
Zebrafish as a Screening Model to Study the Single and Joint Effects of Antibiotics †

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Effects of Individual Antibiotics in Zebrafish

{kind=link}
{kind=link}
{kind=link}
| Class. | Antibiotics | Concentrations (Weight/Water Volume) | Exposure Period | Lifespan Stages | Effects/Toxicity Observed | Refs. |
|---|---|---|---|---|---|---|
| Aminoglycosides | Netilmicin | 10–1000 ng/mL | 3 days at 6–72 hpf and at 3–6 dpf | Embryos at 6 hpf and larvae at 3 dpf | - Cardiotoxicity - Mild teratogenicity | [54] |
| Gentamicin | Injected with defined volumes of 10 mg/mL | From 55 to 72 and 96 hpf | Larvae | - Nephrotoxicity | [53] | |
| 0.2, 1, 2 and 5 μM | 24 h | 5 dpf | - Ototoxicity | [75] | ||
| 5, 10 and 20 μM | 24 h | 5 dpf | - Behavioral changes | [33] | ||
| 10 μM | 48 h | 3 dpf | - Oculotoxicity | [76] | ||
| 0.1–400 μM | Acute and chronic exposure | 5–6 dpf | - Ototoxicity | [77] | ||
| Neomycin | 0.16, 1.6, 8 and 16 μM | 24 h | 5 dpf | - Ototoxicity | [75] | |
| 125 μM | 1 h | 5 dpf | - Behavioral changes | [33] | ||
| 0.1–400 μM | Acute and chronic exposure | 5–6 dpf | - Ototoxicity | [77] | ||
| Streptomycin | 0.1–400 μM | Acute and chronic exposure | 5–6 dpf | - Ototoxicity | [77] | |
| 0.1, 1 and 10 μg/mL | 10 days | 5 dpf | - Dysbiosis - Early mortality | [78] | ||
| Etimicin | Survival test: 2, 5, 10, 20,50, 100, 200 and 500 mg/L | At 24, 48, 72, 96 and 120 hpf | 6 hpf | - Low nephrotoxicity and ototoxicity compared with amikacin and gentamicin | [79] | |
| Amikacin | Hatching and toxicity: 750, 1500 or 2000 mg/L | From 1 to 3 dpf | 1 dpf | - More developmental toxicities to the embryos than gentamicin | ||
| β-Lactams | Amoxicillin | 100 mg/L | 7 days | Young zebrafish | - Behavioral changes - Oxidative stress | [24] |
| 0, 75, 128, 221, 380, 654 and 1125 mg/L | 96 h | Embryos | - Developmental toxicity - Oxidative stress | [23] | ||
| 0, 1, 10, 25, 50 and 100 mg/L | Adults | |||||
