
Anti-Cancer Potential of Afzelin towards AGS Gastric Cancer Cells

 , , , and
, , , and
Abstract
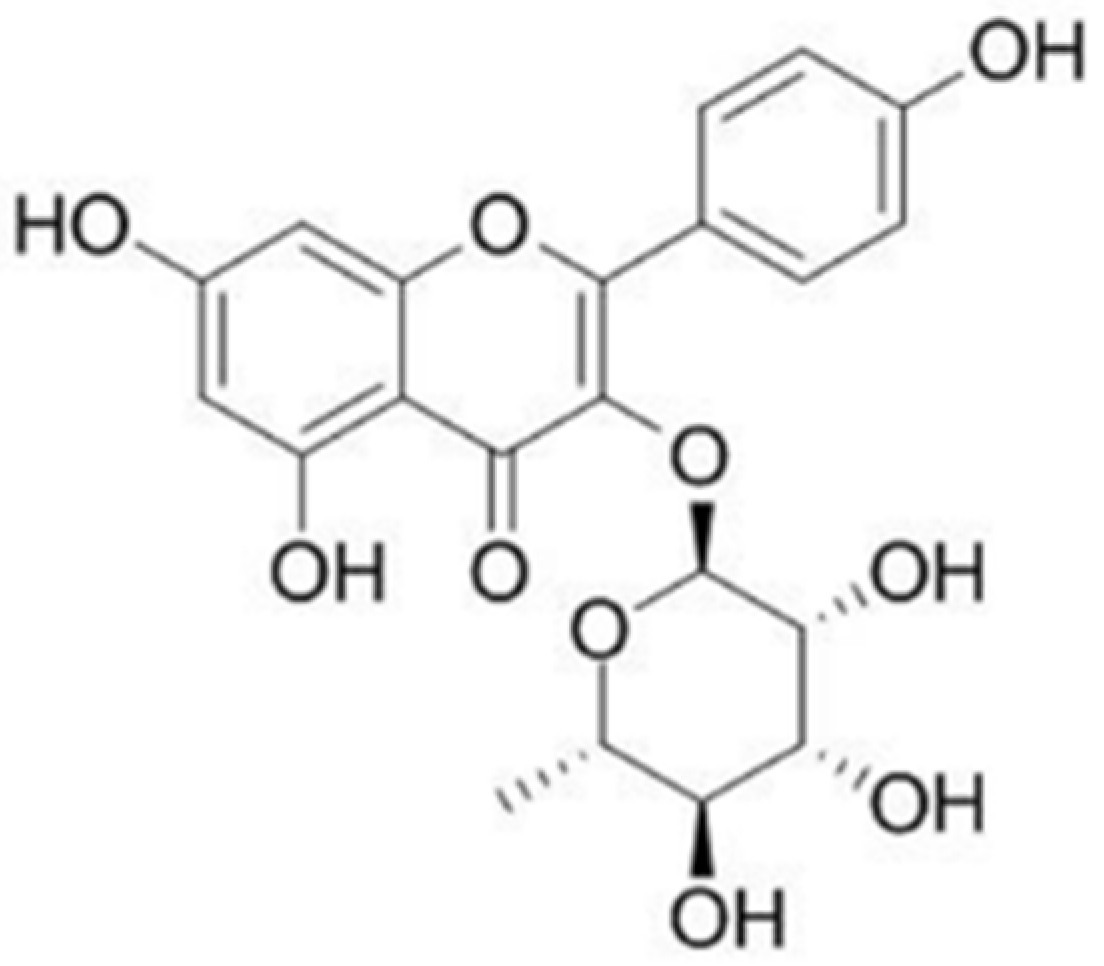
:1. Introduction
2. Results
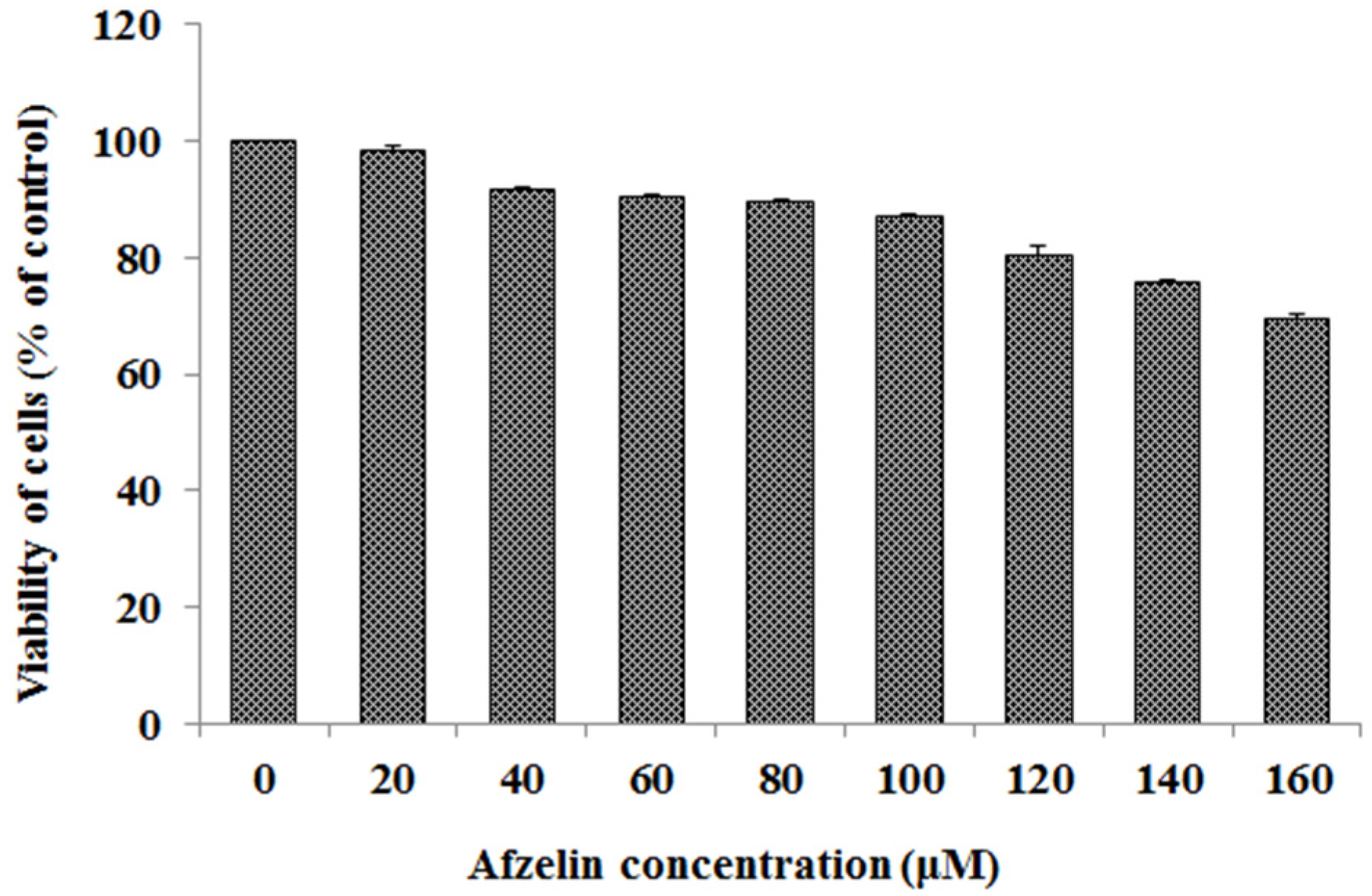
2.1. Viability of AGS Cancer Cells
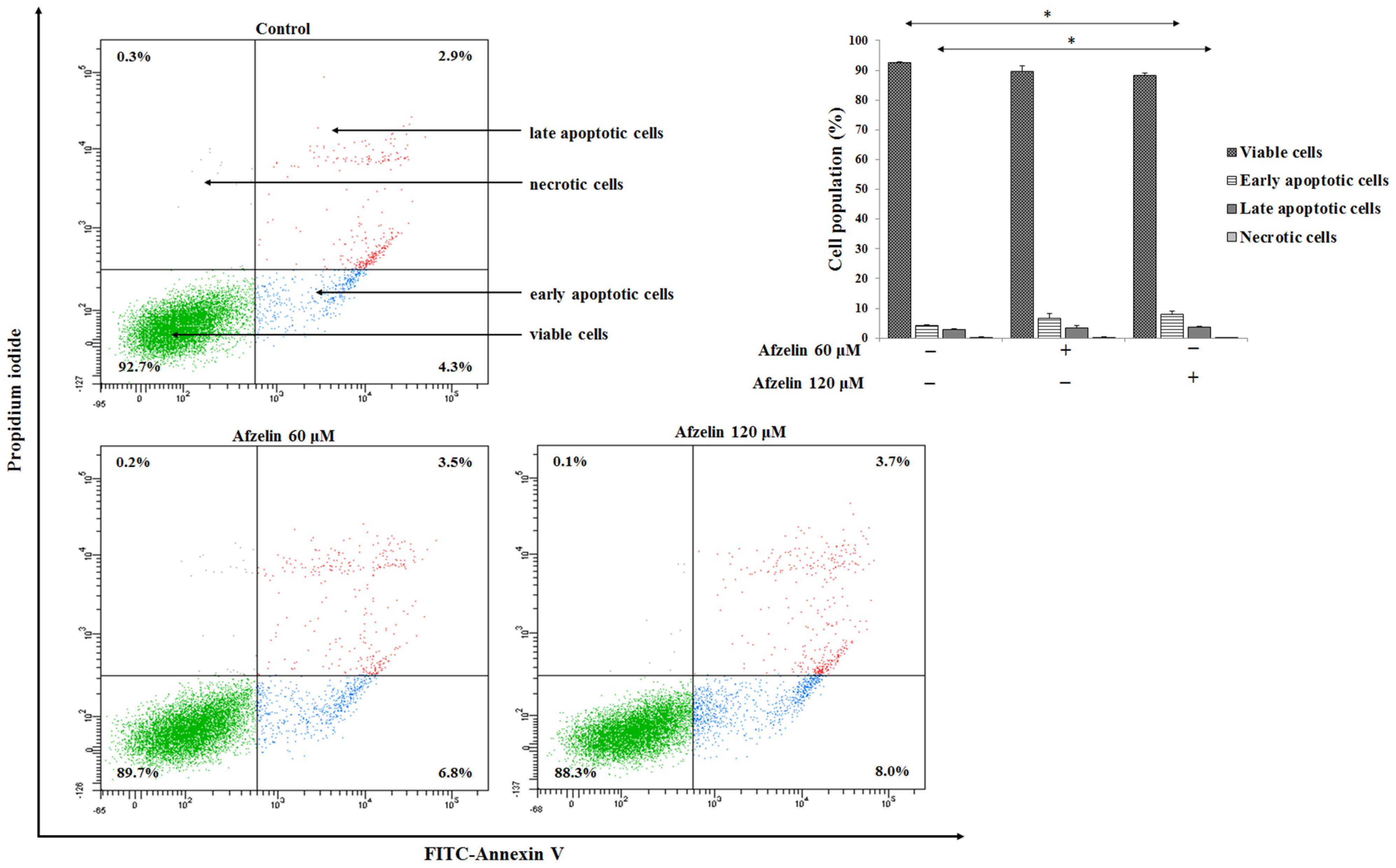
2.2. Impact of Afzelin on Apoptosis
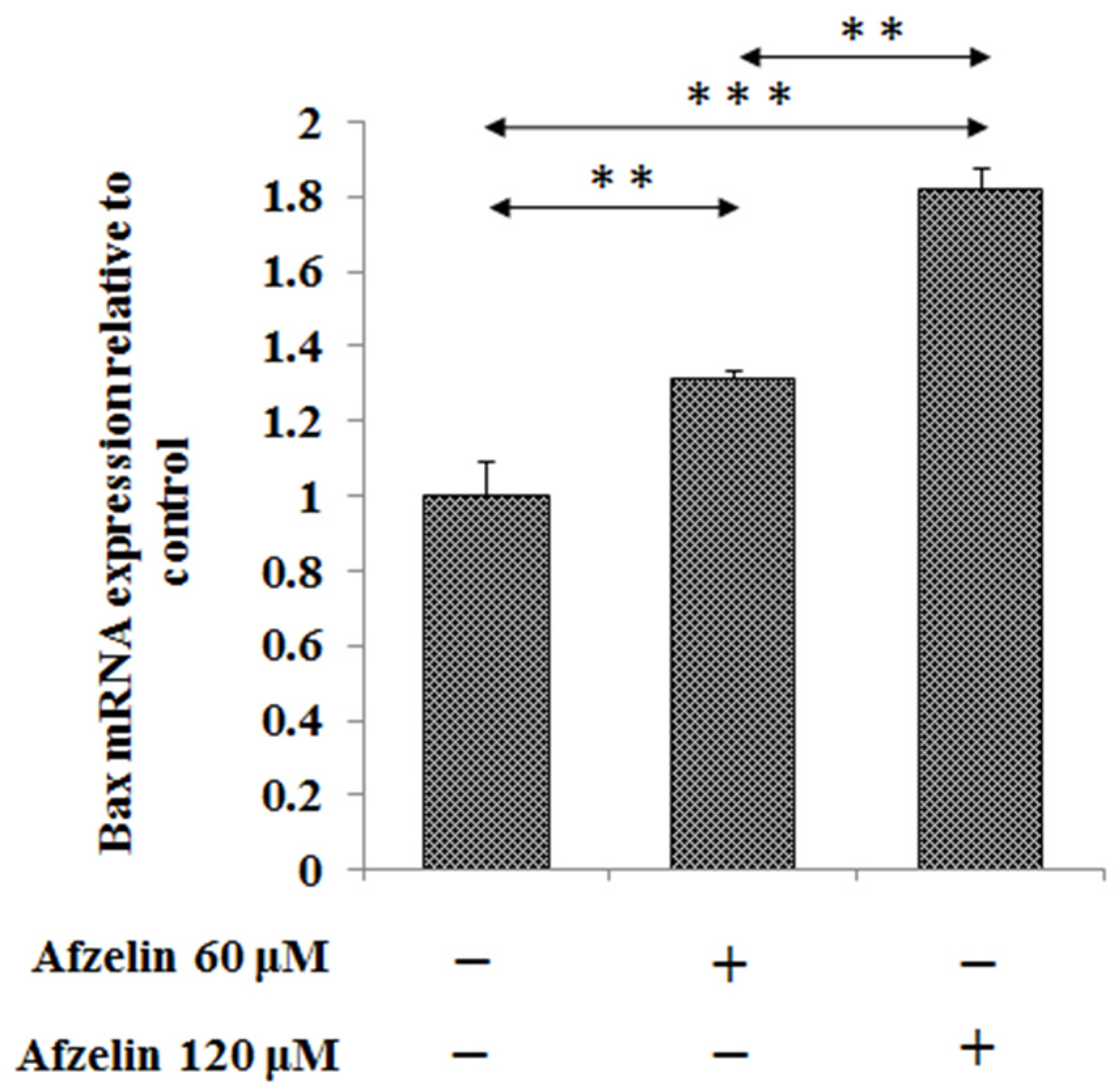
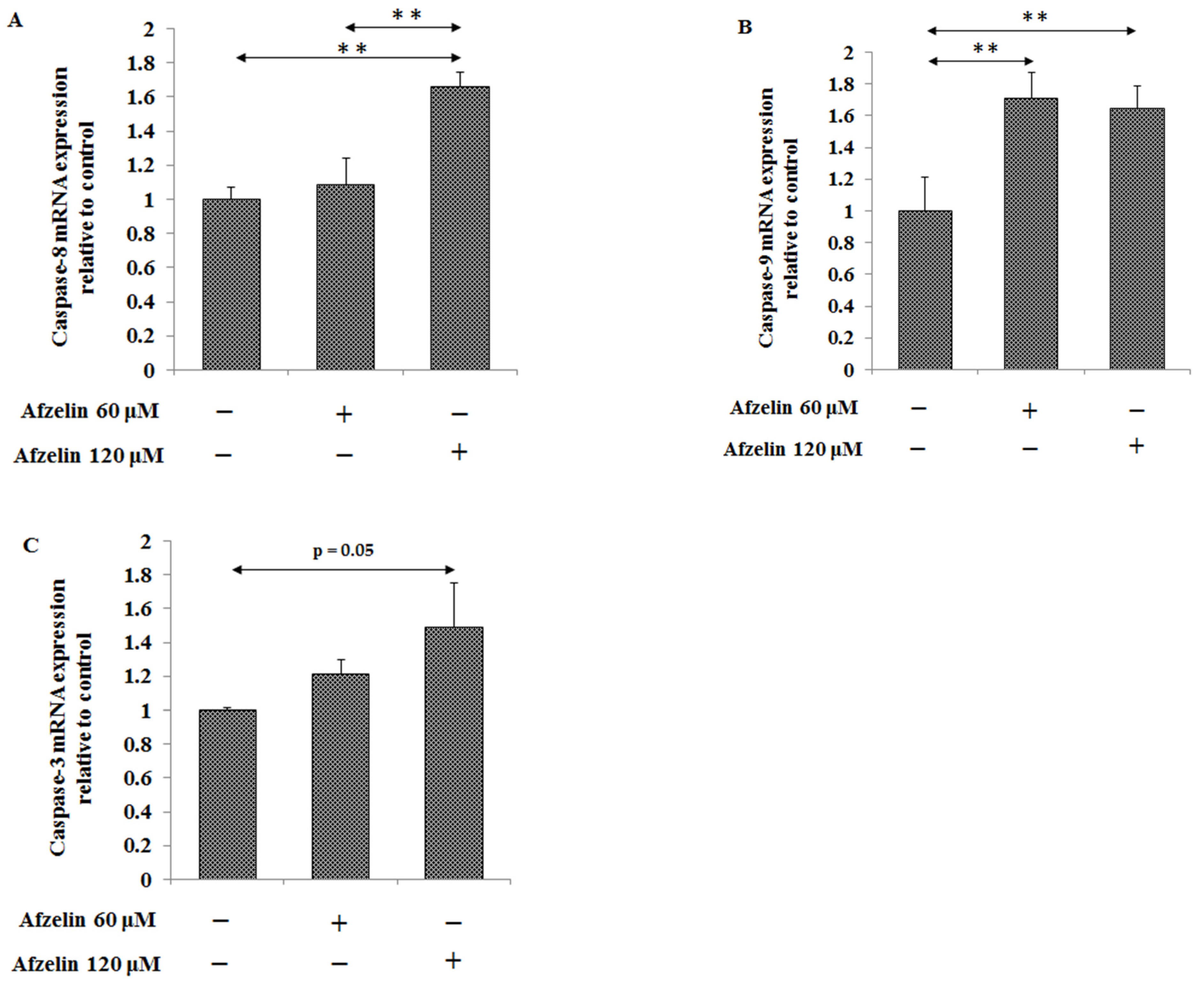
2.3. The Effect of Afzelin on Bax and Caspase-3, -8, -9
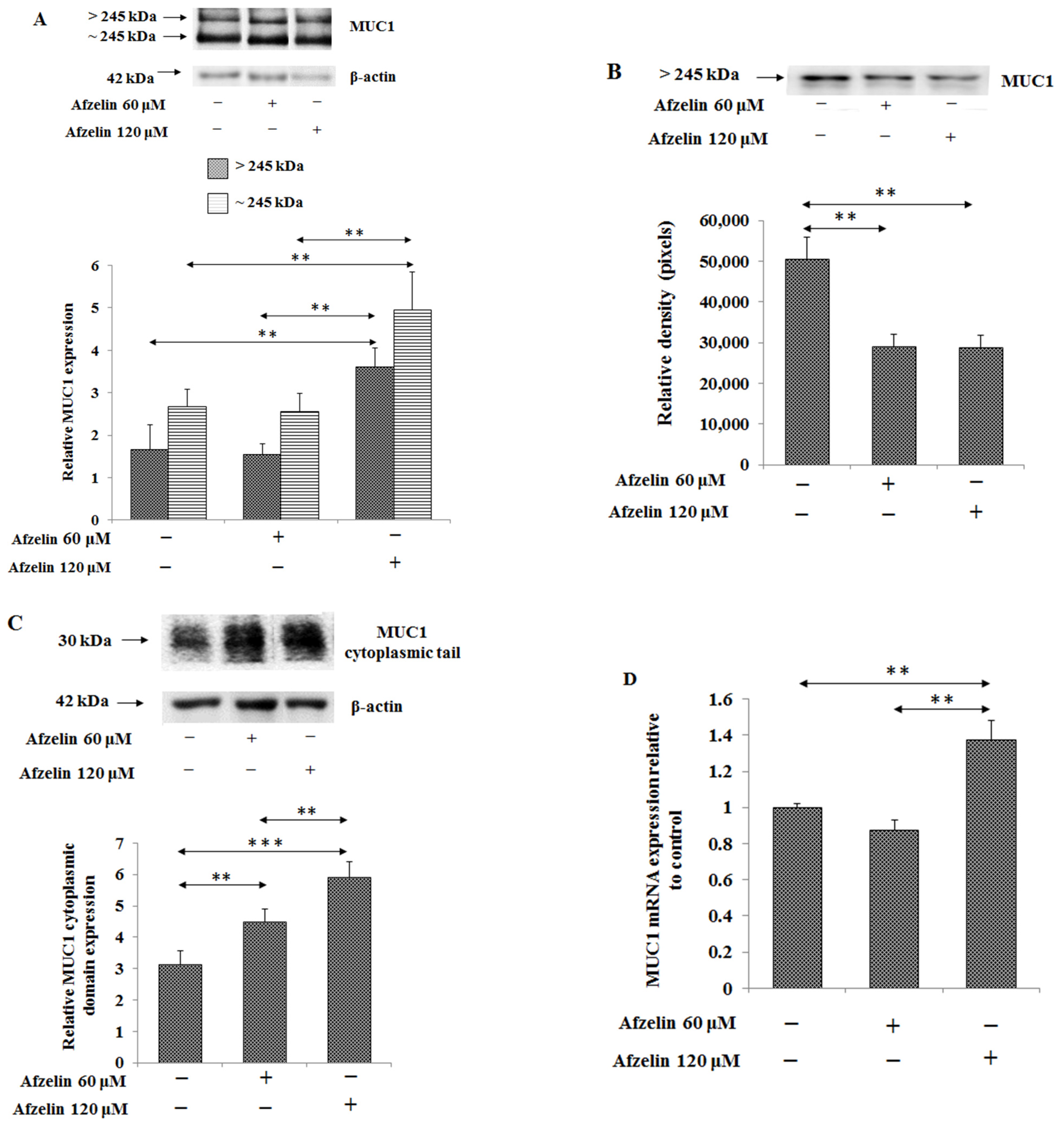
2.4. The Effect of Afzelin on MUC1
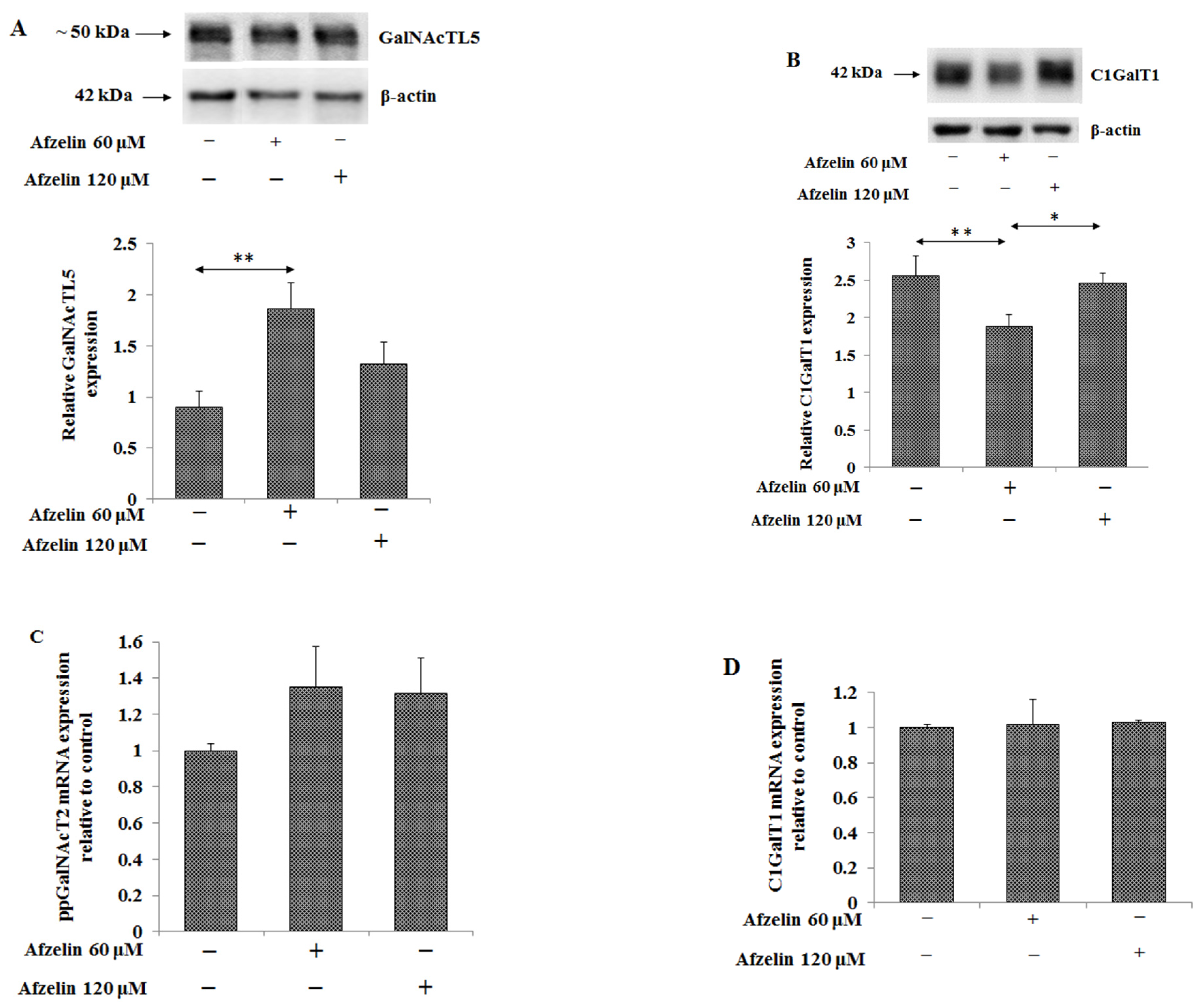
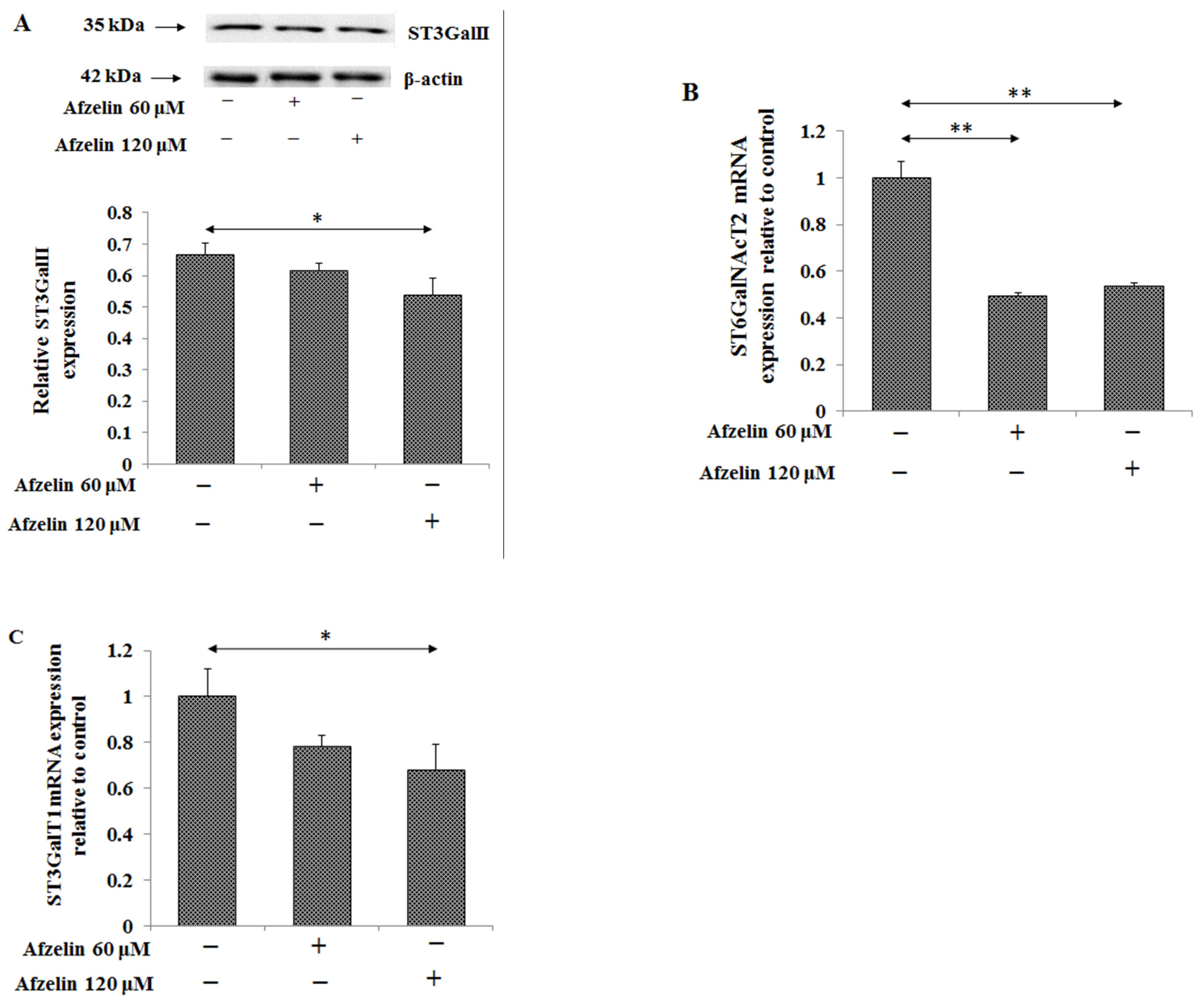
2.5. The Effect of Afzelin on Tn, T, sTn, and sT Antigens
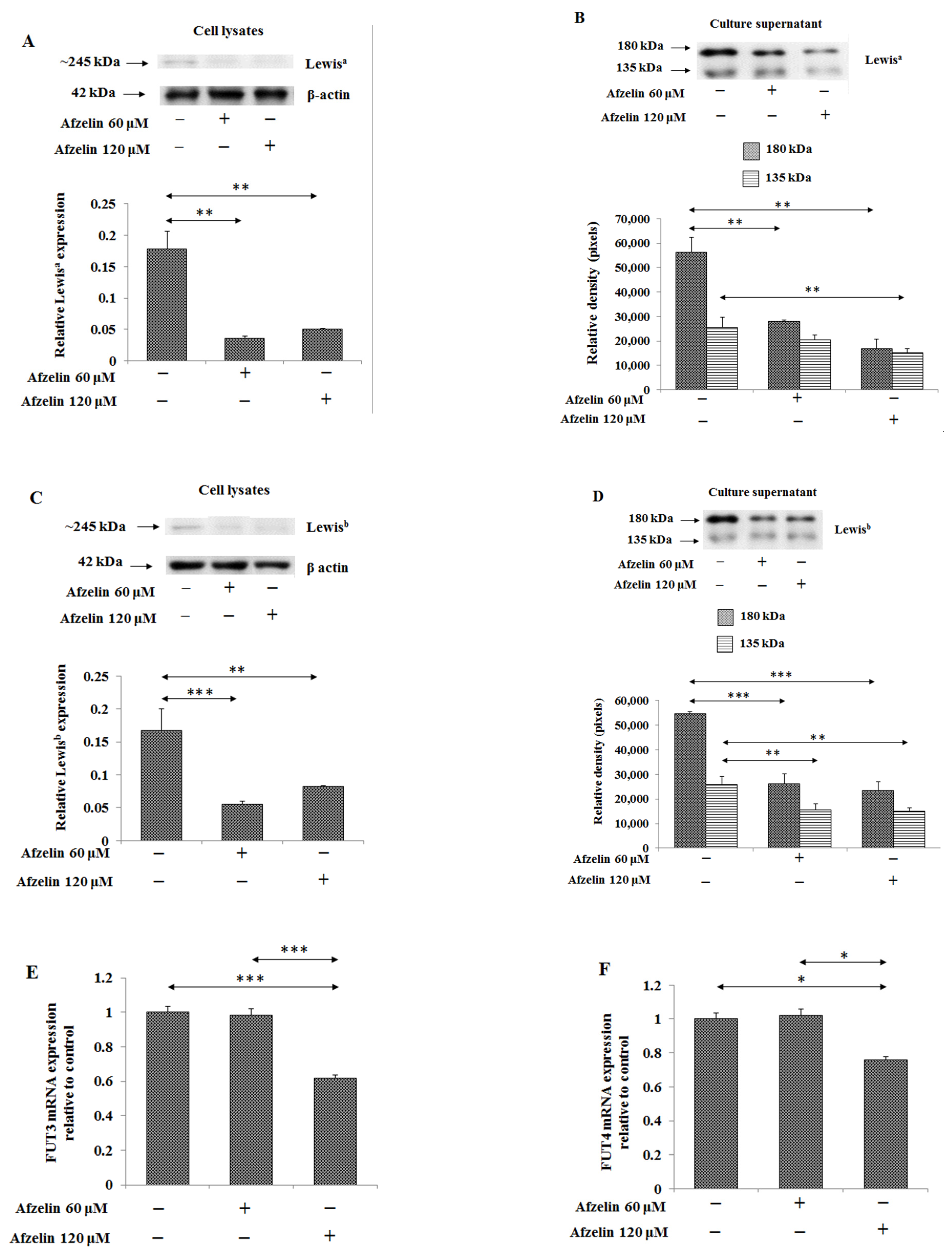
2.6. The Effect of Afzelin on Lewis Antigens
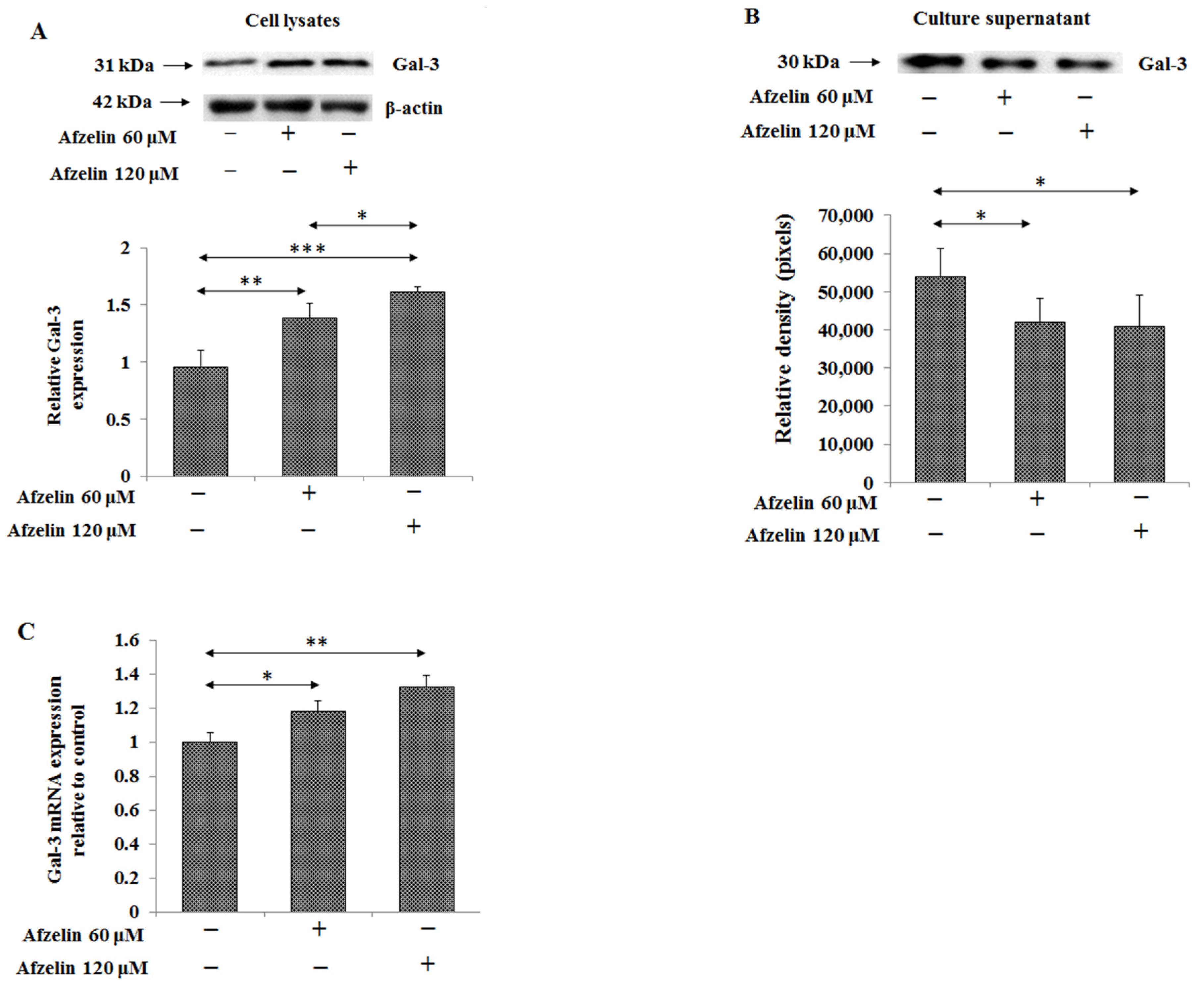
2.7. The Effect of Afzelin on Gal-3
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability Test
4.3. Flow Cytometry Assessment of Annexin V Binding
4.4. Western Blotting
4.5. Real-Time PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Song, Z.; Wu, Y.; Yang, J.; Yang, D.; Fang, X. Progress in the treatment of advanced gastric cancer. Tumor Biol. 2017, 39, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diantini, A.; Subarnas, A.; Lestari, K.; Halimah, E.; Susilawati, Y.; Supriyatna, S.; Julaeha, E.; Achmad, T.H.; Suradji, E.W.; Yamazaki, C.; et al. Kaempferol-3-O-rhamnoside isolated from the leaves of Schima wallichii Korth inhibits MCF-7 breast cancer cell proliferation through activation of the caspase cascade pathway. Oncol. Lett. 