Functional Anatomical Changes in Ulcerative Colitis Patients Determine Their Gut Microbiota Composition and Consequently the Possible Treatment Outcome

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
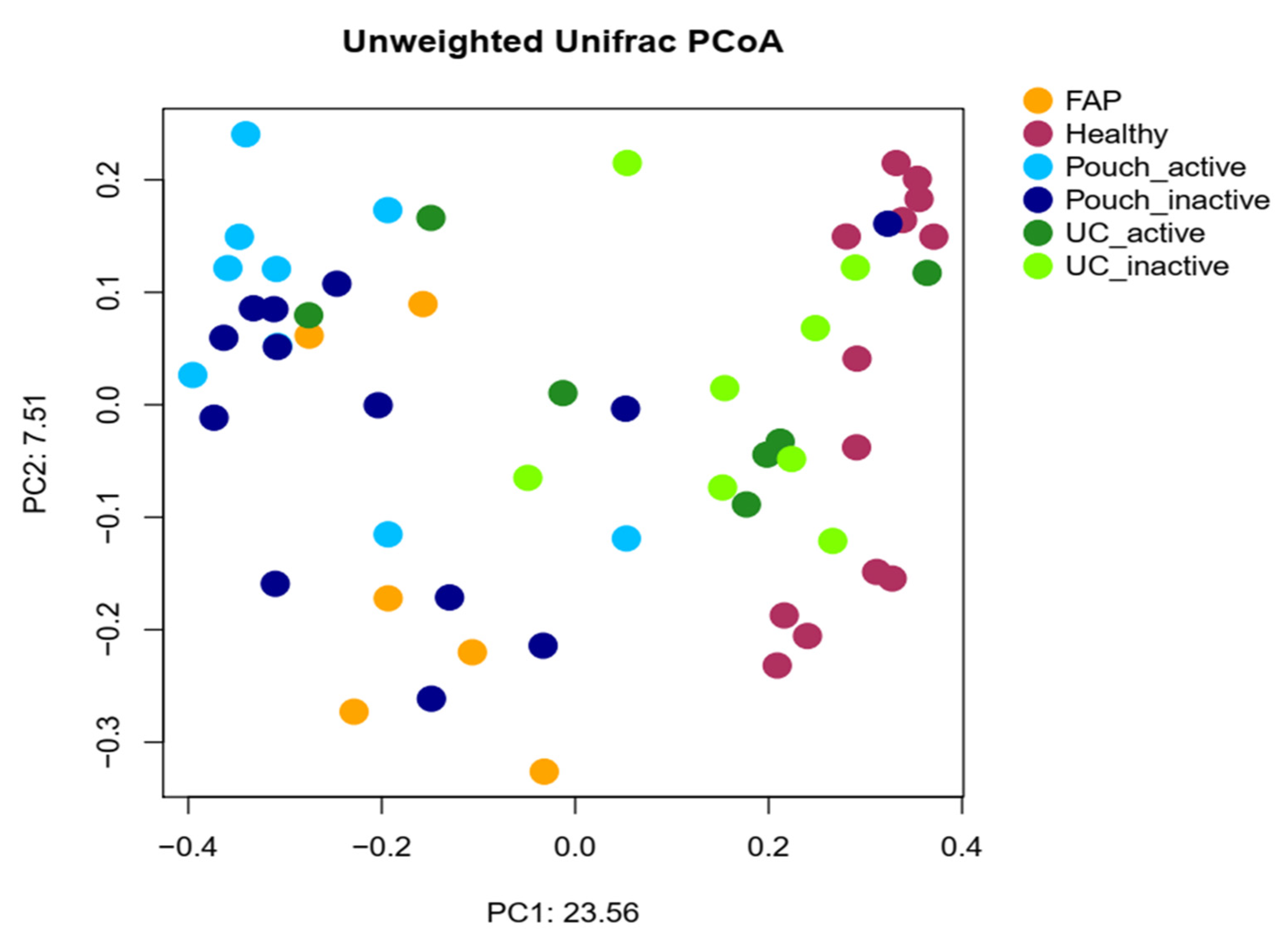
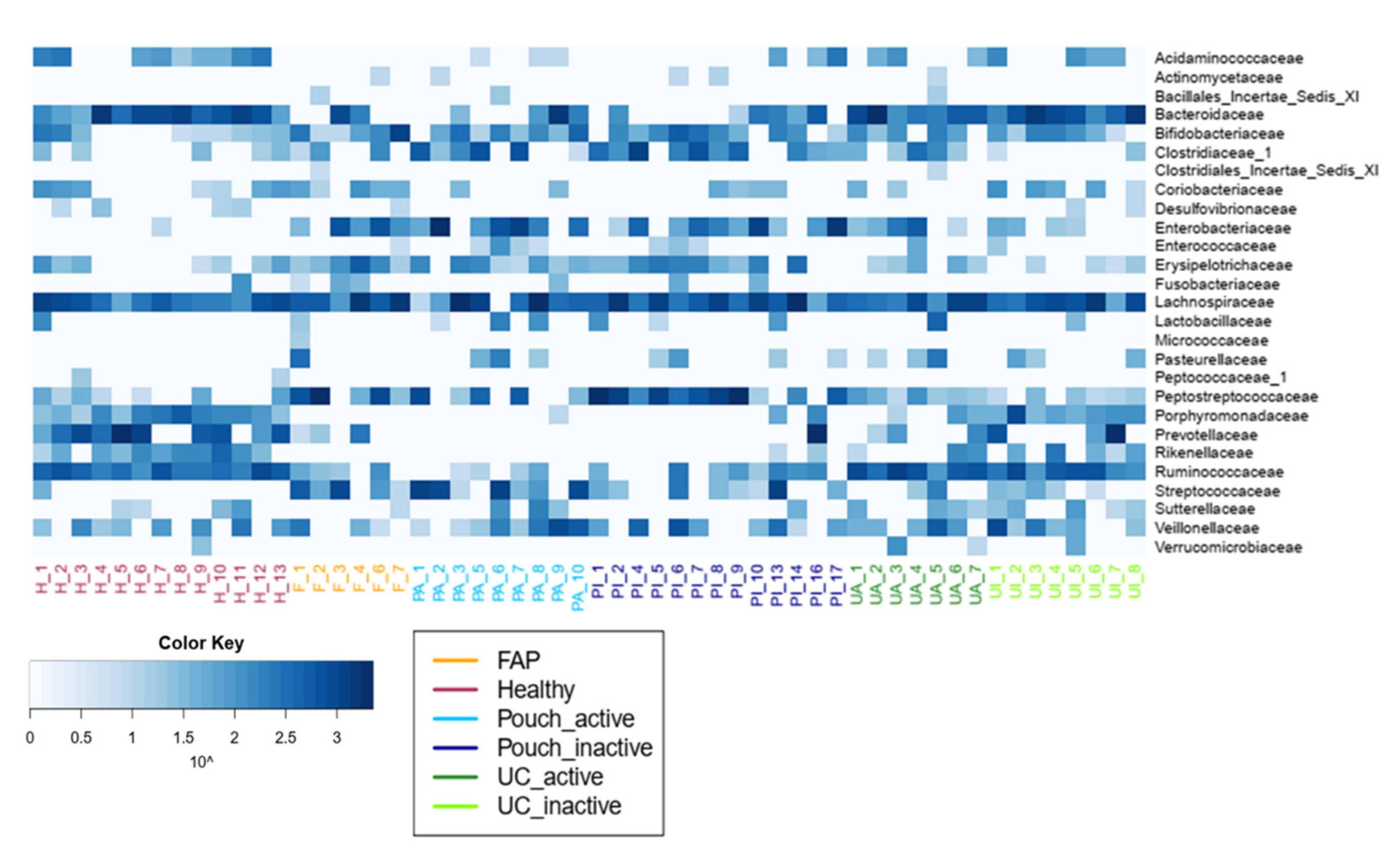
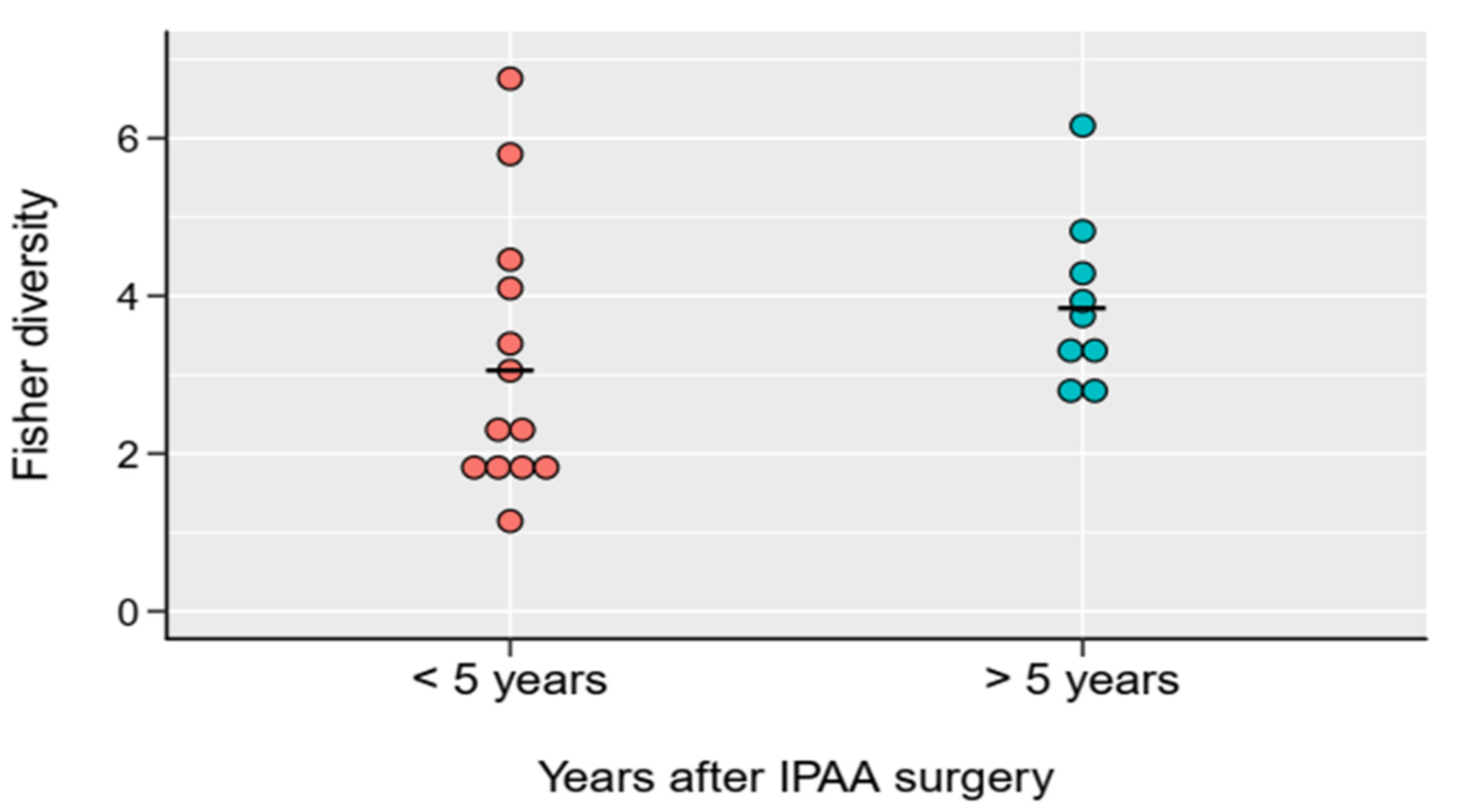
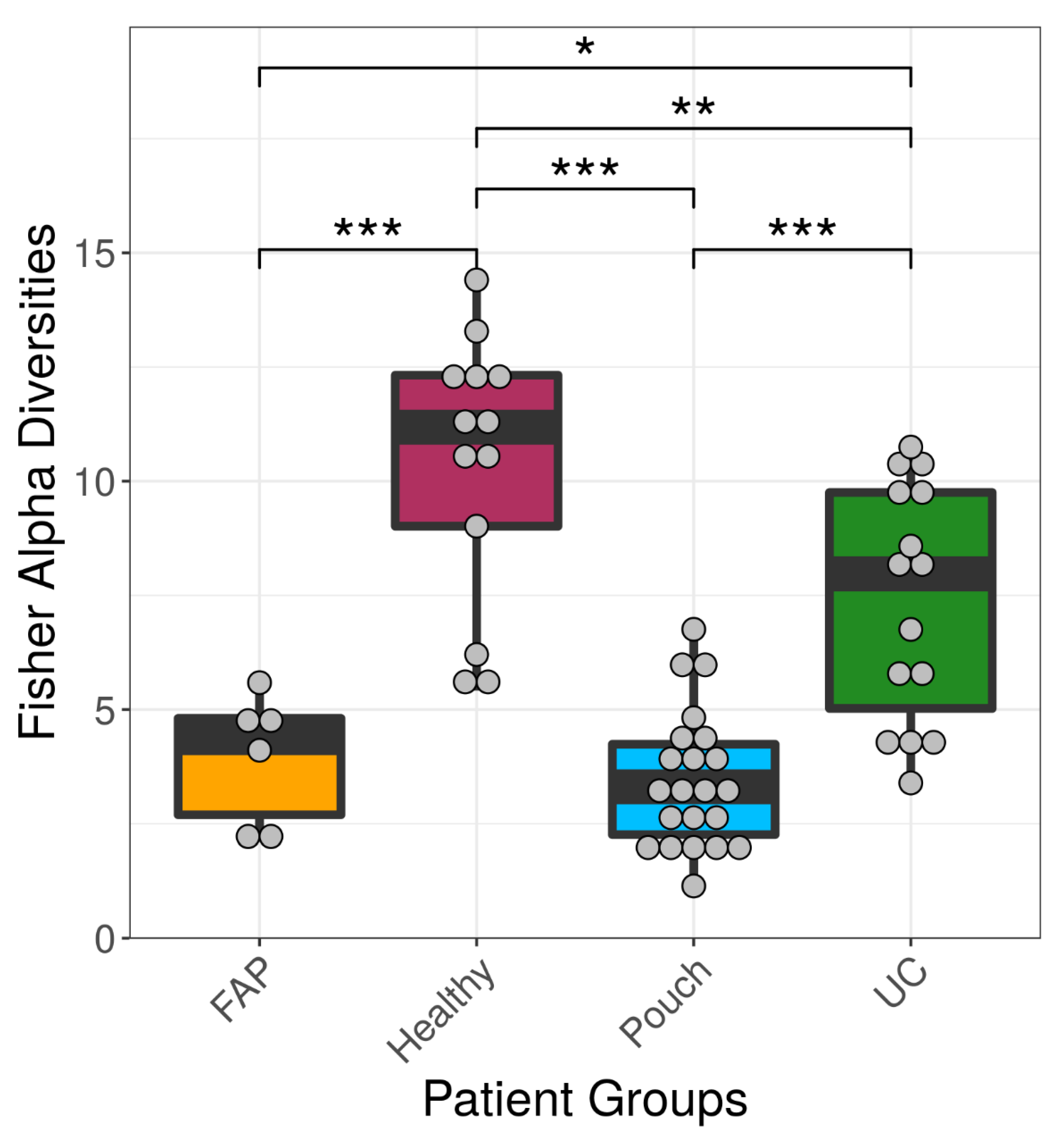
2. Results
2.1. Demographics and Clinical Data
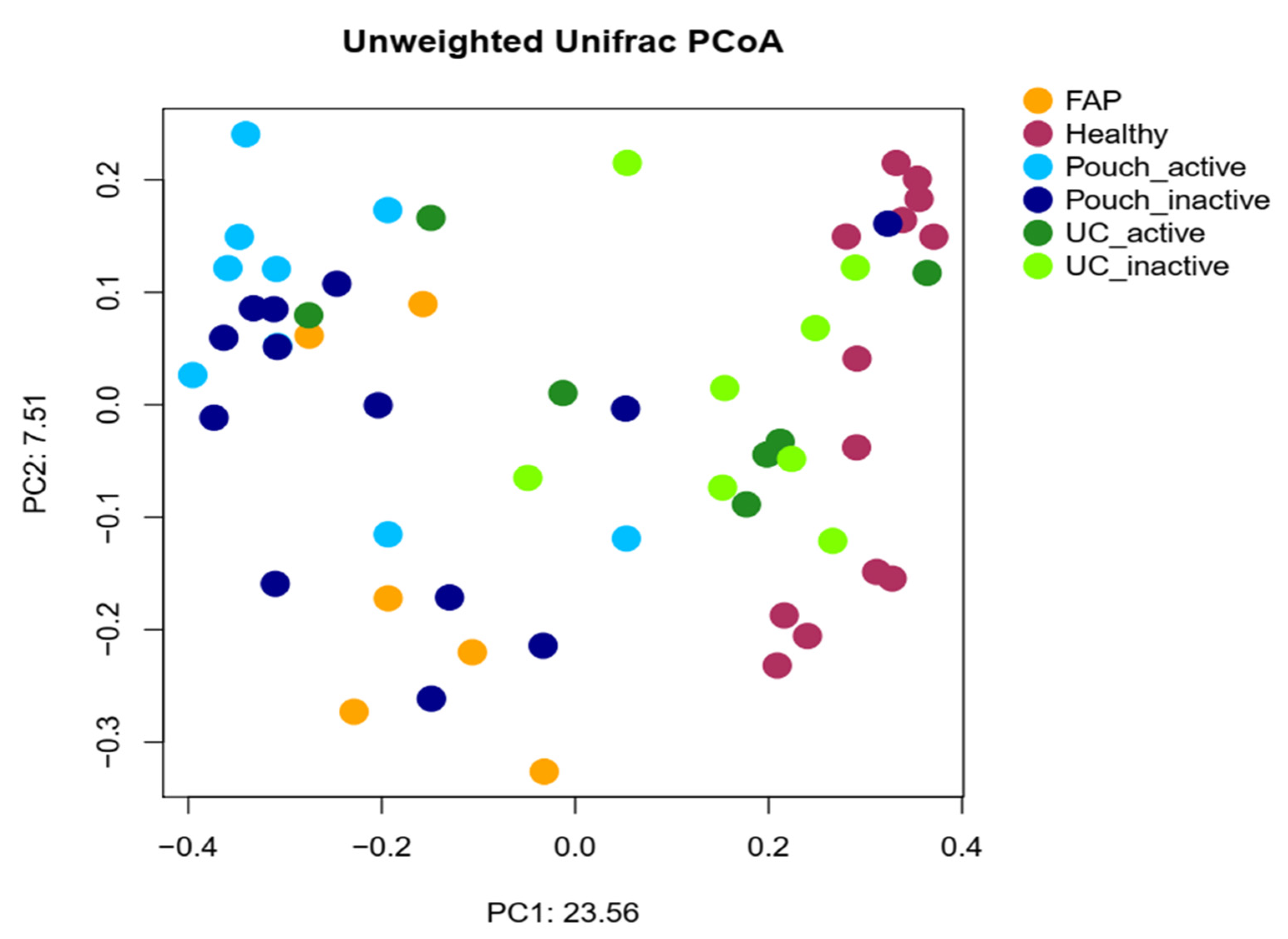
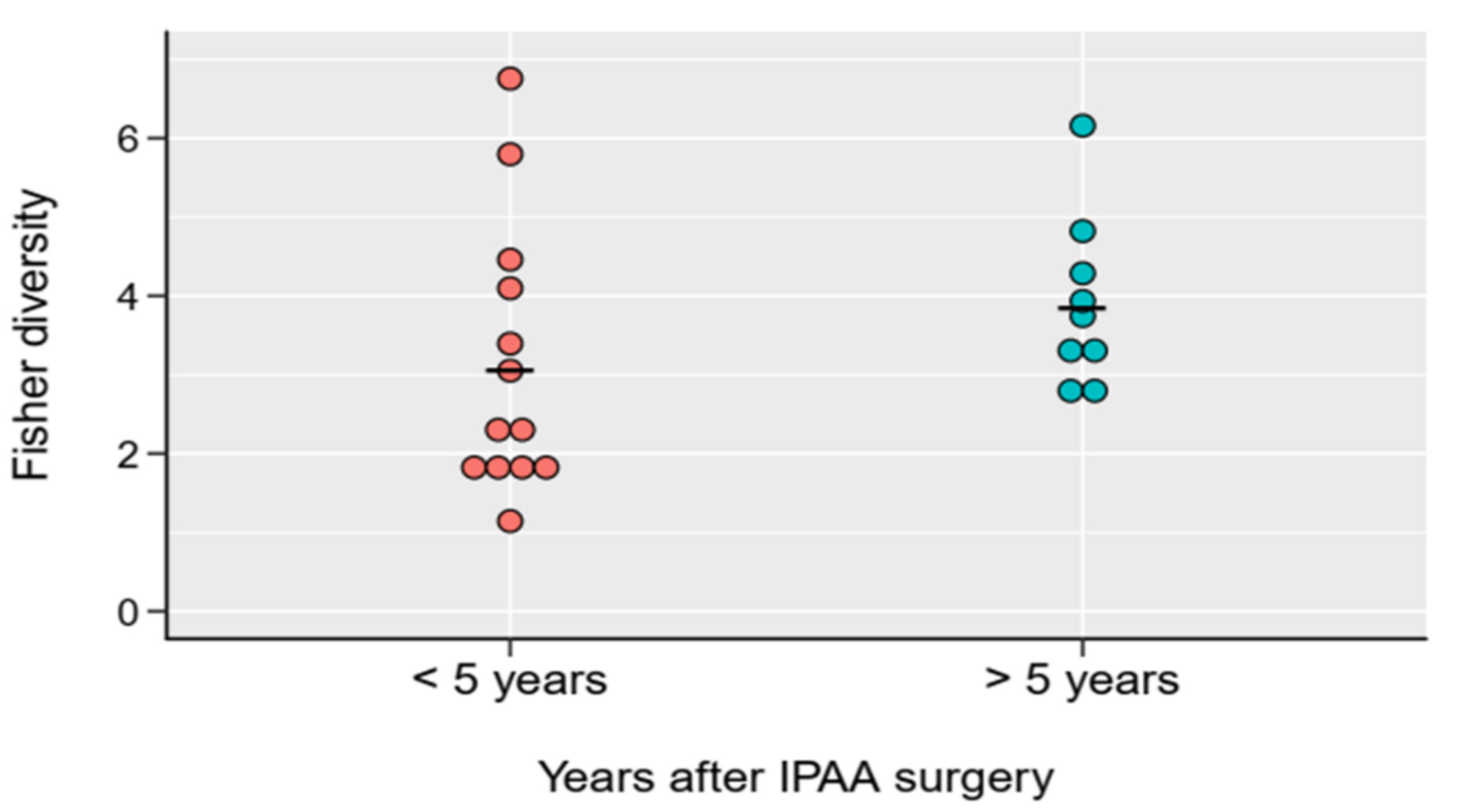
2.2. Bacterial Diversity of the Examined Groups
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Definitions
4.3. Specimen Collection and Storage
4.4. 16S rRNA-Based Bacterial Community Profiling
4.5. Ethics
4.6. Data analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zuo, T.; Ng, S.C. The Gut Microbiota in the Pathogenesis and Therapeutics of Inflammatory Bowel Disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787. [Google Scholar] [CrossRef] [PubMed]
- Lupp, C.; Robertson, M.L.; Wickham, M.E.; Sekirov, I.; Champion, O.L.; Gaynor, E.C.; Finlay, B.B. Host-Mediated Inflammation Disrupts the Intestinal Microbiota and Promotes the Overgrowth of Enterobacteriaceae. Cell Host Microbe 2007, 2, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naftali, T.; Reshef, L.; Kovacs, A.; Porat, R.; Amir, I.; Konikoff, F.M.; Gophna, U. Distinct Microbiotas are Associated with Ileum-Restricted and Colon-Involving Crohnʼs Disease. Inflamm. Bowel Dis. 2016, 22, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Segal, J.P.; Oke, S.; Hold, G.L.; Clark, S.K.; Faiz, O.D.; Hart, A.L. Systematic review: Ileoanal pouch microbiota in health and disease. Aliment. Pharmacol. Ther. 2018, 47, 466–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machiels, K.; Sabino, J.; Vandermosten, L.; Joossens, M.; Arijs, I.; de Bruyn, M.; Eeckhaut, V.; van Assche, G.; Ferrante, M.; Verhaegen, J.; et al. Specific