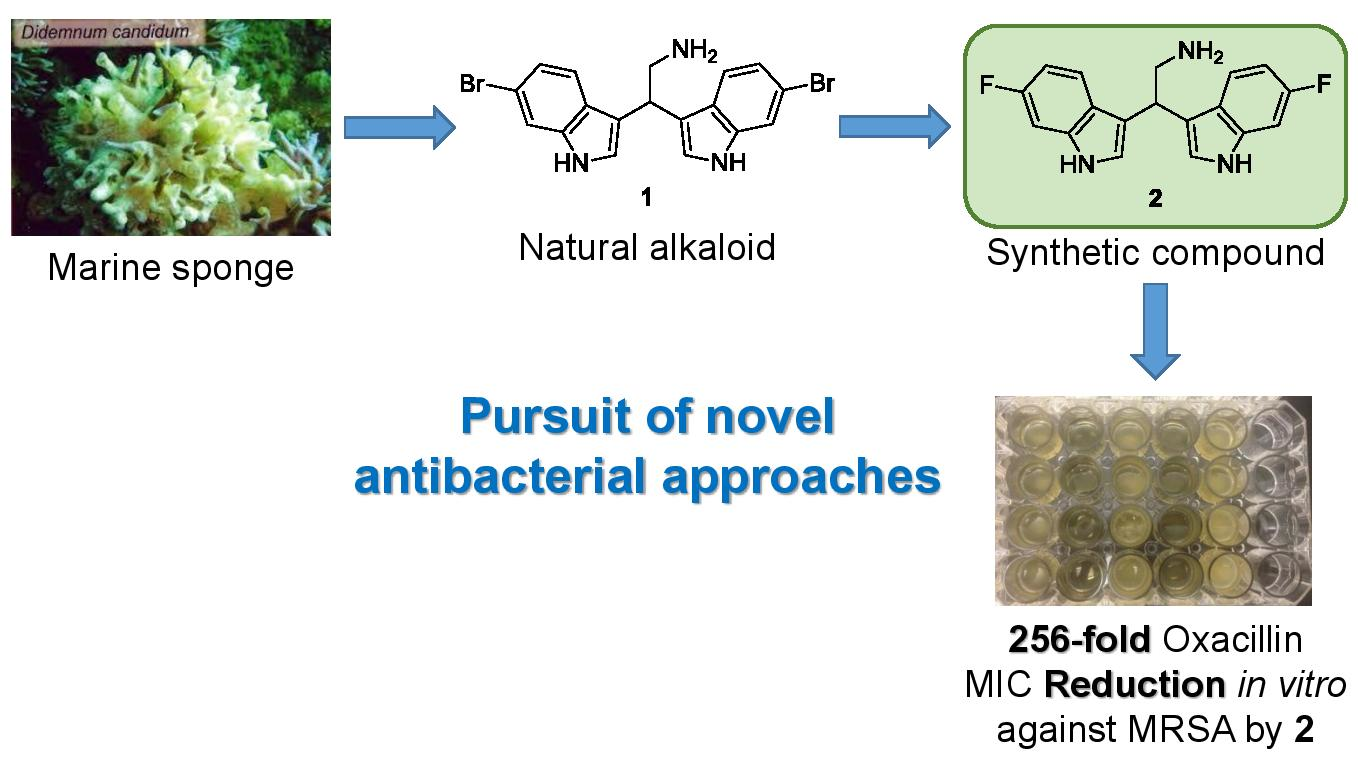
A Fluorinated Analogue of Marine Bisindole Alkaloid 2,2-Bis(6-bromo-1H-indol-3-yl)ethanamine as Potential Anti-Biofilm Agent and Antibiotic Adjuvant Against Staphylococcus aureus

 ,
,  , ,
, ,  and
and
Abstract

1. Introduction
2. Results
2.1. Chemistry
2.2. Antibacterial and Adjuvants Activities of Bisindoles 1 and 2
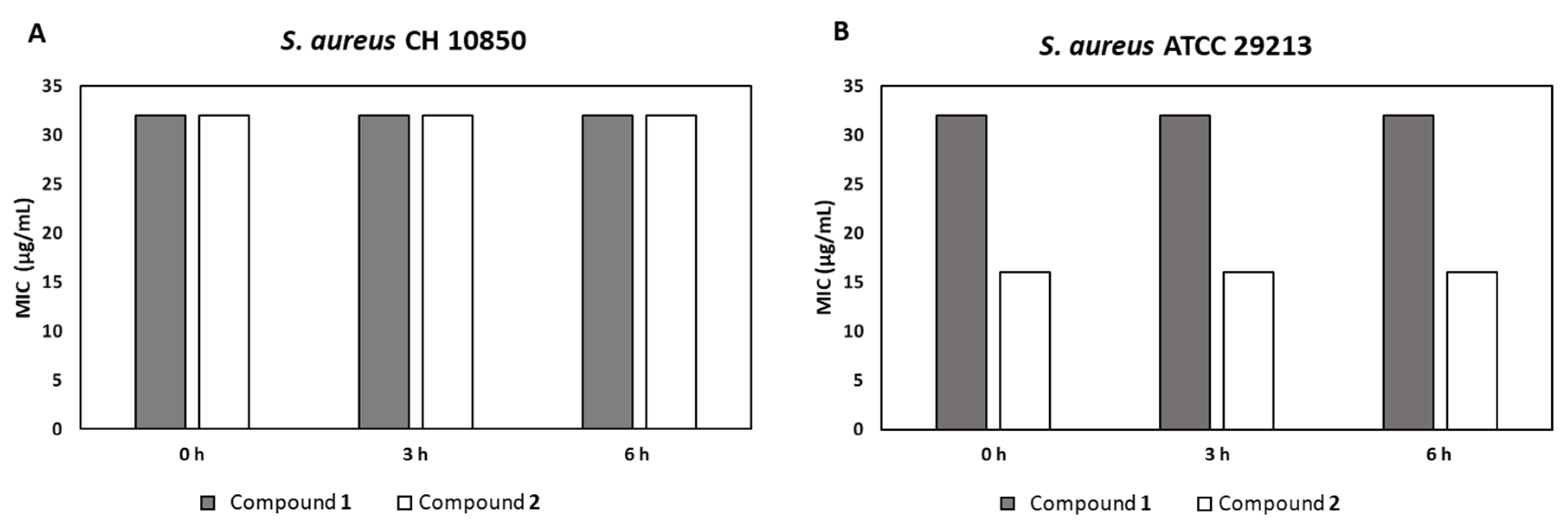
2.3. Antibacterial Activity in Plasma
2.4. Antibiofilm Activity
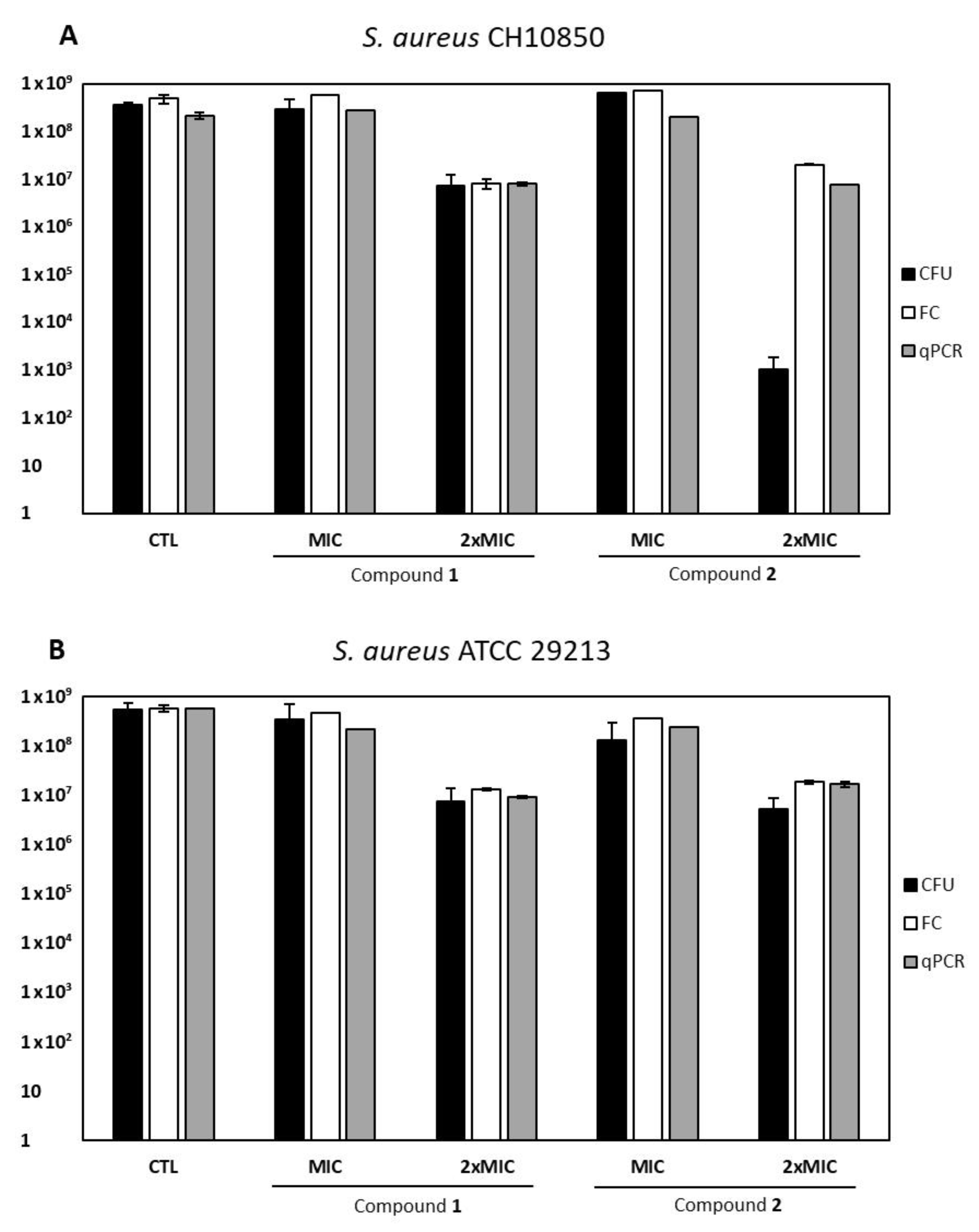
2.5. VBNC Forms Induction
3. Discussion
4. Materials and Methods
4.1. Chemistry
4.1.1. General Procedure for the Synthesis of Derivatives 1–2
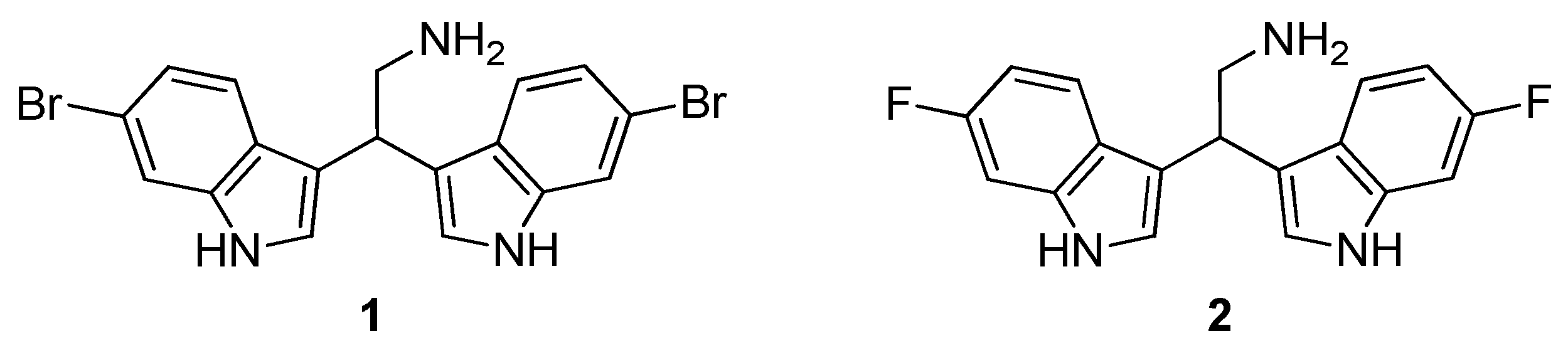
4.1.2. 2,2-Bis(6-bromo-1H-indol-3-yl)ethanamine (1)
4.1.3. 2,2-Bis(6-fluoro-1H-indol-3-yl)ethanamine (2)
4.2. Bacterial Strains
4.3. Determination of Minimum Inhibitory Concentration (MIC)
4.4. Checkerboard Assays
4.5. Hemolytic Activity
4.6. Plasma Stability Assay
4.7. Biofilm Formation Inhibition
4.8. Biofilm-Disrupting Activity
4.9. VBNC Detection
4.9.1. Culture-Based Detection of Staphylococci
4.9.2. Flow Cytometry Detection of Staphylococci
4.9.3. qPCR Detection of Staphylococci
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Calfee, D.P. Trends in community versus health care-acquired methicillin-resistant Staphylococcus aureus infections. Curr. Infect. Dis. Rep. 2017, 19, 48. [Google Scholar] [CrossRef]