2012, 3, 1069–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, K.C.; Sun, J.M.; Shen, J.G.; Jin, J.Z.; Liu, F.; Xu, X.L.; Chen, L.; Liu, L.T.; Lv, J.J. Afzelin exhibits anti-cancer activity against androgen-sensitive LNCaP and androgen-independent PC-3 prostate cancer cells through the inhibition of LIM domain kinase 1. Oncol. Lett. 2015, 10, 2359–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Nie, X. Afzelin attenuates asthma phenotypes by downregulation of GATA3 in a murine model of asthma. Mol. Med. Rep. 2015, 12, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Casio, S.; Finn, O.J. Intra- and extra-cellular events related to altered glycosylation of MUC1 promote chronic inflammation, tumor progression, invasion, and metastasis. Biomolecules 2016, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Nath, S.; Mukherjee, P. MUC1: A multifaceted oncoprotein with a key role in cancer progression. Trends Mol. Med. 2014, 20, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.A.; Magalhaes, A.; Gomes, J.; Peixoto, A.; Gaiteiro, C.; Fernandes, E.; Santos, L.L.; Reis, C.A. Protein glycosylation in gastric and colorectal cancers: Toward cancer detection and target therapeutics. Cancer Lett. 2017, 387, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Kudelka, M.R.; Ju, T.; Heimburg-Molinaro, J.; Cummings, R.D. Simple sugars to complex disease–mucin-type O-glycans in cancer. Adv. Cancer Res. 2015, 126, 53–135. [Google Scholar]