members of the predominant gut microbiota predict pouchitis following colectomy and IPAA in UC. Gut 2017, 66, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Young, V.B.; Raffals, L.H.; Huse, S.M.; Vital, M.; Dai, D.; Schloss, P.D.; Brulc, J.M.; Antonopoulos, D.A.; Arrieta, R.L.; Kwon, J.H.; et al. Multiphasic analysis of the temporal development of the distal gut microbiota in patients following ileal pouch anal anastomosis. Microbiome 2013, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Halfvarson, J.; Brislawn, C.J.; Lamendella, R.; Vazquez-Baeza, Y.; Walters, W.A.; Bramer, L.M.; D’amato, M.; Bonfiglio, F.; McDonald, D.; Gonzalez, A.; et al. Dynamics of the human gut microbiome in inflammatory bowel disease. Nat. Microbiol. 2017, 2, 17004. [Google Scholar] [CrossRef] [Green Version]
- Magro, F.; Gionchetti, P.; Eliakim, R.; Ardizzone, S.; Armuzzi, A.; Barreiro-de Acosta, M.; Burisch, J.; Gecse, K.B.; Hart, A.L.; Hindryckx, P.; et al. Third European Evidence-based Consensus on Diagnosis and Management of Ulcerative Colitis. Part 1: Definitions, Diagnosis, Extra-intestinal Manifestations, Pregnancy, Cancer Surveillance, Surgery, and Ileo-anal Pouch Disorders. J. Crohns Colitis 2017, 11, 649–670. [Google Scholar] [CrossRef]
- Reshef, L.; Kovacs, A.; Ofer, A.; Yahav, L.; Maharshak, N.; Keren, N.; Konikoff, F.M.; Tulchinsky, H.; Gophna, U.; Dotan, I. Pouch Inflammation Is Associated With a Decrease in Specific Bacterial Taxa. Gastroenterology 2015, 149, 718–727. [Google Scholar] [CrossRef]
- Li, K.-Y.; Wang, J.-L.; Wei, J.-P.; Gao, S.; Zhang, Y.; Wang, L.; Liu, G. Fecal microbiota in pouchitis and ulcerative colitis. World J. Gastroenterol. 2016, 22, 8929. [Google Scholar] [CrossRef] [PubMed]
- Tyler, A.D.; Knox, N.; Kabakchiev, B.; Milgrom, R.; Kirsch, R.; Cohen, Z.; McLeod, R.S.; Guttman, D.S.; Krause, D.O.; Silverberg, M.S. Characterization of the Gut-Associated Microbiome in Inflammatory Pouch Complications Following Ileal Pouch-Anal Anastomosis. PLoS ONE 2013, 8, e66934. [Google Scholar] [CrossRef] [PubMed]