- Tverdek, F.P.; Crank, C.W.; Segreti, J. Antibiotic therapy of methicillin-resistant Staphylococcus aureus in critical care. Crit. Care Clin. 2008, 24, 249–260. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Chambers, H.F. Reemergence of antibiotic-resistant Staphylococcus aureus in the genomics era. J. Clin. Investig. 2009, 119, 2464–2474. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, H.; Rudkin, J.K.; Black, N.S.; Gallagher, L.; O’Neill, E.; O’Gara, J.P. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front. Cell Infect. Microbiol. 2015, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Bergkessel, M.; Basta, D.W.; Newman, D.K. The physiology of growth arrest: Uniting molecular and environmental microbiology. Nat. Rev. Microbiol. 2016, 14, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef]
- Ayrapetyan, M.; Williams, T.; Oliver, J.D. Relationship between the viable but nonculturable state and antibiotic persister cells. J. Bacteriol. 2018, 200, e00249-18. [Google Scholar] [CrossRef]
- Melander, R.J.; Melander, C. The challenge of overcoming antibiotic resistance: An adjuvant approach? ACS Infect. Dis. 2017, 3, 559−563. [Google Scholar] [CrossRef] [PubMed]
- Melander, R.J.; Minvielle, M.J.; Melander, C. Controlling bacteria behaviour with indole-containing natural products and derivatives. Tetrahedon 2014, 70, 6363–6372. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J. Search for new bioactive marine natural products and application to drug development. Chem. Pharm. Bull. 2016, 64, 1079–1083. [Google Scholar] [CrossRef]