- Padro, M.; Mejias-Luque, R.; Cobler, L.; Garrido, M.; Perez-Garay, M.; Puig, S.; Peracaula, R.; de Bolos, C. Regulation of glycosyltransferases and Lewis antigens expression by IL-1β and IL-6 in human gastric cancer cells. Glycoconj. J. 2011, 28, 99–110. [Google Scholar] [CrossRef]
- Dong, R.; Znang, M.; Hu, Q.; Zheng, S.; Soh, A.; Zheng, Y.; Yuan, H. Galectin-3 as a novel biomarker for disease diagnosis and a target for therapy (Review). Int. J. Mol. Med. 2018, 41, 599–614. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Guo, X.; Nash, G.B.; Stone, P.C.; Hilkens, J.; Rhodes, J.M.; Yu, L.G. Circulating galectin-3 promotes metastasis by modifying MUC1 localization on cancer cell surface. Cancer Res. 2009, 69, 6799–6806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Liu, Y.; Wu, L.; Sun, X.L. Sialyltransferase inhibition and recent advances. Biochim. Biophys. Acta 2016, 1864, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Hidari, K.I.P.J.; Oyama, K.; Ito, G.; Nakayama, M.; Inai, M.; Goto, S.; Kanai, Y.; Watanabe, K.; Yoshida, K.; Furuta, T.; et al. Identification and characterization of flavonoids as sialyltransferase inhibitors. Biochem. Biophys. Res. Commun. 2009, 382, 609–613. [Google Scholar] [CrossRef]
- Radziejewska, I.; Supruniuk, K.; Nazaruk, J.; Karna, E.; Popławska, B.; Bielawska, A. Rosmarinic acid influences collagen, MMPs, TIMPs, glycosylation and MUC1 in CRL-1739 gastric cancer cell line. Biomed. Pharmacother. 2018, 107, 397–407. [Google Scholar] [CrossRef]
- Radziejewska, I.; Supruniuk, K.; Bielawska, A. Anti-cancer effect of combined action of anti-MUC1 and rormarinic acid in AGS gastric cancer cells. Eur. J. Pharmacol. 2021, 902, 174119. [Google Scholar] [CrossRef]