- Vasapolli, R.; Schütte, K.; Schulz, C.; Vital, M.; Schomburg, D.; Pieper, D.H.; Vilchez-Vargas, R.; Malfertheiner, P. Analysis of Transcriptionally Active Bacteria Throughout the Gastrointestinal Tract of Healthy Individuals. Gastroenterology 2019, 157, 1081–1092. [Google Scholar] [CrossRef] [Green Version]
- Willing, B.P.; Dicksved, J.; Halfvarson, J.; Andersson, A.F.; Lucio, M.; Zheng, Z.; Jarnerot, G.; Tysk, C.; Jansson, J.K.; Engstrand, L. A Pyrosequencing Study in Twins Shows That Gastrointestinal Microbial Profiles Vary With Inflammatory Bowel Disease Phenotypes. Gastroenterology 2010, 139, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shachar, S.; Yanai, H.; Baram, L.; Elad, H.; Meirovithz, E.; Ofer, A.; Brazowski, E.; Tulchinsky, H.; Pasmanik-Chor, M.; Dotan, I. Gene Expression Profiles of Ileal Inflammatory Bowel Disease Correlate with Disease Phenotype and Advance Understanding of Its Immunopathogenesis. Inflamm. Bowel Dis. 2013, 19, 2509–2521. [Google Scholar] [CrossRef]
- Angriman, I. Relationship between pouch microbiota and pouchitis following restorative proctocolectomy for ulcerative colitis. World J. Gastroenterol. 2014, 20, 9665. [Google Scholar] [CrossRef] [PubMed]
- Maharshak, N.; Cohen, N.A.; Reshef, L.; Tulchinsky, H.; Gophna, U.; Dotan, I. Alterations of Enteric Microbiota in Patients with a Normal Ileal Pouch Are Predictive of Pouchitis. J. Crohns Colitis 2017, 11, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.W.; Rogers, G.B.; Bruce, K.D.; Lilley, A.K.; von Herbay, A.; Forbes, A.; Ciclitira, P.J.; Nicholls, J.R. Bacterial community diversity in cultures derived from healthy and inflamed ileal pouches after restorative proctocolectomy. Inflamm. Bowel Dis. 2009, 15, 1803–1811. [Google Scholar] [CrossRef]