| Cefotaxime | 10–1000 ng/mL | 3 days at 6–72 hpf and at 3–6 dpf | Embryos at 6 hpf and larvae at 3 dpf | - Cardiotoxicity - Teratogenicity increased in a dose-dependent manner | [54] | |
| 0, 10, 20 and 30 mM | 1 day | 5 dpf | - Locomotor toxicity - Abnormal expression of different genes | [80] | ||
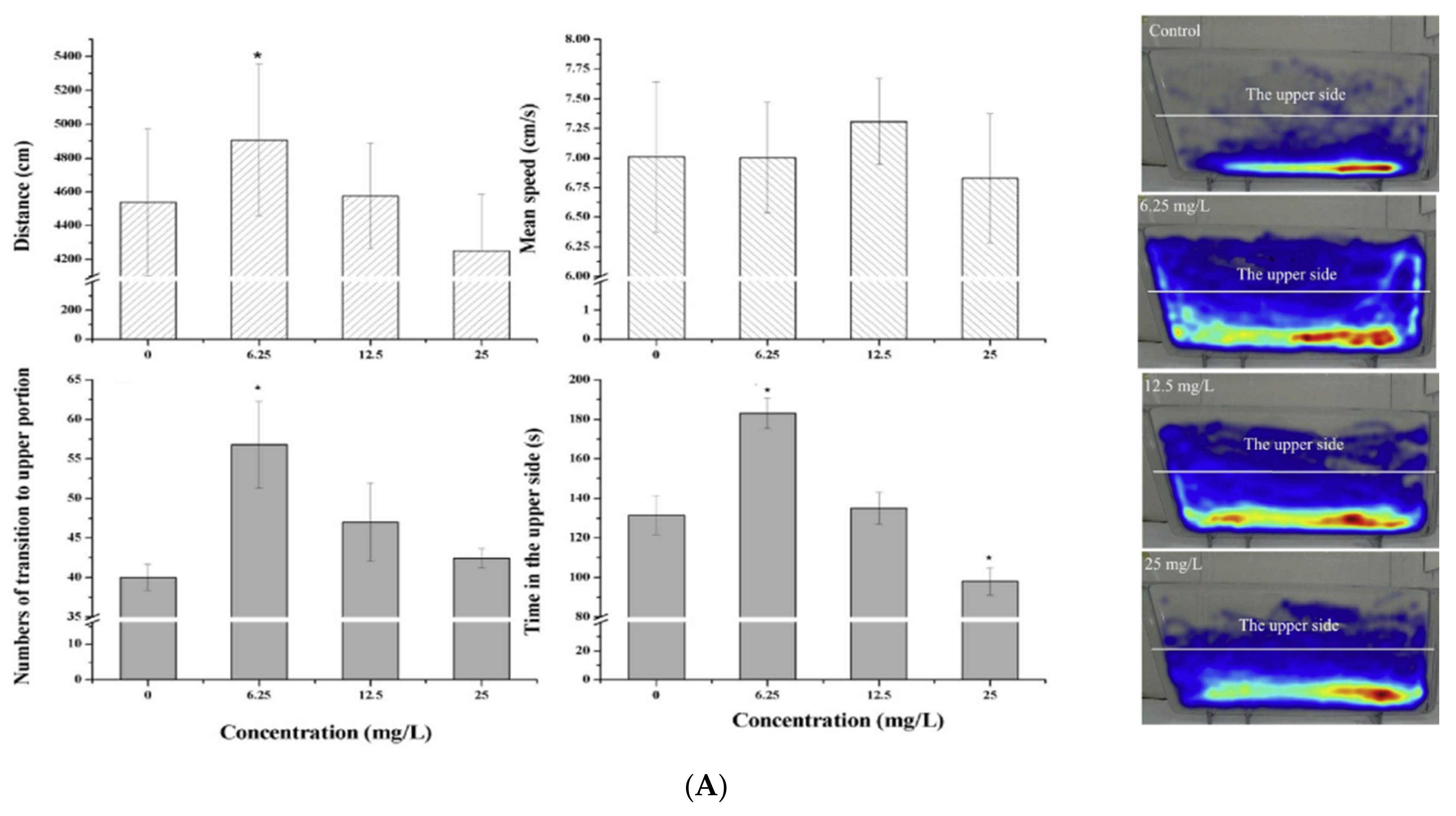
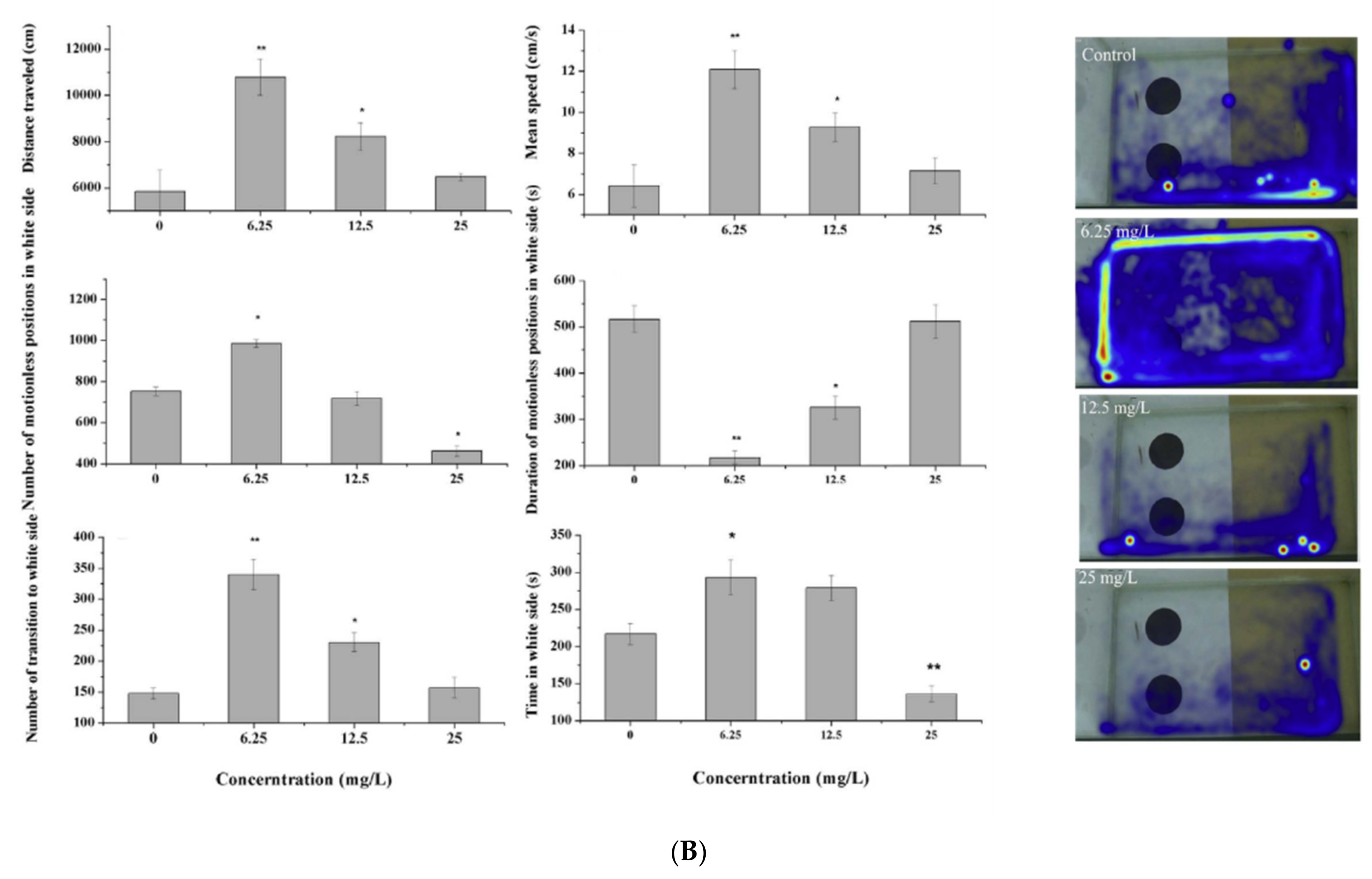
| Ceftazidime | 0, 6.25, 12.5, 25 mg/L | 96 h | Adult | - Behavioral changes | [34] | |
| 10–1000 ng/mL | 3 days at 6–72 hpf and at 3–6 dpf | Embryos at 6 hpf and larvae at 3 dpf | - Cardiotoxicity | [54] | ||
| Macrolides | Erythromycin, clarithromycin, azithromycin, midecamycin, josamycin | 0, 0.1, 1, 10, 100 and 1000 μM | 48 h until 5 dpf | 3 dpf | - Hepatotoxicity | [81] |
| 0, 0.5, 1, 2 | 6 dpf | 3 dpf | - Hepatotoxicity | [82] | ||
| acetylspiramycin | 0, 0.25, 0.5, 1 | 6 dpf | 3 dpf | - Hepatotoxicity | [82] | |
| Erythromycin | 0, 0.017, 0.034, 0.068, 0.136, 0.272 mM | 96 h | 24 hpf | - Cardiotoxicity | [27] | |
| 0, 2, 20, 200 and 2000 μg/L | 96 h | adult | - Abnormal gene expression - Behavioral changes | [68] | ||
| 0, 0.001, 0.01, 0.1, 1 and 10 μg/L | 96 hpf | 2 hpf | - Cardiotoxicity - Developmental toxicity - Enhanced swimming activity | [29] | ||
| Tilmicosin | 0, 0.63, 1.25, 2.5, 5, 10, 20 and 40 mg/L | 4 dpf | 2 dpf | - Developmental toxicity - Cardiotoxicity - Teratogenic effects - Oxidative stress - Apoptosis in embryos | [36] | |
| Quinolones | Ciprofloxacin | 0, 6.25, 12.5, 25 mg/L | 96 h | Adult | - Behavioral changes | [34] |
| 5 μg/L | 6–9 dpf | Larvae | - Increased expression of immune system cytokine genes | [83] | ||
| Norfloxacin | 0, 2, 20 and 200 μg/L | 3 weeks | Larvae | - Reproductive toxicity | [84] | |
| 600, 900, 1200 mg/L | 72 hpf | Embryos | - Neurotoxicity - Impaired hatching rate - Increased mortality - Malformation | [85] | ||
