challenges. Analyst 2014, 139, 1576–1588. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Qi, J.; Zhang, Y.; Ren, Y.; Su, M.; Jia, B.; Wang, Y.; Wei, Q.; Du, B. Ultrasensitive photoelectrochemical aptasensing of mir-155 using efficient and stable ch3nh3pbi3 quantum dots sensitized zno nanosheets as light harvester. Biosens. Bioelectron. 2016, 85, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Zhao, L.; Bao, T.; Wen, W.; Zhang, X.; Wang, S. Integrated amplified aptasensor with in-situ precise preparation of copper nanoclusters for ultrasensitive electrochemical detection of microRNA 21. Biosens. Bioelectron. 2017, 98, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Morgunova, E.; Jolma, A.; Kaasinen, E.; Sahu, B.; Khund-Sayeed, S.; Das, P.K.; Kivioja, T.; Dave, K.; Zhong, F.; et al. Impact of cytosine methylation on DNA binding specificities of human transcription factors. Science 2017, 356. [Google Scholar] [CrossRef] [PubMed]
- Zaghlool, S.B.; Mook-Kanamori, D.O.; Kader, S.S.; Stephan, N.; Halama, A.; Engelke, R.; Sarwath, H.; Al-Dous, E.K.; Mohamoud, Y.A.; Roemisch-Margl, W.; et al. Deep molecular phenotypes link complex disorders and physiological insult to cpg methylation. Hum. Mol. Genet. 2018, 27, 1106–1121. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Jenison, R.D.; Gill, S.C.; Pardi, A.; Polisky, B. High-resolution molecular discrimination by RNA. Science 1994, 263, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Sassanfar, M.; Szostak, J.W. An RNA motif that binds atp. Nature 1993, 364, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Lister, N.; Shevchenko, G.; Walshe, J.L.; Groen, J.; Johnsson, P.; Vidarsdóttir, L.; Grander, D.; Ataide, S.F.; Morris, K.V. The molecular dynamics of long noncoding RNA control of transcription in pten and its pseudogene. Proc. Natl. Acad. Sci. USA 2017, 114, 9942–9947. [Google Scholar] [CrossRef] [PubMed]
- Di Ruscio, A.; Ebralidze, A.K.; Benoukraf, T.; Amabile, G.; Goff, L.A.; Terragni, J.; Figueroa, M.E.; De Figueiredo Pontes, L.L.; Alberich-Jorda, M.; Zhang, P.; et al. Dnmt1-interacting RNAs block gene-specific DNA methylation. Nature 2013, 503, 371–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catuogno, S.; Esposito, C.L.; Ungaro, P.; De Franciscis, V. Nucleic Acid Aptamers Targeting Epigenetic Regulators: An Innovative Therapeutic Option. Pharmaceuticals 2018, 11, 79. https://doi.org/10.3390/ph11030079
Catuogno S, Esposito CL, Ungaro P, De Franciscis V. Nucleic Acid Aptamers Targeting Epigenetic Regulators: An Innovative Therapeutic Option. Pharmaceuticals. 2018; 11(3):79. https://doi.org/10.3390/ph11030079
Chicago/Turabian StyleCatuogno, Silvia, Carla Lucia Esposito, Paola Ungaro, and Vittorio De Franciscis. 2018. "Nucleic Acid Aptamers Targeting Epigenetic Regulators: An Innovative Therapeutic Option" Pharmaceuticals 11, no. 3: 79. https://doi.org/10.3390/ph11030079
APA StyleCatuogno, S., Esposito, C. L., Ungaro, P., & De Franciscis, V. (2018). Nucleic Acid Aptamers Targeting Epigenetic Regulators: An Innovative Therapeutic Option. Pharmaceuticals, 11(3), 79. https://doi.org/10.3390/ph11030079