- Lim, M.; Adams, J.D.W.; Wilcox, M.; Finan, P.; Sagar, P.; Burke, D. An Assessment of Bacterial Dysbiosis in Pouchitis Using Terminal Restriction Fragment Length Polymorphisms of 16S Ribosomal DNA From Pouch Effluent Microbiota. Dis. Colon. Rectum. 2009, 52, 1492–1500. [Google Scholar] [CrossRef]
- Forbes, J.D.; Van Domselaar, G.; Bernstein, C.N. Microbiome Survey of the Inflamed and Noninflamed Gut at Different Compartments Within the Gastrointestinal Tract of Inflammatory Bowel Disease Patients. Inflamm. Bowel Dis. 2016, 22, 817–825. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, S.D.; Walker, A.W.; Churcher, C.; Clark, S.K.; Tekkis, P.P.; Johnson, M.W.; Parkhill, J.; Ciclitira, P.J.; Dougan, G.; Nicholls, R.J.; et al. The Bacteriology of Pouchitis: A Molecular Phylogenetic Analysis Using 16S rRNA Gene Cloning and Sequencing. Ann. Surg. 2010, 252, 90–98. [Google Scholar] [CrossRef]
- Contijoch, E.J.; Britton, G.J.; Yang, C.; Mogno, I.; Li, Z.; Ng, R.; Llewellyn, S.R.; Hira, S.; Johnson, C.; Rabinowitz, K.M.; et al. Gut microbiota density influences host physiology and is shaped by host and microbial factors. eLife 2019, 8, e40553. [Google Scholar] [CrossRef]
- Tropini, C.; Earle, K.A.; Huang, K.C.; Sonnenburg, J.L. The Gut Microbiome: Connecting Spatial Organization to Function. Cell Host Microbe 2017, 21, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohyama, A.; Ogawa, H.; Funayama, Y.; Takahashi, K.; Benno, Y.; Nagasawa, K.; Tomita, S.; Sasaki, I.; Fukushima, K. Bacterial population moves toward a colon-like community in the pouch after total proctocolectomy. Surgery 2009, 145, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Hinata, M.; Kohyama, A.; Ogawa, H.; Haneda, S.; Watanabe, K.; Suzuki, H.; Shibata, C.; Funayama, Y.; Takahashi, K.; Sasaki, I.; et al. A Shift from Colon- to Ileum-Predominant Bacteria in Ileal-Pouch Feces Following Total Proctocolectomy. Dig. Dis. Sci. 2012, 57, 2965–2974. [Google Scholar] [CrossRef] [PubMed]