| Levofloxacin, gatifloxacin, norfloxacin, sparfloxacin, gemifloxacin, enoxacin, pefloxacin, prulifloxacin, lomefloxacin, moxifloxacin, ciprofloxacin, antofloxacin, 2-methylpiperazine, n-methylpiperazine, 4-n-Boc-2-methyl-piperazine | 0, 0.1, 0.2, 0.5, 1, 2, 5, 10, 20 mM | 6–72 hpf | Embryos | - Head deformation - Shortened tail - Tail tip malformation - Scoliosis and spinal curvature—reduced pigmentation - Developmental retardation - Pericardial edema - Heart malformation - Death | [86] | |
| Enrofloxacin | 0.01, 1, 100 μg/L | 120 h | Larvae | - Decreased body lengths - Deformed body shape - Disrupted metabolic processes | [87] | |
| Ofloxacin, Enrofloxacin | 5 μg/L | 20 days | Adult | - Accumulation in liver, skin, muscles and gills | [88] | |
| Gatifloxacin, Ciprofloxacin | - 413, 1238, 3713, 4239 mg/L - 156, 469, 1407 and 1949 mg/L | 24 h | Adult | - Cardiotoxicity | [89] | |
| Sulfonamides | Sulfamethoxazole | 0, 0.09, 0.19, 0.39, 0.79, 1.58 mM | 96 h | 24 hpf | - Developmental toxicity - Cardiotoxicity (pericardial edema, bradycardia) - Oxidative stress | [27] |
| 80 and 100 mg/kg body weight per day | 6 weeks | Adult | - Higher digestive enzyme activities | [41] | ||
| 0, 50, 100 and 500 μg/L | 14 days | Adult | - Lipid peroxidation | [90] | ||
| 260 ng/L | 6 weeks | Adult | - Impair the gut health - Higher metabolic rate | [91] | ||
| 0, 2, 20 and 200 μg/L | 3 weeks | Larvae | - Developmental toxicity - Reproductive toxicity - Oxidative stress - Impact on the development of the zebrafish offspring | [84] | ||
| 0, 0.1, 1, 10 and 100 μg/L | 120 h | 5 hpf | - Developmental toxicityoxidative stress - Inflammation | [92] | ||
| Sulfamethoxazole, sulfapyridine, sulfadiazine, sulfameter and sulfamerazine | 0, 20, 40, 80 and 160 μg/L | 24 and 168 hpf | Embryos | - Developmental toxicity - Cardiotoxicity | [93] | |
| Sulfamethazine | 0, 0.2, 20 and 2000 μg/L | 120 hpf | Embryo | - Developmental toxicity - Cardiotoxicity - Oxidative stress - Lipid peroxidation | [94] | |
| 1: 7 days exposure, 7 days post-exposure 2: 1 day exposure, 2 days resting and 4 days re-exposure | Adult | |||||