- Radziejewska, I.; Borzym-Kluczyk, M.; Leszczyńska, K. Luteolin alters MUC1 extracellular domain, sT antygen, ADAM-17, IL-8, IL-10 and NF-κB expression in Helicobacter pylori infected gastric cancer CRL-1739 cells: A preliminary study. Biomed. Rep. 2021, 14, 19. [Google Scholar] [CrossRef]
- Bisol, A.; de Campos, P.S.; Lamers, M.L. Flavonoids as anticancer therapies: A systematic review of clinical trials. Phytother. Res. 2020, 34, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, H.; Sun, X.; Li, M. Apoptosis-induction is a novel therapeutic strategy for gastrointestinal and liver cancers. Curr. Gene Ther. 2015, 15, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Rahn, J.J.; Chow, J.W.; Horne, G.J.; Mah, B.K.; Emerman, J.T.; Hoffman, P.; Hugh, J.C. MUC1 mediates trans endothelial migration in vitro by ligating endothelial cell ICAM-1. Clin. Exp. Metastasis 2005, 22, 475–483. [Google Scholar] [CrossRef]
- Zhao, Q.; Piyush, T.; Chen, C.; Hollingsworth, M.A.; Hilkens, J.; Rhodes, J.M.; Yu, L.G. MUC1 extracellular domain confers resistance of epithelial cancer cell to anoikis. Cell Death Dis. 2014, 5, e1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascio, S.; Zhang, L.; Finn, O.J. MUC1 protein expression in tumor cells regulates transcription of proinflammatory cytokines by forming a complex with nuclear factor-κB p65 and binding to cytokine promoters. J. Biol. Chem. 2011, 286, 42248–42256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Rajabi, H.; Kufe, D.M. Mucin 1 C-terminal subunit oncoprotein is a target for small-molecule inhibitors. Mol. Pharmacol. 