- Oka, A.; Sartor, R.B. Microbial-Based and Microbial-Targeted Therapies for Inflammatory Bowel Diseases. Dig. Dis. Sci. 2020, 65, 757–788. [Google Scholar] [CrossRef] [Green Version]
- Jadhav, P.; Jiang, Y.; Jarr, K.; Layton, C.; Ashouri, J.F.; Sinha, S.R. Efficacy of Dietary Supplements in Inflammatory Bowel Disease and Related Autoimmune Diseases. Nutrients 2020, 12, 2156. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Tremaine, W.J.; Batts, K.P.; Pemberton, J.H.; Phillips, S.F. Pouchitis After Ileal Pouch-Anal Anastomosis: A Pouchitis Disease Activity Index. Mayo Clin. Proc. 1994, 69, 409–415. [Google Scholar] [CrossRef]
- Lewis, J.D.; Chuai, S.; Nessel, L.; Lichtenstein, G.R.; Aberra, F.N.; Ellenberg, J.H. Use of the noninvasive components of the mayo score to assess clinical response in Ulcerative Colitis. Inflamm. Bowel Dis. 2008, 14, 1660–1666. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Kintses, B.; Méhi, O.; Ari, E.; Szamel, M.; Gyorkei, A.; Jangir, P.K.; Nagy, I.; Pal, F.; Fekete, G.; Tengolics, R.; et al. Phylogenetic barriers to horizontal transfer of antimicrobial peptide resistance genes in the human gut microbiota. Nat. Microbiol. 2019, 4, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Rettedal, E.A.; Gumpert, H.; Sommer, M.O.A. Cultivation-based multiplex phenotyping of human gut microbiota allows targeted recovery of previously uncultured bacteria. Nat. Commun. 2014, 5, 4714. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic. Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The relation between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravorty, S.; Helb, D.; Burday, M.; Connell, N.; Alland, D. A detailed analysis of 16S ribosomal RNA gene segments for the diagnosis of pathogenic bacteria. J. Microbiol. Methods 2007, 69, 330–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, V.; Österlund, T.; Nerman, O.; Kristiansson, E. Statistical evaluation of methods for identification of differentially abundant genes in comparative metagenomics. BMC Genom. 