- Mari, M.; Tassoni, A.; Lucarini, S.; Fanelli, M.; Piersanti, G.; Spadoni, G. Brønsted acid catalyzed bisindolization of α-amido acetals: Synthesis and anticancer activity of bis (indolyl) ethanamino derivatives. Eur. J. Org. Chem. 2014, 18, 3822–3830. [Google Scholar] [CrossRef]
- Mantenuto, S.; Lucarini, S.; De Santi, M.; Piersanti, G.; Brandi, G.; Favi, G.; Mantellini, F. One-pot synthesis of biheterocycles based on indole and azole scaffolds using tryptamines and 1, 2-diaza-1, 3-dienes as building blocks. Eur. J. Org. Chem. 2016, 19, 3193–3199. [Google Scholar] [CrossRef]
- Salucci, S.; Burattini, S.; Buontempo, F.; Orsini, E.; Furiassi, L.; Mari, M.; Lucarini, S.; Martelli, A.M.; Falcieri, E. Marine bisindole alkaloid: A potential apoptotic inducer in human cancer cells. Eur. J. Histochem. 2018, 62, 2881. [Google Scholar] [CrossRef] [PubMed]
- Choppara, P.; Bethu, M.S.; Vara Prasad, Y.; Venkateswara Rao, J.; Uday Ranjan, T.J.; Siva Prasad, G.V.; Doradla, R.; Murthy, Y.L.N. Synthesis, characterization and cytotoxic investigations of novel bis(indole) analogues besides antimicrobial study. Arab. J. Chem. 2015, 12, 2721–2731. [Google Scholar] [CrossRef]
- Campana, R.; Favi, G.; Baffone, W.; Lucarini, S. Marine alkaloid 2,2-bis(6-bromo-3-indolyl) ethylamine and its synthetic derivatives inhibit microbial biofilms formation and disaggregate developed biofilms. Microrganisms 2019, 7, 28. [Google Scholar] [CrossRef]
