Can Leaves and Stems of Rubus idaeus L. Handle Candida albicans Biofilms?

 ,
,
Abstract
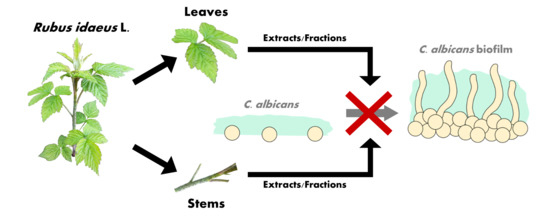
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
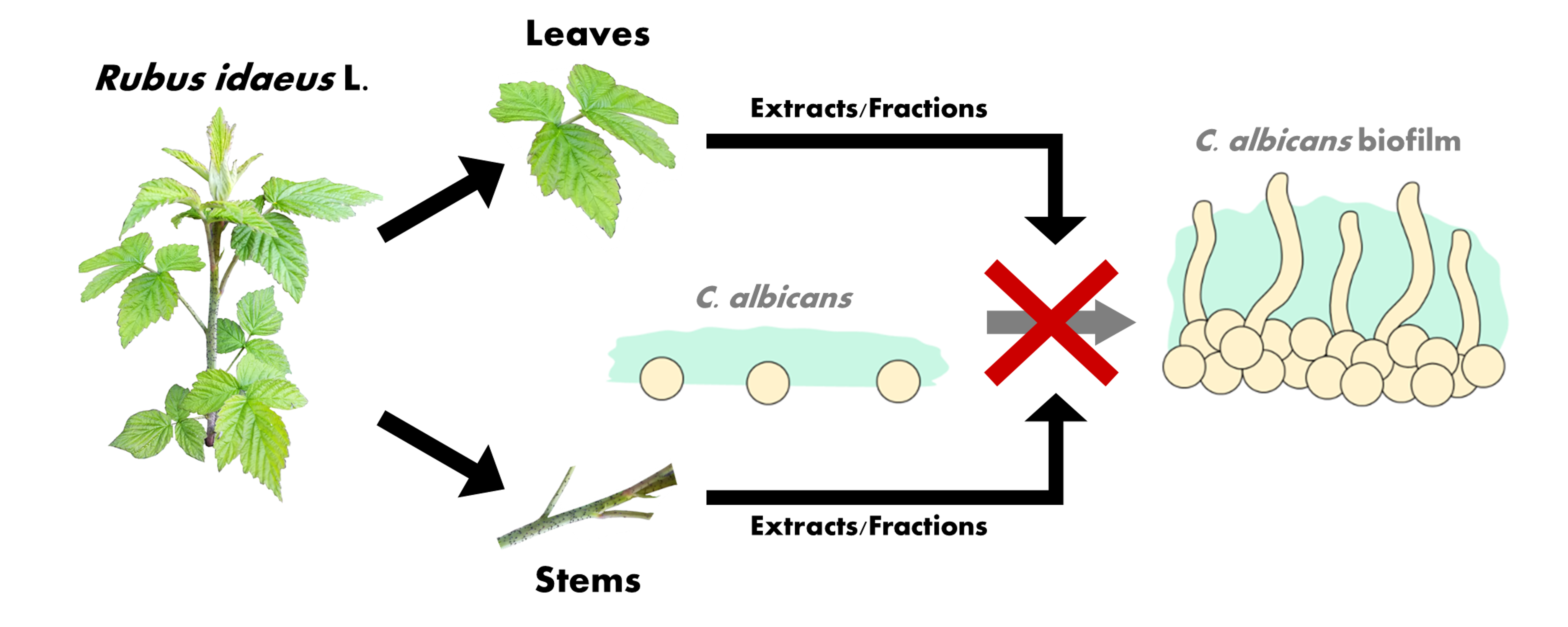
3.3. Preparation of Extracts
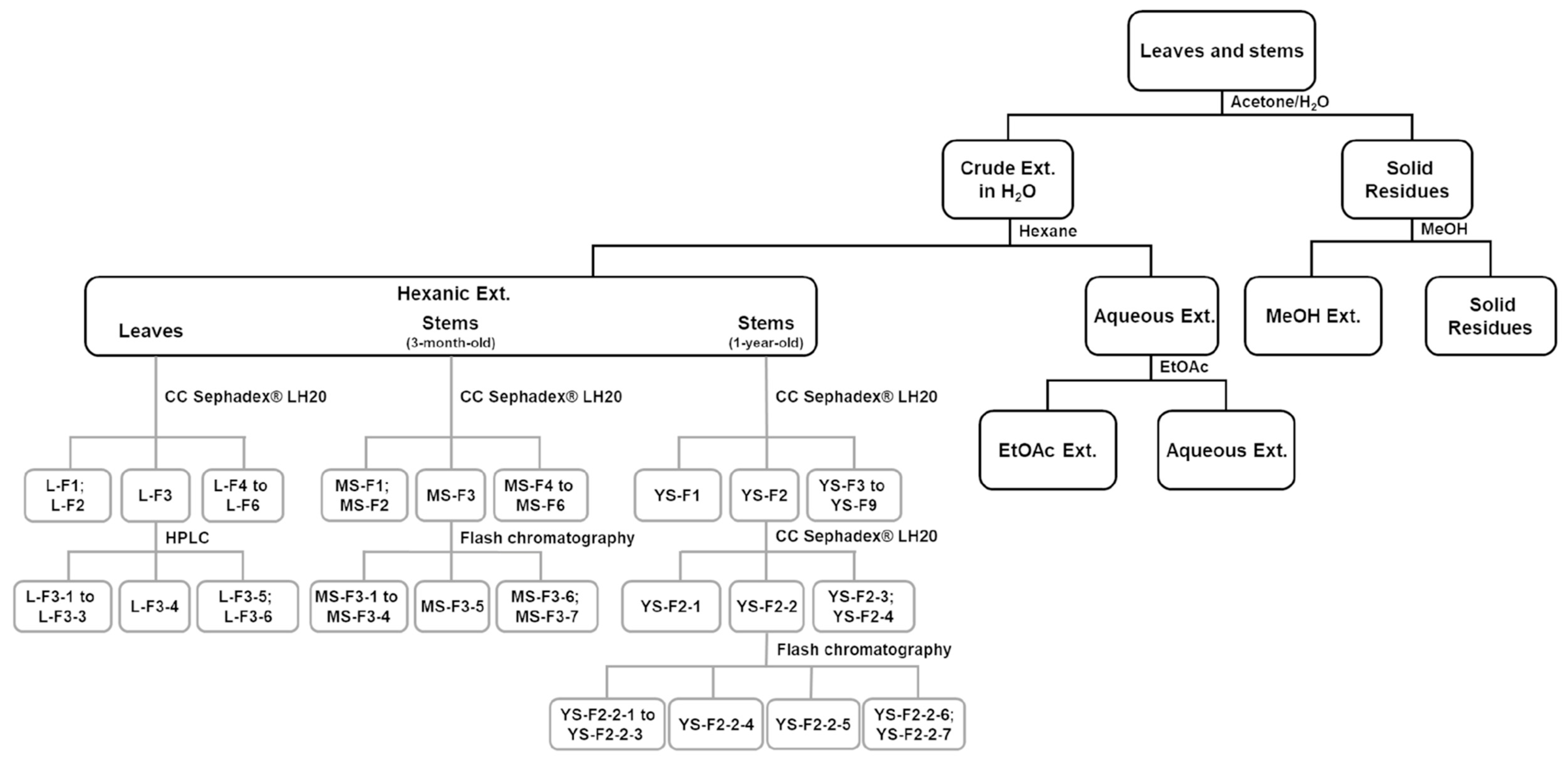
3.4. Fractionation of Active Extracts
3.5. Anti-Biofilm Growth Test
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Sudbery, P. Candida albicans, a major human fungal pathogen. J. Microbiol. 2011, 49, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.M.; George, T.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Antifungal susceptibility of Candida biofilms: Unique efficacy of amphotericin B lipid formulations and echinocandins. Antimicrob. Agents Chemother. 2002, 46, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Chandra, J. Candida biofilm resistance. Drug Resist. Updat. 2004, 7, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Kojic, E.M.; Darouiche, R.O. Candida infections of medical devices. Clin. Microbiol. Rev. 2004, 17, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Moser, C.; Bassi, G.L.; Coenye, T.; Donelli, G.; Hall-Stoodley, L.; Holá, V.; Imbert, C.; Kirketerp-Møller, K.; et al. ESCMID* guideline for the diagnosis and treatment of biofilm infections 2014. Clin. Microbiol. Infect. 2015, 21, S1–S25. [Google Scholar] [CrossRef] [PubMed]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef]
- Ghannoum, M.; Roilides, E.; Katragkou, A.; Petraitis, V.; Walsh, T.J. The role of echinocandins in Candida biofilm-related vascular catheter infections: In vitro and in vivo Model Systems. Clin. Infect. Dis. 2015, 61, S618–S621. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Perlin, D.S. Echinocandin resistance: An emerging clinical problem? Curr. Opin. Infect. Dis. 2014, 27, 484–492. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Red raspberries and their bioactive polyphenols: Cardiometabolic and neuronal health links. Adv. Nutr. 2016, 7, 44–65. [Google Scholar] [CrossRef]