2016, 17, 78. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ulcerative colitis (UC) | UC-Pouch (P) | FAP (F) | Healthy (H) | |||||
|---|---|---|---|---|---|---|---|---|
| Active | Inactive | Total | Active | Inactive | Total | |||
| No. of patients | 7 | 8 | 15 | 9 | 13 | 22 | 6 | 13 |
| Gender (male/female), No. of patients | 6/1 | 5/3 | 11/4 | 4/5 | 8/5 | 12/10 | 3/3 | 6/7 |
| age at inclusion (SD, years) | 48.6 (21.7) | 45.1 (14.6) | 46.7 (17.6) | 51.5 (13.9) | 41.3 (12.8) | 45(13.9) | 31.7 (6.5) | 32.35 (7.45) |
| disease duration at inclusion (SD, years) | 10.9 (11.1) | 11.4 (9) | 11.1 (9.6) | 14 (6.7) | 15.1 (10.5) | 15 (9) | 11.9 (7.3) | NA |
| time after IPAA surgery (SD, years) | NA | NA | NA | 4.6 (4.2) | 5 (5) | 5 (4.6) | 8.7 (7.4) | NA |
| Therapy (No. of patients) | ||||||||
| none | 1 | 0 | 1 | 2 | 9 | 11 | 6 | 13 |
| oral 5-ASA | 3 | 6 | 9 | 0 | 0 | 0 | 0 | 0 |
| topical 5-ASA | 1 | 3 | 4 | 2 | 0 | 2 | 0 | 0 |
| oral corticosteroid | 3 | 2 | 5 | 1 | 0 | 1 | 0 | 0 |
| topical corticosteroid | 2 | 2 | 4 | 0 | 0 | 0 | 0 | 0 |
| azathioprine | 2 | 4 | 6 | 0 | 0 | 0 | 0 | 0 |
| biological therapy (IFX, ADA) | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Laboratory parameters | ||||||||
| faecalcalprotectin (median, min–max, μg/g) | 1000 (116.4–1000) | 303.4 (12.6–1000) | 360.2 (12.6–1000) | 1000 (38.8–1800) | 428 (100–1222) | 691.7 (38.8–1800) | 284.8 (145.1–711.7) | 11.8 (6.9–20.5) |
| CRP (mean, SD, mg/L) | 17.8 (17.5) | 7.7 (5.1) | 12.7 (13.5) | 9.3 (5.1) | 7.3 (4.6) | 7.8 (4.6) | 2.1 (0.2) | NA |
| serum iron (mean, SD, μmol/L) | 10.9 (7.3) | 16.6 (5.7) | 13.7 (7) | 9.1 (8.5) | 12 (6.8) | 11 (7.3) | NA | NA |