- Hubble, V.B.; Hubbard, B.A.; Minrovic, B.M.; Melander, R.J.; Melander, C. Using small-molecule adjuvants to repurpose azithromycin for use against Pseudomonas aeruginosa. ACS Infect. Dis. 2019, 5, 141–151. [Google Scholar] [CrossRef]
- Isanbor, C.; O’Hagan, D. Fluorine in medicinal chemistry: A review of anti-cancer agents. J. Fluor. Chem. 2006, 127, 303–319. [Google Scholar] [CrossRef]
- Zhang, Q.; Teschers, C.S.; Callejo, R.; Yang, M.; Wang, M.; Silk, P.J.; Ryall, K.; Roscoe, L.E.; Cordes, D.B.; Slawin, A.M.Z.; et al. Fluorine in pheromones: Synthesis of fluorinated 12-dodecanolides as emerald ash borer pheromone mimetics. Tetrahedron 2019, 75, 2917–2922. [Google Scholar] [CrossRef]
- Gillam, E.M.J.; Notley, L.M.; Cai, H.; De Voss, J.J.; Guengerich, F.P. Oxidation of indole by cytochrome P450 enzymes. Biochemistry 2000, 45, 13817–13824. [Google Scholar] [CrossRef] [PubMed]
- Mor, M.; Silva, C.; Vacondio, F.; Plazzi, P.V.; Bertoni, S.; Spadoni, G.; Diamantini, G.; Bedini, A.; Tarzia, G.; Zusso, M.; et al. Indole-based analogs of melatonin: In Vitro antioxidant and cytoprotective activities. J. Pineal Res. 2004, 36, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Ciulla, M.G.; Kumar, K. The natural and synthetic indole weaponry against bacteria. Tetrahedron Lett. 2018, 59, 3223–3233. [Google Scholar] [CrossRef]