- Ryan, T.; Wilkinson, J.M.; Cavanagh, H.M.A. Antibacterial activity of raspberry cordial in vitro. Res. Vet. Sci. 2001, 71, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Četojević-Simin, D.D.; Velićanski, A.S.; Cvetković, D.D.; Markov, S.L.; Ćetković, G.S.; Tumbas Šaponjac, V.T.; Vulić, J.J.; Čanadanović-Brunet, J.M.; Djilas, S.M. Bioactivity of Meeker and Willamette raspberry (Rubus idaeus L.) pomace extracts. Food Chem. 2015, 166, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Dutreix, L.; Bernard, C.; Juin, C.; Imbert, C.; Girardot, M. Do raspberry extracts and fractions have antifungal or anti-adherent potential against Candida spp.? Int. J. Antimicrob. Agents 2018, 52, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Gudej, J.; Tomczyk, M. Determination of flavonoids, tannins and ellagic acid in leaves from Rubus L. species. Arch. Pharm. Res. 2004, 27, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Krauze-Baranowska, M.; Głód, D.; Kula, M.; Majdan, M.; Hałasa, R.; Matkowski, A.; Kozłowska, W.; Kawiak, A. Chemical composition and biological activity of Rubus idaeus shoots—A traditional herbal remedy of Eastern Europe. BMC Complement. Altern. Med. 2014, 14, 480. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.V.; Rojas-Vera, J.; Dacke, C.G. Therapeutic constituents and actions of Rubus species. Curr. Med. Chem. 2004, 11, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Ponder, A.; Hallmann, E. Phenolics and carotenoid contents in the leaves of different organic and conventional raspberry (Rubus idaeus L.) cultivars and their in vitro activity. Antioxidants 2019, 8, 458. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lin, H.S. Antioxidant activity in fruits and leaves of blackberry, raspberry, and strawberry varies with cultivar and developmental stage. J. Agric. Food Chem. 2000, 48, 140–146. [Google Scholar] [CrossRef]
- Shepherd, T.; Robertson, G.W.; Griffiths, D.W.; Birch, A.N.E. Epicuticular wax composition in relation to aphid infestation and resistance in red raspberry (Rubus idaeus L.). Phytochemistry 1999, 52, 1239–1254. [Google Scholar] [CrossRef]