| Tetracyclines | Oxytetracycline | 0, 0.1, 10 and 10,000 μg/L | 2 months | Adult | - Behavioral changes (boldness and hyperactivity) - Impairments at biochemical level - Alteration of bacterial communities of fish gut | [25] |
| 80 and 100 mg/kg body weight per day | 6 weeks | Adult | - Higher digestive enzyme activities - More oxygen consumption rate | [41] | ||
| 1 and 5 ng/L | From 2 hpf to 120 dpf | Embryo | - Thyroid dysfunction - Developmental toxicity | [95] | ||
| 0, 75, 100, 150, 300, 600 and 900 mg/L | 96 h | Embryos | - Developmental toxicity - Oxidative stress | [23] | ||
| 0, 1, 10, 25, 50 and 100 mg/L | Adult | |||||
| 0, 0.05, 0.5 and 5 mg/L | 48 h | 72 hpf | - Oxidative stress | [96] | ||
| 420 ng/L | 6 weeks | Adult | - Impair the gut health - Higher metabolic rate | [91] | ||
| 10 μg/L | 5 days and 2 months exposure 5 days and 1 month of post-exposure | Adult | - The effects are partially reversible | [43] | ||
| Tetracycline HCl | 0, 1 and 100 μg/L | 1 month | Juvenile | - Body weight increase - No change in the body length | [42] | |
| 0, 0.1, 1, 10, 100 and 1000 μM | 48 h until 5 dpf | 3 dpf | - Hepatotoxicity | [81] | ||
| 0, 2, 10, 20, 200, 2000 and 20,000 μg/L | 96 h | 4 hpf | - Developmental toxicity - Oxidative stress | [44] | ||
| Minocycline | 10–1000 ng/mL | 3 days at 6–72 hpf and at 3–6 dpf | Embryos at 6 hpf and larvae at 3 dpf | - Fish died at 0.4 mg/mL | [54] | |
| Chlortetracycline | 0, 6.25, 12.5, 25 mg/L | 96 h | Adult | - Behavioral changes | [34] | |
| 0, 0.2, 2 and 20 mg/L | 48 h | 72 hpf | - Oxidative stress | [96] | ||
| 0, 6.25, 12.5, 25 mg/L | 96 h | Adult | - Behavioral changes | [34] | ||
| Polyether antibiotics | Maduramicin | 0, 0.1, 0.5 and 2.5 mg/L | 14 days | Adult | - Oxidative stress - Tissue damage in the gill, liver and intestine | [22] |
| 0, 10, 11.89, 14.14, 16.81 and 20 mg/L | 96 h | Adult | - Impact on the survival - LC50 = 13.568 mg/L | |||
