
Influence of Estrogenic Metabolic Pathway Genes Polymorphisms on Postmenopausal Breast Cancer Risk

 ,
,  ,
, 
Abstract
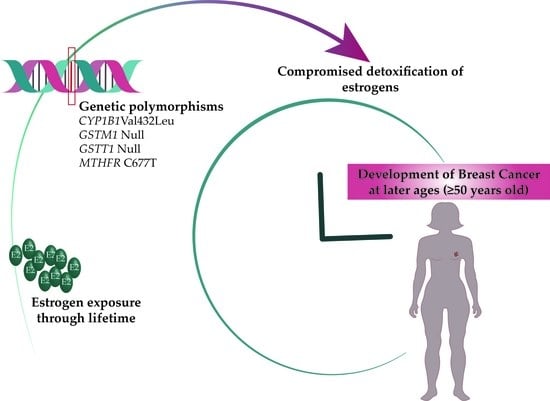
:
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. DNA Extraction
4.3. Genotyping
4.3.1. CYP1B1
4.3.2. MTHFR
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of incidence and mortality of cervical cancer in 2018: A worldwide analysis. Lancet Glob. Health 2019, 8, e191–e203. [Google Scholar] [CrossRef] [Green Version]
- Momenimovahed, Z.; Salehiniya, H. Epidemiological characteristics of and risk factors for breast cancer in the world. Breast Cancer 2019, 11, 151–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giudice, A.; Barbieri, A.; Bimonte, S.; Cascella, M.; Cuomo, A.; Crispo, A.; D’arena, G.; Galdiero, M.; Della Pepa, M.E.; Botti, G. Dissecting the prevention of estrogen-dependent breast carcinogenesis through Nrf2-dependent and independent mechanisms. Onco Targets Ther. 2019, 12, 4937–4953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yager, J.D.; Davidson, N.E. Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef] [Green Version]
- De Bock, G.H.; Mourits, M.J.E.; Schutte, M.; Krol-Warmerdam, E.M.M.; Seynaeve, C.; Blom, J.; Brekelmans, C.T.M.; Meijers-Heijboer, H.; van Asperen, C.J.; Cornelisse, C.J.; et al. Association between the CHEK2* 1100delc germ line mutation and estrogen receptor status. Int. J. Gynecol. Cancer 2006, 16, 552–555. [Google Scholar] [CrossRef]
- Cybulski, C.; Huzarski, T.; Byrski, T.; Gronwald, J.; Debniak, T.; Jakubowska, A.; Górski, B.; Wokołorczyk, D.; Masojć, B.; Narod, S.A.; et al. Estrogen receptor status in CHEK2-positive breast cancers: Implications for chemoprevention. Clin. Genet 2009, 75, 72–78. [Google Scholar] [CrossRef]
- Fanale, D.; Incorvaia, L.; Filorizzo, C.; Bono, M.; Fiorino, A.; Calò, V.; Brando, C.; Corsini, L.R.; Barraco, N.; Badalamenti, G.; et al. Detection of germline mutations in a cohort of 139 patients with bilateral breast cancer by multi-gene panel testing: Impact of pathogenic variants in other genes beyond BRCA1/2. Cancers 2020, 12, 2415. [Google Scholar] [CrossRef]
- Samavat, H.; Kurzer, M.S. Estrogen metabolism and breast cancer. Cancer Lett. 2015, 356, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Xie, D.W.; Jin, F.; Cheng, J.R.; Dai, Q.; Wen, W.Q.; Shu, X.O.; Gao, Y.T. Genetic polymorphism of cytochrome P450–1B1 and risk of breast cancer. Cancer Epidemiol. Biomark. Prev. 2000, 9, 147–150. [Google Scholar]
- Ghisari, M.; Long, M.; Bonefeld-Jørgensen, E.C. Genetic polymorphisms in CYP1A1, CYP1B1 and COMT genes in Greenlandic Inuit and Europeans. Int. J. Circumpolar Health 2013, 72, 21113. [Google Scholar] [CrossRef]
- Shimada, T.; Watanabe, J.; Kawajiri, K.; Sutter, T.R.; Guengerich, F.P.; Gillam, E.M.; Inoue, K. Catalytic properties of polymorphic human cytochrome P450 1B1 variants. Carcinogenesis 1999, 20, 1607–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.Y.; Chou, Y.C.; Wu, M.H.; Huang, H.B.; Jeng, Y.L.; Wu, C.C.; Yu, C.P.; Yu, J.C.; You, S.L.; Chu, T.Y.; et al. The MTHFR C677T polymorphism, estrogen exposure and breast cancer risk: A nested case-control study in Taiwan. Anticancer Res. 2004, 24, 3863–3868. [Google Scholar] [PubMed]