- Ağalar, H.G.; Çiftçi, G.A.; Göger, F.; Kırımer, N. Activity guided fractionation of Arum italicum miller tubers and the LC/MS-MS profiles. Rec. Nat. Prod. 2018, 12, 64–75. [Google Scholar] [CrossRef]
- McDougall, G.J.; Allwood, J.W.; Pereira-Caro, G.; Brown, E.M.; Verrall, S.; Stewart, D.; Latimer, C.; McMullan, G.; Lawther, R.; O’Connor, G.; et al. Novel colon-available triterpenoids identified in raspberry fruits exhibit antigenotoxic activities in vitro. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Han, J.; Chen, H.; Zheng, J.; Guo, D. Analysis of phenolic compounds in rhubarbs using liquid chromatography coupled with electrospray ionization mass spectrometry. J. Am. Soc. Mass Spectrom. 2007, 18, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.T.; Wu, X.; Rui, W.; Guo, J.; Feng, Y.F. UPLC/Q-TOF-MS analysis for identification of hydrophilic phenolics and lipophilic diterpenoids from radix Salviae miltiorrhizae. Acta Chromatogr. 2015, 27, 711–728. [Google Scholar] [CrossRef]
- Kajdžanoska, M.; Gjamovski, V.; Stefova, M. HPLC-DAD-ESI-MSn identification of phenolic compounds in cultivated strawberries from Macedonia. Maced. J. Chem. Chem. Eng. 2010, 29, 181–194. [Google Scholar] [CrossRef]
- Jahan, N.; Ahmad, M.; Saeed, F.; Rehman, A.; Muhammad, S. Anti-nociceptive activity of seed extract of Vernonia anthelmintica willd. Pak. J. Pharm. Sci. 2014, 27, 2177–2181. [Google Scholar] [PubMed]
- Xu, Q.M.; Liu, Y.L.; Li, X.R.; Li, X.; Yang, S.L. Three new fatty acids from the roots of Boehmeria nivea (L.) Gaudich and their antifungal activities. Nat. Prod. Res. 2011, 25, 640–647. [Google Scholar] [CrossRef]
- Costea, T.; Vlase, L.; Gostin, I.N.; Olah, N.K.; Mihaela, G.; Predan, I. Botanical characterization, phytochemical analysis and antioxidant activity of indigenous red raspberry (Rubus idaeus L.) leaves. Studia Univ. Vasile Goldis Arad. Seria Stiintele Vietii 2016, 26, 463–472. [Google Scholar]
- Celik, F.; Ercisli, S. Lipid and fatty acid composition of wild and cultivated red raspberry fruits (Rubus idaeus L.). J. Med. Plants Res. 2009, 3, 583–585. [Google Scholar]