| Monensin | 4, 4.34, 4.7, 5.1,5.53 mg/L | 96 h | Adult | - LC50 = 4.76 mg/L | [39] | |
| Others | Ceftazidime | 10–1000 ng/mL | 3 days at 6–72 hpf and at 3–6 dpf | Embryos at 6 hpf and larvae at 3 dpf | - Cardiotoxicity | [54] |
2.1. Aminoglycosides
2.2. β-Lactams
2.3. Macrolides
2.4. (Fluoro) Quinolones
2.5. Sulfonamides
2.6. Tetracyclines
2.7. Polyether Antibiotics
3. Effects of Antibiotic Mixtures in Zebrafish
| Class | Antibiotics | Mixture Concentrations (Weight/Water Volume) | Exposure Period | Lifespan Stages | Effects Observed | Refs. |
|---|---|---|---|---|---|---|
| β-Diketones (fluoroquinolones and tetracyclines) | Ofloxacin, ciprofloxacin, enrofloxacin, doxycycline, chlortetracycline and oxytetracycline | 0, 6.25 and 12.5 mg/L | From embryos (4 hpf) to larvae (90 dpf) stage | Larvae and adult | - Abnormal expression of differentially expressed miRNAs - Vacuolation of interstitial cells, reduced number of neurons, glial cell proliferation and deformation of glial scar | [113] |
| 0, 12.5 and 25 mg/L | From embryos (2hpf) to larvae (5dpf) stage | 72 hpf or 120 hpf | - Ototoxicity | [31] | ||
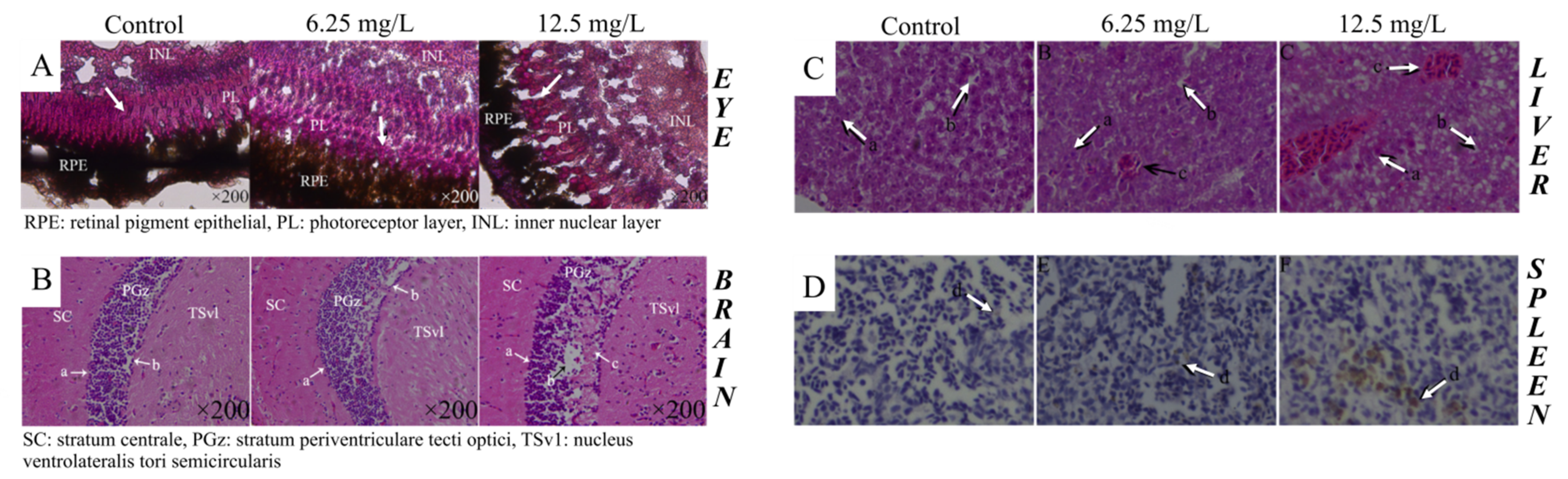
| 0, 6.25 and 12.5 mg/L | 3 months | 90 dpf | - Immunotoxicity (abnormal expression of immune genes and enzymes and variable levels of damage to immune-related organs) | [112] | ||
| 0, 6.25, 12.5 and 25 mg/L | 3 months | Embryos at 6 hpf | - Neurotoxicity (behavioral abnormality and anxiety, pathological changes of nerve cells, changes in appb and cdh6 transcriptional level) | [28] | ||
| FQs: ciprofloxacin, ofloxacin, norfloxacin, enrofloxacin, | 0, 25, 50, 100, 200, 300, 400 and 600 mg/L | 6–120 hpf | 72 hpf or120 hpf | - Abnormal hatching - Mortality- Malformation | [37] | |
| TCs: chlortetracycline and doxycycline | 0, 1.56, 3.13, 6.25, 12.5, 25 and 50 mg/L | |||||
| FQs + TCs | 0, 4.69, 9.38, 18.75, 37.5, 75, 150, 300 and 450 mg/L | |||||
| FQs: | 0, 12.5, 25, 50 mg/L | 6–96 hpf | 144 hpf | - Higher and basal swimming speed | ||