| haematocrit (mean, SD, L/L) | 39 (5.5) | 42.3 (5) | 40.6 (5.3) | 38 (4.4) | 42.9 (3.5) | 41(4.5) | 42 (3.3) | NA |
| thrombocytes (mean, SD, G/L) | 335.4 (93.4) | 262 (51.4) | 298.7 (81.9) | 349 (121.1) | 291.3 (73.5) | 315.6 (97.8) | 309.5 (162.7) | NA |
| albumin (mean, SD, g/L) | 42.4 (5.3) | 47.3 (3.7) | 44.9 (5.1) | 42 (6.9) | 46.8 (2.9) | 45.3 (4.8) | 41.5 (2.1) | NA |
| Compared Groups | Higher or Lower Abundance Group | Taxonomic Annotation | Adjusted p Value |
|---|---|---|---|
| Healthy vs. Pouch active | less abundant in Pouch active | Acidaminococcaceae | 0.032 |
| Porphyromonadaceae | 0.007 | ||
| Prevotellaceae | <0.001 | ||
| Rikenellaceae | <0.001 | ||
| Ruminococcaceae | 0.007 | ||
| more abundant in Pouch active | Actinomycetaceae | 0.040 | |
| Clostridiaceae | 0.016 | ||
| Enterobacteriaceae | <0.001 | ||
| Enterococcaceae | 0.011 | ||
| Pasteurellaceae | 0.007 | ||
| Streptococcaceae | <0.001 | ||
| Healthy vs. Pouch inactive | less abundant in Pouch inactive | Bacteroidaceae | 0.018 |
| Desulfovibrionaceae | 0.018 | ||
| Porphyromonadaceae | 0.018 | ||
| Rikenellaceae | 0.002 | ||
| Ruminococcaceae | 0.037 | ||
| more abundant in Pouch inactive | Clostridiaceae | 0.018 | |
| Enterobacteriaceae | <0.001 | ||
| Pasteurellaceae | 0.018 | ||
| Peptostreptococcaceae | 0.004 | ||
| Streptococcaceae | 0.018 | ||
| Pouch active vs. UC inactive | less abundant in UC inactive | Clostridiaceae_1 | 0.04 |
| more abundant in UC inactive | Acidaminococcaceae | 0.01 | |
| Porphyromonadaceae | <0.001 | ||
| Prevotellaceae | <0.001 | ||
| Rikenellaceae | <0.001 | ||
| Ruminococcaceae | <0.001 | ||
| Pouch active vs. UC active | more abundant in UC active | Prevotellaceae | 0.015 |
| Rikenellaceae | 0.020 | ||
| Ruminococcaceae | 0.015 | ||
| Verrucomicrobiaceae | 0.015 | ||
| Pouch inactive vs. UC active | more abundant in UC active | Verrucomicrobiaceae | 0.031 |