- Haddock, E.A.; Gupta, R.K.; Al-Shafi, S.M.K.; Haslam, E.; Magnolato, D. The metabolism of gallic acid and hexahydroxydiphenic acid in plants. Part 1. Introduction. Naturally occurring galloyl esters. J. Chem. Soc. Perkin Trans. 1 1982, 2515–2524. [Google Scholar] [CrossRef]
- Gudej, J. Kaempferol and quercetin glycosides from Rubus idaeus L. leaves. Acta Pol. Pharm. Drug Res. 2003, 60, 313–316. [Google Scholar]
- Zia-Ul-Haq, M.; Riaz, M.; De Feo, V.; Jaafar, H.Z.E.; Moga, M. Rubus fruticosus L.: Constituents, biological activities and health related uses. Molecules 2014, 19, 10998–11029. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Juárez, I.; García-Contreras, R.; Velázquez-Guadarrama, N.; Soto-Hernández, M.; Martínez-Vázquez, M. Amphypterygium adstringens anacardic acid mixture inhibits quorum sensing-controlled virulence factors of Chromobacterium violaceum and Pseudomonas aeruginosa. Arch. Med. Res. 2013, 44, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Sajeevan, S.E.; Chatterjee, M.; Paul, V.; Baranwal, G.; Kumar, V.A.; Bose, C.; Banerji, A.; Nair, B.G.; Prasanth, B.P.; Biswas, R. Impregnation of catheters with anacardic acid from cashew nut shell prevents Staphylococcus aureus biofilm development. J. Appl. Microbiol. 2018, 125, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.H.; Kock, J.L.F.; Thibane, V.S. Antifungal free fatty acids: A review. Sci. Against Microb. Pathog. Curr. Res. Technol. Adv. 2011, 1, 61–71. [Google Scholar]
- Zhou, L.; Ding, Y.; Chen, W.; Zhang, P.; Chen, Y.; Lv, X. The in vitro study of ursolic acid and oleanolic acid inhibiting cariogenic microorganisms as well as biofilm. Oral Dis. 2013, 19, 494–500. [Google Scholar] [CrossRef]
- Skalicka-Woźniak, K.; Los, R.; Głowniak, K.; Malm, A. Antimicrobial activity of fatty acids from fruits of Peucedanum cervaria and P. alsaticum. Chem. Biodivers. 2010, 7, 2748–2754. [Google Scholar] [CrossRef]
- Pandit, S.; Cai, J.N.; Song, K.Y.; Jeon, J.G. Identification of anti-biofilm components in Withania somnifera and their effect on virulence of Streptococcus mutans biofilms. J. Appl. Microbiol. 2015, 119, 571–581. [Google Scholar] [CrossRef]