- Chen, X.; Ahamada, H.; Zhang, T.; Bai, Z.; Wang, C. Association of Intake Folate and Related Gene Polymorphisms with Breast Cancer. J. Nutr. Sci. Vitaminol. (Tokyo) 2019, 65, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, I.; Tran, P.; Christensen, B.; Sibani, S.; Rozen, R. A second genetic polymorphism in methylenetetrahydrofolate reductase (MTHFR) associated with decreased enzyme activity. Mol. Genet Metab. 1998, 64, 169–172. [Google Scholar] [CrossRef] [PubMed]
- AL-Eitan, L.N.; Rababa’h, D.M.; Alghamdi, M.A.; Khasawneh, R.H. Association of GSTM1, GSTT1 and GSTP1 polymorphisms with breast Cancer among Jordanian women. Onco Targets Ther. 2019, 12, 7757–7765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalhinho, A.C.; Fonseca-Moutinho, J.A.; Breitenfeld, L. Glutathione S-transferase M1, T1, and P1 genotypes and breast cancer risk: A study in a Portuguese population. Mol. Cell Biochem. 2011, 355, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Ramalhinho, A.C.; Fonseca-Moutinho, J.A.; Breitenfeld, L. Positive association of polymorphisms in estrogen biosynthesis gene, CYP19A1, and metabolism, GST, in breast cancer susceptibility. DNA Cell Biol. 2012, 31, 1100–1106. [Google Scholar] [CrossRef]
- Pearson, W.R.; Vorachek, W.R.; Xu, S.; Berger, R.; Hart, I.; Vannais, D.; Patterson, D. Identification of class-mu glutathione transferase genes GSTM1-GSTM5 on human chromosome 1p13. Am. J. Hum. Genet 1993, 53, 220–233. [Google Scholar]
- Xu, S.J.; Wang, Y.; Roe, B.; Pearson, W.R. Characterization of the human class Mu glutathione S-transferase gene cluster and the GSTM1 deletion. J. Biol. Chem. 1998, 273, 3517–3527. [Google Scholar] [CrossRef] [Green Version]
- Sprenger, R.; Schlagenhaufer, R.; Kerb, R.; Bruhn, C.; Brockmöller, J.; Roots, I.; Brinkmann, U. Characterization of the glutathione S-transferase GSTT1 deletion: Discrimination of all genotypes by polymerase chain reaction indicates a trimodular genotype–phenotype correlation. Pharmacogenetics 2000, 10, 557–565. [Google Scholar] [CrossRef]
- Kalimutho, M.; Nones, K.; Srihari, S.; Duijf, P.H.; Waddell, N.; Khanna, K.K. Patterns of genomic instability in breast cancer. Trends Pharmacol. Sci. 2019, 40, 198–211. [Google Scholar] [CrossRef]
- Fan, P.; Jordan, V.C. New insights into acquired endocrine resistance of breast cancer. Cancer Drug Resist 2019, 2, 198–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, C.; Wu, L.; Fu, L.; Wang, B.; Zhou, H. Unifying mechanism in the initiation of breast cancer by metabolism of estrogen. Mol. Med. Rep. 2017, 16, 1001–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, T.L.; Rae, J.M.; Colacino, J.A. Implication of environmental estrogens on breast cancer treatment and progression. Toxicology 2019, 421, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Pilleron, S.; Sarfati, D.; Janssen-Heijnen, M.; Vignat, J.; Ferlay, J.; Bray, F.; Soerjomataram, I. Global cancer incidence in older adults, 2012 and 2035: A population-based study. Int. J. Cancer 2019, 144, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Reljic, A.; Simundic, A.-M.; Topic, E.; Nikolac, N.; Justinic, D.; Stefanovic, M. The methylenetetrahydrofolate reductase (MTHFR) C677T polymorphism and cancer risk: The Croatian case–control study. Clin. Biochem. 2007, 40, 981–985. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Patients n (%) | Mean (SD, Min–Max) | |