- Campana, R.; Sisti, M.; Sabatini, L.; Lucarini, S. Marine bisindole alkaloid 2,2-bis(6-bromo-3-indolyl) ethylamine to control and prevent fungal growth on building material: A potential antifungal agent. Appl. Microbiol. Biotechnol. 2019, 103, 5607–5616. [Google Scholar] [CrossRef]
- Zoraghi, R.; Worrall, L.; See, R.H.; Strangman, W.; Popplewell, W.L.; Gong, H.; Samaai, T.; Swayze, R.D.; Kaur, S.; Vuckovic, M.; et al. Methicillin-resistant Staphylococcus aureus (MRSA) pyruvate kinase as a target for bis-indole alkaloids with antibacterial activities. J. Biol. Chem. 2011, 286, 44716–44725. [Google Scholar] [CrossRef]
- Veale, C.G.L.; Zoraghi, R.; Young, R.M.; Morrison, J.; Pretheeban, M.; Lobb, K.A.; Reiner, N.E.; Andrersen, R.J.; Davies-Coleman, M.T. Synthetic analogues of the marine bisindole deoxytopsentin: Potent selective inhibitors of MRSA pyruvate kinase. J. Nat. Prod. 2015, 78, 355−362. [Google Scholar] [CrossRef]
- Worthington, R.J.; Richards, J.J.; Melander, C. Small molecule control of bacterial biofilms. Org. Biomol. Chem. 2012, 10, 7457–7474. [Google Scholar] [CrossRef]
- Lee, J.H.; Cho, H.S.; Kim, Y.; Kim, J.A.; Banskota, S.; Cho, M.H.; Lee, J. Indole and 7-benzyloxyindole attenuate the virulence of Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2013, 97, 4543–4552. [Google Scholar] [CrossRef]
- Organic Chemistry Portal. Available online: https://www.organic-chemistry.org/prog/ (accessed on 22 November 2019).