- Ismail, S.; Jalilian, F.A.; Talebpour, A.H.; Zargar, M.; Shameli, K.; Sekawi, Z.; Jahanshiri, F. Chemical composition and antibacterial and cytotoxic activities of Allium hirtifolium boiss. BioMed Res. Int. 2013, 2013, 696835. [Google Scholar] [CrossRef]
- Rendeková, K.; Fialová, S.; Jánošová, L.; Mucaji, P.; Slobodníková, L. The activity of Cotinus coggygria scop. Leaves extract on Staphylococcus aureus strains in planktonic and biofilm growth forms. Molecules 2016, 21, 50. [Google Scholar] [CrossRef]
- Ming, D.; Wang, D.; Cao, F.; Xiang, H.; Mu, D.; Cao, J.; Li, B.; Zhong, L.; Dong, X.; Zhong, X.; et al. Kaempferol inhibits the primary attachment phase of biofilm formation in Staphylococcus aureus. Front. Microbiol. 2017, 8, 2263. [Google Scholar] [CrossRef]
- Freires, I.A.; Queiroz, V.C.P.P.; Furletti, V.F.; Ikegaki, M.; de Alencar, S.M.; Duarte, M.C.T.; Rosalen, P.L. Chemical composition and antifungal potential of Brazilian propolis against Candida spp. J. Mycol. Med. 2016, 26, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.I.; Lee, J.H.; Park, K.H.; Sangurdekar, D.; Chang, W.S. Effect of soybean coumestrol on Bradyrhizobium japonicum nodulation ability, biofilm formation, and transcriptional profile. Appl. Environ. Microbiol. 2012, 78, 2896–2903. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Regmi, S.C.; Kim, J.A.; Cho, M.H.; Yun, H.; Lee, C.S.; Lee, J. Apple flavonoid phloretin inhibits Escherichia coli O157:H7 biofilm formation and ameliorates colon inflammation in rats. Infect. Immun. 2011, 79, 4819–4827. [Google Scholar] [CrossRef] [PubMed]
- Brand, A. Hyphal growth in human fungal pathogens and its role in virulence. Int. J. Microbiol. 2012, 2012, 517529. [Google Scholar] [CrossRef] [PubMed]
- Clément, M.; Tremblay, J.; Lange, M.; Thibodeau, J.; Belhumeur, P. Whey-derived free fatty acids suppress the germination of Candida albicans in vitro. FEMS Yeast Res. 2007, 7, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Vediyappan, G.; Dumontet, V.; Pelissier, F.; D’Enfert, C. Gymnemic acids inhibit hyphal growth and virulence in Candida albicans. PLoS ONE 2013, 8, e74189. [Google Scholar] [CrossRef]