2011, 79, 886–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Yang, L.; Li, S.; Ye, D.; Yang, L.; Liu, Q.; Zhao, Z.; Cai, Q.; Tan, J.; Li, X. Quercetin inhibits breast cancer stem cells via downregulation of aldehyde dehydrogenase 1A1 (ALDH1A1), chemokine receptor type 4 (CXCR4), mucin 1 (MUC1), and epithelial cell adhesion molecule (EpCAM). Med. Sci. Monit. 2018, 24, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Mereiter, S.; Balmana, M.; Gomes, J.; Magalhaes, A.; Reis, C.A. Glycomic approaches for the discovery of targets in gastrointestinal cancer. Front Oncol 6:55Moriwaki K, Miyoshi E (2010) Fucosylation and gastrointestinal cancer. World J. Hepatol. 2016, 2, 151–161. [Google Scholar]
- Hua, D.; Shen, L.; Xu, L.; Jiang, Z.; Zhou, Y.; Yue, A.; Zou, S.; Cheng, Z.; Wu, S. Polypeptide N-acetylgalactosaminyltransferase 2 regulates cellular metastasis-associated behavior in gastric cancer. Int. J. Mol. Med. 2012, 30, 1267–1274. [Google Scholar] [CrossRef] [Green Version]
- Gomes, J.; Marcos, N.T.; Berois, N.; Osinaga, E.; Magalhaes, A.; Pinto-de-Sousa, J.; Almeida, R.; Gartner, F.; Reis, C.A. Expression of UDP-N-acetyl-D-galactosamine: Polypeptide N-acetylgalactosaminyltransferase-6 in gastric mucosa, intestinal metaplasia, and gastric carcinoma. J. Histochem. Cytochem. 