- Pasquaroli, S.; Zandri, G.; Vignaroli, C.; Vuotto, C.; Donelli, G.; Biavasco, F. Antibiotic pressure can induce the viable but non-culturable state in Staphylococcus aureus growing in biofilms. J. Antimicrob. Chemother. 2013, 68, 1812–1817. [Google Scholar] [CrossRef]
- Pasquaroli, S.; Citterio, B.; Mangiaterra, G.; Biavasco, F.; Vignaroli, C. Influence of sublethal concentrations of vancomycin and quinupristin/dalfopristin on the persistence of viable but non-culturable Staphylococcus aureus growing in biofilms. J. Antimicrob. Chemother. 2018, 73, 3526–3529. [Google Scholar] [CrossRef]
- Donelli, G.; Francolini, I.; Romoli, D.; Guaglianone, E.; Piozzi, A.; Ragunath, C.; Kaplan, J.B. Synergistic activity of Dispersin B and Cefamandole Nafate in inhibition of Staphylococcal biofilm growth on polyurethanes. Antimicrob. Agents Chemother. 2007, 51, 2733–2740. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). M07-A10: Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 10th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; p. 66. [Google Scholar]
- El-Azizi, M. Novel microdilution method to assess double and triple antibiotic combination therapy in vitro. Int. J. Microbiol. 2016, 4612021. [Google Scholar]
- Ghosh, C.; Manjunath, G.B.; Akkapeddi, P.; Yarlagadda, V.; Hoque, J.; Uppu, D.S.; Konai, M.M.; Haldar, J. Small molecular antibacterial peptoid mimics: The simpler the better! J. Med. Chem. 2014, 57, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, A.; Luna, G.M.; Vignaroli, C.; Pasquaroli, S.; Tota, S.; Paroncini, P.; Biavasco, F. Aquaculture can promote the presence and spread of antibiotic-resistant Enterococci in marine sediments. PLoS ONE 2013, 8, e62838. [Google Scholar] [CrossRef]
- Mangiaterra, G.; Amiri, M.; Di Cesare, A.; Pasquaroli, S.; Manso, E.; Cirilli, N.; Citterio, B.; Vignaroli, C.; Biavasco, F. Detection of viable but non-culturable Pseudomonas aeruginosa in cystic fibrosis by qPCR: A validation study. BMC Infect. Dis. 2018, 18, 701. [Google Scholar] [CrossRef]
- Hein, I.; Lehner, A.; Rieck, P.; Klein, K.; Brandl, E.; Wagner, M. Comparison of different approaches to quantify Staphylococcus aureus cells by real-time quantitative PCR and application of this technique for examination of cheese. Appl. Environ. Microbiol. 2001, 67, 3122–3126. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