|---|---|---|
| Total | 157 | |
| Age | 63.71 (13.777, min 34–max 95) | |
| <50 | 31 (19.75%) | 43.65 (4.270, min 34–max 49) |
| ≥50 | 126 (80.25%) | 68.65 (10.395, min 50–max 95) |
| Genotype | Age, n (%) | OR (95% CI) a | p-Value | |
|---|---|---|---|---|
| <50 | ≥50 | |||
| GSTT1 | ||||
| Present | 27 (17.2) | 83 (52.9) | 1 | |
| Null | 4 (2.5) | 43 (27.4) | 3.497 (1.149–10.641) | 0.021 * |
| GSTM1 | ||||
| Present | 17 (10.8) | 48 (30.6) | 1 | |
| Null | 14 (8.9) | 78 (49.7) | 1.973 (0.892–4.363) | 0.090 |
| GSTT1 | GSTM1 | Age, n (%) | OR (95% CI) a | p-Value | |
|---|---|---|---|---|---|
| <50 | ≥50 | ||||
| Present | Present | 16 (10.2) | 34 (21.7) | 1 | |
| Present | Null | 11 (7) | 49 (31.2) | 2.096 (0.866–5.072) | 0.097 |
| Null | Present | 1 (0.6) | 14 (8.9) | 6.588 (0.796–54.558) | 0.050 * |
| Null | Null | 3 (1.9) | 29 (18.5) | 4.549 (1.204–17.181) | 0.018 * |
| GSTT1 | CYP1B1 Val432Leu | Age, n (%) | OR (95% CI) a | p-Value | |
|---|---|---|---|---|---|
| <50 | ≥50 | ||||
| Present | Leu/Leu (WT) | 10 (6.4) | 21 (13.4) | 1 | |
| Present | Leu/Val + Val/Val | 17 (10.8) | 62 (39.5) | 1.737 (0.689–4.378) | 0.239 |
| Null | Leu/Leu (WT) | 0 | 8 (5.1) | NA b | 0.062 |
| Null | Leu/Val + Val/Val | 4 (2.5) | 35 (22.3) | 4.167 (1.159–14.979) | 0.022 * |
| GSTM1 | CYP1B1 Val432Leu | Age, n (%) | OR (95% CI) a | p-Value | |
|---|---|---|---|---|---|
| <50 | ≥50 | ||||
| Present | Leu/Leu (WT) | 8 (5.1) | 10 (6.4) | 1 | |
| Present | Leu/Val + Val/Val | 9 (5.7) | 38 (24.2) | 3.378 (1.038–10.992) | 0.038 * |
| Null | Leu/Leu (WT) | 2 (1.3) | 19 (12.1) | 7.600 (1.350–42.799) | 0.013 * |
| Null | Leu/Val + Val/Val | 12 (7.6) | 59 (37.6) | 3.933 (1.286–12.029) | 0.012 * |
| GSTT1 | MTHFR C677T | Age, n (%) | OR (95% CI) a | p-Value | |
|---|---|---|---|---|---|
| <50 | ≥50 | ||||
| Present | CC (WT) | 10 (6.4) | 30 (19.1) | 1 | |
| Present | CT+TT | 17 (10.8) | 53 (33.8) | 1.039 (0.422–2.557) | 0.933 |
| Null | CC (WT) | 2 (1.3) | 13 (8.3) | 2.167 (0.415–11.302) | 0.351 |
| Null | CT+TT | 2 (1.3) | 30 (19.1) | 5 (1.009–24.773) | 0.034 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, M.; Soares, M.; Fonseca-Moutinho, J.; Ramalhinho, A.C.; Breitenfeld, L. Influence of Estrogenic Metabolic Pathway Genes Polymorphisms on Postmenopausal Breast Cancer Risk. Pharmaceuticals 2021, 14, 94. https://doi.org/10.3390/ph14020094
Almeida M, Soares M, Fonseca-Moutinho J, Ramalhinho AC, Breitenfeld L. Influence of Estrogenic Metabolic Pathway Genes Polymorphisms on Postmenopausal Breast Cancer Risk. Pharmaceuticals. 2021; 14(2):94. https://doi.org/10.3390/ph14020094
Chicago/Turabian StyleAlmeida, Micaela, Mafalda Soares, José Fonseca-Moutinho, Ana Cristina Ramalhinho, and Luiza Breitenfeld. 2021. "Influence of Estrogenic Metabolic Pathway Genes Polymorphisms on Postmenopausal Breast Cancer Risk" Pharmaceuticals 14, no. 2: 94. https://doi.org/10.3390/ph14020094
APA StyleAlmeida, M., Soares, M., Fonseca-Moutinho, J., Ramalhinho, A. C., & Breitenfeld, L. (2021). Influence of Estrogenic Metabolic Pathway Genes Polymorphisms on Postmenopausal Breast Cancer Risk. Pharmaceuticals, 14(2), 94. https://doi.org/10.3390/ph14020094