- Kazuko, O.S.; Sato, Y.; Azuma, T. Resveratrol impaired the morphological transition of Candida albicans under various hyphae-inducing conditions. J. Microbiol. Biotechnol. 2010, 20, 942–945. [Google Scholar] [CrossRef]
- Nicol, M.; Alexandre, S.; Luizet, J.B.; Skogman, M.; Jouenne, T.; Salcedo, S.P.; Dé, E. Unsaturated fatty acids affect quorum sensing communication system and inhibit motility and biofilm formation of Acinetobacter baumannii. Int. J. Mol. Sci. 2018, 19, 214. [Google Scholar] [CrossRef]
- Paul Bhattacharya, S.; Mitra, A.; Bhattacharya, A.; Sen, A. Quorum quenching activity of pentacyclic triterpenoids leads to inhibition of biofilm formation by Acinetobacter baumannii. Biofouling 2020, 36, 922–937. [Google Scholar] [CrossRef]
- Ilk, S.; Sağlam, N.; Özgen, M.; Korkusuz, F. Chitosan nanoparticles enhances the anti-quorum sensing activity of kaempferol. Int. J. Biol. Macromol. 2017, 94, 653–662. [Google Scholar] [CrossRef]
- Girardot, M.; Guerineau, A.; Boudesocque, L.; Costa, D.; Bazinet, L.; Enguehard-Gueiffier, C.; Imbert, C. Promising results of cranberry in the prevention of oral Candida biofilms. Pathog. Dis. 2014, 70, 432–439. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R. idaeus | Extracts | Weight (g) | Yield (%) | Anti-Biofilm Growth Activity IC50 (μg/mL) |
|---|---|---|---|---|
| Leaves | Hexane | 0.56 | 1.1 | 500 |
| EtOAc | 1.57 | 3.2 | 1000 | |
| MeOH | 0.88 | 1.8 | >2000 | |
| Aqueous | 5.08 | 10.2 | 2000 | |
| 3-month-old stems | Hexane | 0.23 | 0.5 | 500 |
| EtOAc | 0.62 | 1.3 | 2000 | |
| MeOH | 0.41 | 0.8 | 1000 | |
| Aqueous | 8.01 | 16 | 2000 | |
| 1-year-old stems | Hexane | 1.76 | 0.4 | 250 |
| EtOAc | 13.87 | 2.8 | >2000 | |
| MeOH | 2.65 | 0.5 | >2000 | |
| Aqueous | 58.93 | 11.8 | 1000 |
| R. idaeus | Fractions and Subfractions | Weight (mg) | Anti-Biofilm Growth Activity IC50 (μg/mL) |
|---|---|---|---|
| Leaves | L-F1; L-F2 | 50–80 | 200 |
| L-F3 | 40 | 50 | |
| L-F4 to L-F6 | 50–290 | ≥200 | |
| L-F3-1 to L-F3-3 | 5–7 | ≥250 | |