2009, 57, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Hung, J.S.; Huang, J.; Lin, Y.C.; Huang, M.J.; Lee, P.H.; Lai, H.S. C1GALT1 overexpression promotes the invasive behavior of colon cancer cells through modifying O-glycosylation of FGFR2. Oncotarget 2014, 5, 2096–2106. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.H.; Huang, N.J.; Chen, C.H.; Shyu, M.K.; Huang, J.; Hung, J.S.; Huang, C.S.; Huang, M.C. Up-regulation of C1GalT1 promotes breast cancer cell growth through MUC1-C signaling pathway. Oncotarget 2015, 6, 6123–6135. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Hirohashi, Y.; Murai, A.; Nishidate, T.; Okita, K.; Wang, L.; Ikehara, Y.; Satoyoshi, T.; Usui, A.; Kubo, T.; et al. ST6GALNA1 plays important roles in enhancing cancer stem phenotypes of colorectal cancer via the Akt patway. Oncotarget 2017, 8, 112550–112564. [Google Scholar] [CrossRef] [Green Version]
- Pinho, S.S.; Carvalho, S.; Marcos-Pinto, R.; Magalhaes, A.; Oliveira, C.; Gu, J.; Dinis-Ribeiro, M.; Carneiro, F.; Seruca, R.; Reis, C.A. Gastric cancer: Adding glycosylation to the equation. Trends Mol. Med. 2013, 19, 664–676. [Google Scholar] [CrossRef] [PubMed]
- Blanas, A.; Sahasrabudhe, N.M.; Rodrigez, E.; Kooyk, Y.; Vilet, S.J. Fucosylated antigens in cancer: An alliance toward tumor progression, metastasis, and resistance to chemotherapy. Front. Oncol. 2018, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, E.; Moriwaki, K.; Terao, N.; Tan, C.C.; Terao, M.; Nakagawa, T.; Matsumoto, H.; Shinzaki, S.; Kamada, Y. Fucosylation is a promising target for cancer diagnosis and therapy. Biomolecules 2012, 2, 