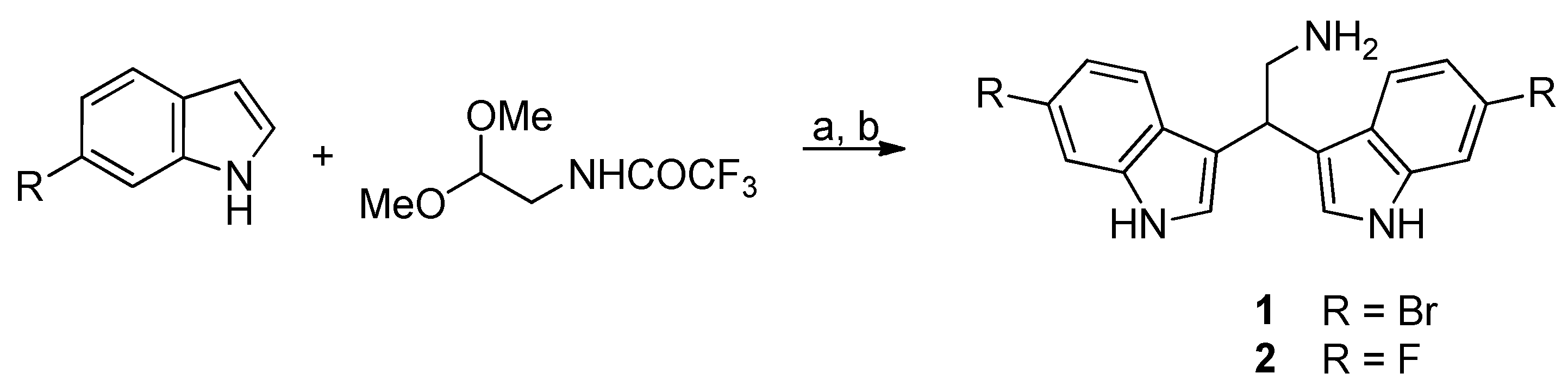{kind=link}
| Compound | MIC (µg/mL) | Hemolysis (%) | |
|---|---|---|---|
| S. aureus CH 10850 (MRSA) | S. aureus ATCC 29213 | ||
| 1 | 2 | 2 | 2.96 ± 0.02 |
| 2 | 32 | 16 | 2.32 ± 0.06 |
| Compound | Concentration (µg/mL) | Oxacillin MIC (µg/mL) | ΣFIC | |
|---|---|---|---|---|
| None | - | 256 | ||
| 1 | 1 | 128 | 1.0 | Additive |
| 2 | 16 | 1 | 0.5 | Synergistic |
| S. aureus Strain | Biofilm Formation Inhibition | Disaggregating Ability | ||||
|---|---|---|---|---|---|---|
| Compound 1 | Compound 2 | Compound 1 | Compound 2 | |||
| MIC | 2× MIC | MIC | 2× MIC | MIC | MIC | |
| CH 10850 (MRSA) | 52.6% | 100% | 61.0% | 76.5% | 37.5% | 56.3% |
| ATCC 29213 | 49.6% | 100% | 58.9% | 100% | 28.0% | 53.9% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campana, R.; Mangiaterra, G.; Tiboni, M.; Frangipani, E.; Biavasco, F.; Lucarini, S.; Citterio, B. A Fluorinated Analogue of Marine Bisindole Alkaloid 2,2-Bis(6-bromo-1H-indol-3-yl)ethanamine as Potential Anti-Biofilm Agent and Antibiotic Adjuvant Against Staphylococcus aureus. Pharmaceuticals 2020, 13, 210. https://doi.org/10.3390/ph13090210
Campana R, Mangiaterra G, Tiboni M, Frangipani E, Biavasco F, Lucarini S, Citterio B. A Fluorinated Analogue of Marine Bisindole Alkaloid 2,2-Bis(6-bromo-1H-indol-3-yl)ethanamine as Potential Anti-Biofilm Agent and Antibiotic Adjuvant Against Staphylococcus aureus. Pharmaceuticals. 2020; 13(9):210. https://doi.org/10.3390/ph13090210
Chicago/Turabian StyleCampana, Raffaella, Gianmarco Mangiaterra, Mattia Tiboni, Emanuela Frangipani, Francesca Biavasco, Simone Lucarini, and Barbara Citterio. 2020. "A Fluorinated Analogue of Marine Bisindole Alkaloid 2,2-Bis(6-bromo-1H-indol-3-yl)ethanamine as Potential Anti-Biofilm Agent and Antibiotic Adjuvant Against Staphylococcus aureus" Pharmaceuticals 13, no. 9: 210. https://doi.org/10.3390/ph13090210
APA StyleCampana, R., Mangiaterra, G., Tiboni, M., Frangipani, E., Biavasco, F., Lucarini, S., & Citterio, B. (2020). A Fluorinated Analogue of Marine Bisindole Alkaloid 2,2-Bis(6-bromo-1H-indol-3-yl)ethanamine as Potential Anti-Biofilm Agent and Antibiotic Adjuvant Against Staphylococcus aureus. Pharmaceuticals, 13(9), 210. https://doi.org/10.3390/ph13090210