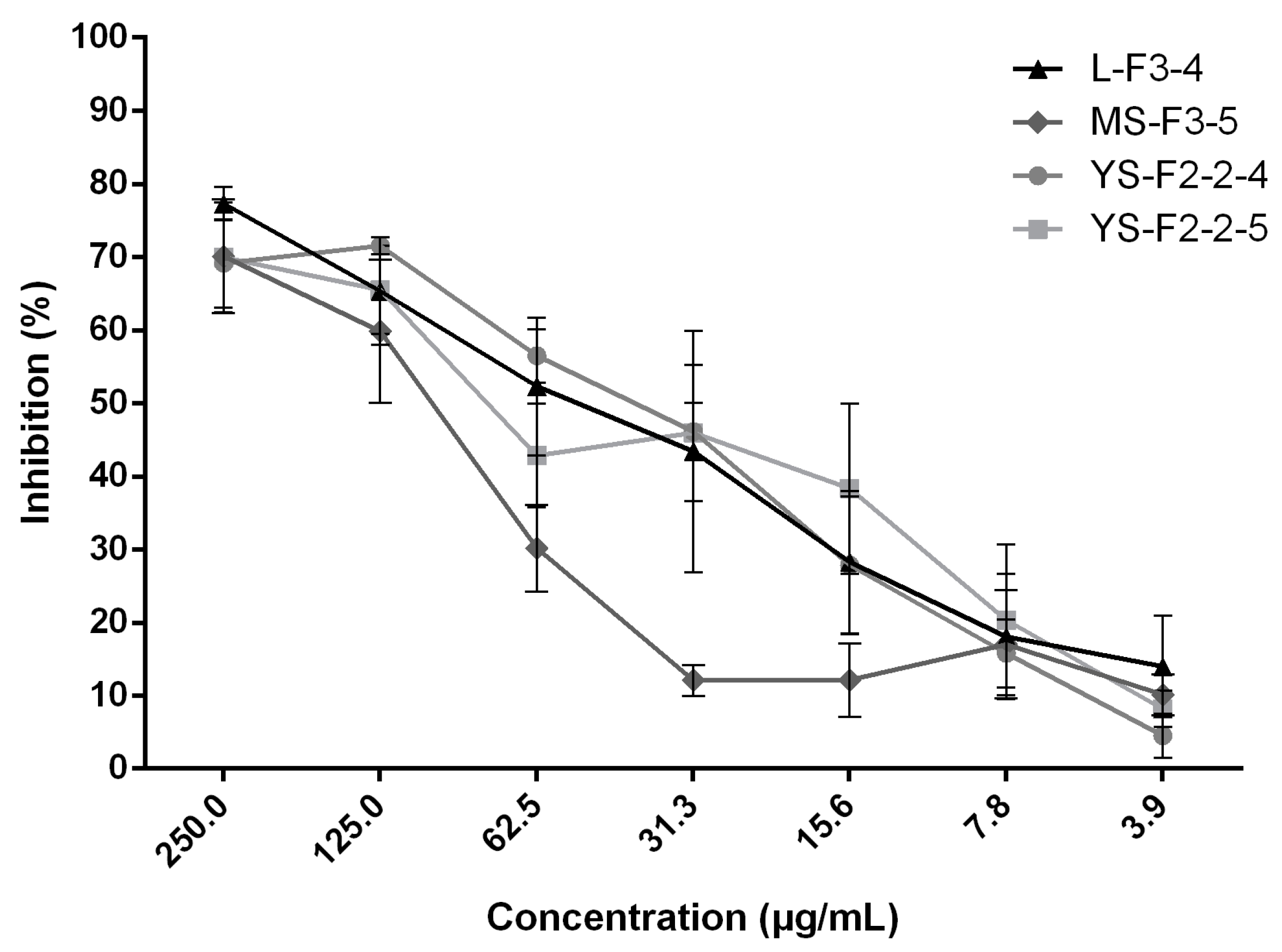
| L-F3-4 | 2 | 62.5 | |
| L-F3-5; L-F3-6 | 0.5–4 | ≥250 | |
| 3-month-old stems | MS-F1; MS-F2 | 25–40 | ≥1000 |
| MS-F3 | 40 | 250 | |
| MS-F4 to MS-F6 | 3–20 | ≥1000 | |
| MS-F3-1 to MS-F3-4 | 0.8–3.5 | ≥250 | |
| MS-F3-5 | 2 | 125 | |
| MS-F3-6; MS-F3-7 | 2–5.3 | ≥250 | |
| 1-year-old stems | YS-F1 | 227 | >400 |
| YS-F2 | 295 | 100 | |
| YS-F3 to YS-F9 | 10–247 | ≥400 | |
| YS-F2-1 | 50 | 200 | |
| YS-F2-2 | 186 | 100 | |
| YS-F2-3; YS-F2-4 | 15–38 | > 400 | |
| YS-F2-2-1 to YS-F2-2-3 | 6–28 | ≥250 | |
| YS-F2-2-4 | 16 | 62.5 | |
| YS-F2-2-5 | 10 | 125 | |
| YS-F2-2-6; YS-F2-2-7 | 5–17 | ≥250 |
| Fraction | Tentative Identification | RT (min) | Formula | Mw | MS Data (m/z) | MS/MS Data (m/z) | Reference |
|---|---|---|---|---|---|---|---|
| L-F3-4 | 12,13-epoxy-9Z-octadecenoic acid | 21.44 | C18H32O3 | 296.23 | 295.18 [M − H]− | 277.29; 259.27; 233.28; 195.18; 183.14; 171.13; 113.11 | UT000014 (NORMAN MassBank) CID 5,356,421 (PubChem Database) |
| trihydroxy-octadecenoic acid | 20.88 | C18H34O5 | 330.24 | 329.19 [M − H]− | 293.30; 211.18; 171.14 | [20] | |
| Ursolic acid based triterpenoid | 22.67 | 517.26 | 455.46; 375.11 | [21] | |||
| p-galloyl-p-coumaroyl-p-cinnamoyl glucose | 30.83 | C31H28O13 | 608.15 | 607.39 [M − H]− | 571.64; 293.30 | [22] | |
| MS-F3-5 | 9-Oxo-10E,12Z-octadecadienoic acid | 21.49 | C18H30O3 | 294.21 | 249.02 [M− CO2 − H]¯ | 185.04; 125.12 | [23] |
| 13S-hydroperoxy-9Z,11E-octadecadienoic acid | 24.10 | C18H32O4 | 312.23 | 311.29 [M − H]− | 293.30; 223.23; 181.16; 171.14; 155.14 | UT000068 (NORMAN MassBank) | |
| Unidentified | 6.27 | 345.27 | 309.30; 291.28; 281.06; 238.22; 209.17; 197.16; 171.14 | ||||
| kaempferol-3-O-malonyl glucoside | 10.27 | C24H22O14 | 534.42 | 533.49 [M − H]− | 487.50, 447.20, 285.10 | [24] | |
| 13S-hydroperoxy-9Z,11E-octadecadienoic acid dimer | 24.14 | (C18H32O4)2 | 312.23 | 623.61 [2M − H]− | 511.51; 329.33; 311.31; 293.27; 249.03 | UT000068 (NORMAN MassBank) | |