34–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Lee, S.H.; Kawasaki, N.; Itoh, S.; Kang, K.; Ryu, S.H.; Hashii, N.; Kim, J.M.; Kim, J.Y.; Kim, J.H. α1-3/4 fucosylation at Asn 241 of β-haptoglobin is a novel marker for colon cancer: A combinatorial approach for development of glycan biomarkers. Int. J. Cancer 2012, 130, 2366–2376. [Google Scholar] [CrossRef] [PubMed]
- Bieg, D.; Sypniewski, D.; Nowak, E.; Bednarek, I. Morin decreases galectin-3 expression and sensitizes ovarian cancer cells to cisplatin. Arch. Gynecol. Obstet. 2018, 298, 1181–1194. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, J.; Degraff, W.; Gazdar, A.; Minna, J.; Mitchell, J. Evaluation of a tetrazolium-based semi-automated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Clone | Source |
|---|---|---|
| Anti-MUC1; extracellular domain (mouse IgG) Anti-MUC1; cytoplasmic tail (Armenian hamster IgG) Anti-Lewisa (mouse IgG) Anti-Lewisb (mouse IgG) Anti-C1GalT1 (mouse IgG) Anti-ST3GalII (mouse IgG) Anti-GalNAcTL5 (mouse IgG) Anti-Gal-3 (mouse IgG) | BC2 CT2 7LE LWB01 F-31 34-K F-5 B2C10 | Abcam Abcam Santa Cruz Thermo Scientific Santa Cruz Santa Cruz Santa Cruz Santa Cruz |
| Anti-β-actin (rabbit IgG) Anti-mouse IgG peroxidase conjugated Anti-rabbit IgG peroxidase conjugated Anti-Armenian hamster IgG peroxidase conjugated | Sigma Sigma Sigma Abcam |
| Gene | Forward Primer (5′ → 3′) | Reverse Primer (5′ → 3′) |
|---|---|---|
| Bax | TTGCTTCAGGGTTTCATCCA | CAGCCTTGAGCACCAGTTTG |
| Caspase-3 | CAGTGGAGGCCGACTTCTTG | TGGCACAAAGCGACTGGAT |
| Caspase-8 | TTTCTGCTGAAGTCCATCTTTTT | TAGGGGACTCGGAGACTGC |