| Pouch inactive vs. UC inactive | less abundant in UC inactive | Peptostreptococcaceae | 0.029 |
| Healthy vs. UC active | more abundant in UC active | Pasteurellaceae | 0.011 |
| Enterobacteriaceae | <0.001 | ||
| Enterococcaceae | 0.041 | ||
| Healthy vs. UC inactive | more abundant in UC inactive | Pasteurellaceae | 0.042 |
| Bacillales_Incertae_Sedis_XI | 0.039 | ||
| Enterobacteriaceae | <0.001 | ||
| Micrococcaceae | 0.049 | ||
| Pasteurellaceae | <0.001 | ||
| Peptostreptococcaceae | 0.001 | ||
| Streptococcaceae | 0.001 | ||
| more abundant in healthy | Acidaminococcaceae | 0.004 | |
| Porphyromonadaceae | 0.002 | ||
| FAP vs. UC inactive | less abundant in UC inactive | Peptostreptococcaceae | 0.019 |
| more abundant in UC inactive | Porphyromonadaceae | 0.009 | |
| Rikenellaceae | 0.040 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bálint, A.; Farkas, K.; Méhi, O.; Kintses, B.; Vásárhelyi, B.M.; Ari, E.; Pál, C.; Madácsy, T.; Maléth, J.; Szántó, K.J.; et al. Functional Anatomical Changes in Ulcerative Colitis Patients Determine Their Gut Microbiota Composition and Consequently the Possible Treatment Outcome. Pharmaceuticals 2020, 13, 346. https://doi.org/10.3390/ph13110346
Bálint A, Farkas K, Méhi O, Kintses B, Vásárhelyi BM, Ari E, Pál C, Madácsy T, Maléth J, Szántó KJ, et al. Functional Anatomical Changes in Ulcerative Colitis Patients Determine Their Gut Microbiota Composition and Consequently the Possible Treatment Outcome. Pharmaceuticals. 2020; 13(11):346. https://doi.org/10.3390/ph13110346
Chicago/Turabian StyleBálint, Anita, Klaudia Farkas, Orsolya Méhi, Bálint Kintses, Bálint Márk Vásárhelyi, Eszter Ari, Csaba Pál, Tamara Madácsy, József Maléth, Kata Judit Szántó, and et al. 2020. "Functional Anatomical Changes in Ulcerative Colitis Patients Determine Their Gut Microbiota Composition and Consequently the Possible Treatment Outcome" Pharmaceuticals 13, no. 11: 346. https://doi.org/10.3390/ph13110346