| YS-F2-2-4 | 12,13-epoxy-9Z-octadecenoic acid | 21.54 | C18H32O3 | 296.23 | 295.26 [M − H]− | 277.29; 259.27; 233.28; 195.18; 183.14; 171.13; 113.11 | UT000014 (NORMAN MassBank) CID 5,356,421 (PubChem Database) |
| trihydroxy-octadecenoic acid | 22.10 | C18H34O5 | 330.24 | 329.28 [M − H]− | 293.30; 211.18; 171.14 | [20] | |
| Anacardic acid | 22.11 | C22H30O3 | 342.21 | 341.28 [M − H]− | 323.28 295.30; 277.29 | [23] | |
| Daidzein-8-C-glucoside | 6.30 | C21H20O9 | 416.11 | 415.33 [M − H]− | 295.31 | [20] | |
| 12,13-epoxy-9Z-octadecenoic acid, dimer | 21.56 | (C18H32O3)2 | 296.23 | 591.56 [2M − H]− | 545.48; 329.33; 277.29; 195.18; 171.14 | UT000014 (NORMAN MassBank) CID 5,356,421 (PubChem Database) | |
| YS-F2-2-5 | 9-Oxo-10E,12Z-octadecadienoic acid | 21.49 | C18H30O3 | 294.21 | 293.25 [M − H]− | 197.18; 149.12; 125.11 | [23] |
| 15S-hydroperoxy-11Z,13E-eicosadienoic acid | 23.56 | C20H36O4 | 340.50 | 339.27 [M − H]− | 321.27; 307.27 | DFA8147 Lipidbank (JCBL) | |
| 9-Oxo-10E,12Z-octadecadienoic acid, dimer | 21.50 | (C18H30O3)2 | 294.21 | 587.53 [2M − H]− | 293.29; 265.21; 249.02 | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernard, C.; Juin, C.; Vitry, M.; Le, V.T.D.; Verdon, J.; Toullec, A.-S.; Imbert, C.; Girardot, M. Can Leaves and Stems of Rubus idaeus L. Handle Candida albicans Biofilms? Pharmaceuticals 2020, 13, 477. https://doi.org/10.3390/ph13120477
Bernard C, Juin C, Vitry M, Le VTD, Verdon J, Toullec A-S, Imbert C, Girardot M. Can Leaves and Stems of Rubus idaeus L. Handle Candida albicans Biofilms? Pharmaceuticals. 2020; 13(12):477. https://doi.org/10.3390/ph13120477
Chicago/Turabian StyleBernard, Clément, Camille Juin, Marine Vitry, Van Thanh Danh Le, Julien Verdon, Anne-Solène Toullec, Christine Imbert, and Marion Girardot. 2020. "Can Leaves and Stems of Rubus idaeus L. Handle Candida albicans Biofilms?" Pharmaceuticals 13, no. 12: 477. https://doi.org/10.3390/ph13120477
APA StyleBernard, C., Juin, C., Vitry, M., Le, V. T. D., Verdon, J., Toullec, A.-S., Imbert, C., & Girardot, M. (2020). Can Leaves and Stems of Rubus idaeus L. Handle Candida albicans Biofilms? Pharmaceuticals, 13(12), 477. https://doi.org/10.3390/ph13120477