| Caspase-9 | CCCATATGATCGAGGACATCCA | ACAACTTTGCTGCTTGCCTGTTAG |
| MUC1 | TGCCTTGGCTGTCTGTCAGT | GTAGGTATCCCGGGCTGGAA |
| C1GalT1 | AAGCAGGGCTACATGAGTGG | GCATCTCCCCAGTGCTAAGT |
| ppGalNAcT2 | AAGAAAGACCTTCATCACAGCAATGGAGAA | ATCAAAACCGCCCTTCAAGTCAGCA |
| ST6GalNAcT2 | CCTTCTGAACGGCTCAGAGAGT | GCACACCGGATACACTTTGGA |
| ST3GalT1 | TCGGCCTGGTTCGATGA | CGCGTTCTGGGCAGTCA |
| FUT3 | GCCGACCGCAAGGTGTAC | TGACTTAGGGTTGGACATGATATCC |
| FUT4 | AAGCCGTTGAGGCGGTTT | ACAGTTGTGTATGAGATTTGGAAGCT |
| Gal-3 | GCAGACAATTTTTCGCTCCATG | CTGTTGTTCTCATTGAAGCGTG |
| GAPDH | GTGAACCATGAGAAGTATGACAA | CATGAGTCCTTCCACGATAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radziejewska, I.; Supruniuk, K.; Czarnomysy, R.; Buzun, K.; Bielawska, A. Anti-Cancer Potential of Afzelin towards AGS Gastric Cancer Cells. Pharmaceuticals 2021, 14, 973. https://doi.org/10.3390/ph14100973
Radziejewska I, Supruniuk K, Czarnomysy R, Buzun K, Bielawska A. Anti-Cancer Potential of Afzelin towards AGS Gastric Cancer Cells. Pharmaceuticals. 2021; 14(10):973. https://doi.org/10.3390/ph14100973
Chicago/Turabian StyleRadziejewska, Iwona, Katarzyna Supruniuk, Robert Czarnomysy, Kamila Buzun, and Anna Bielawska. 2021. "Anti-Cancer Potential of Afzelin towards AGS Gastric Cancer Cells" Pharmaceuticals 14, no. 10: 973. https://doi.org/10.3390/ph14100973
APA StyleRadziejewska, I., Supruniuk, K., Czarnomysy, R., Buzun, K., & Bielawska, A. (2021). Anti-Cancer Potential of Afzelin towards AGS Gastric Cancer Cells. Pharmaceuticals, 14(10), 973. https://doi.org/10.3390/ph14100973