| TCs: | 0, 1.56, 3.13, 6.25 mg/L | |||||
| FQs + TCs | 0, 4.69, 9.38, 18.75 mg/L | |||||
| FQs + TCs | 0, 9.38, 18.75, 37.5, 75, 150 mg/L | 6–72 hpf | 48, 60, 72 hpf | - Decreased heart rate | ||
| FQs + TCs | 0, 45, 60, 95 mg/L | 2–4 months | - | - Severe edema in sarcoplasmic reticulum, melted muscle fiber and edema in mitochondria (skeletal muscle) - Disordered arrangement of muscle fibers, melted fiber, partial edematous membrane cell nuclear materials, edema in mitochondria, abnormal mitochondria (heart) - Changes in transcriptional levels of acta1a, myl7 and gle1b genes, which are involved in heart development and skeletal muscle formation | ||
| Ciprofloxacin, ofloxacin, enrofloxacin, oxytetracycline, chlortetracycline and doxycycline | 0, 6.25, 12.5 and 25 mg/L | 6 hpf until 144 hpf | 90 dpfwild-type adult zebrafish | - Physiological impairment - Reproductive toxicity | [114] | |
| Ciprofloxacin, ofloxacin, norfloxacin, enrofloxacin, chlortetracycline and doxycycline | 0, 18.75, 37.5, 75, 150, and 300 mg/L | From 6 hpf to 120 hpf | Studied every 12 h using a microscope | - Abnormal hatching - Malformation and mortality - Decreased heart rate | [117] | |
| 0, 2.34, 9.38 and 37.5 mg/L | From 6 hpf to 96 hpf | Embryos (<72 hpf) and larvae (>72 hpf) | - Locomotor toxicity - Oxidative stress (SOD and GSH) | |||
| Ciprofloxacin, ofloxacin, enrofloxacin, oxytetracycline, chlortetracycline and doxycycline | 0, 6.25, 12.5 mg/L | From 6 hpf to 90 dpf | 90 dpf | - Physiological impairment - Reproductive toxicity | [32] | |
| - Abnormal expression of some lncRNAs and their regulating target genes - Liver and spleen toxicity | [115] | |||||
| Ciprofloxacin, ofloxacin, norfloxacin, enrofloxacin, chlortetracycline and doxycycline | 0, 9.38 mg/L | From 6 hpf to 144 hpf | 90 dpf | - 47 differential expression proteins vs. control with 14 up-regulated and 12 down-regulated | [30] | |
| 0, 4.69, 9.38, 18.75 and 37.5 mg/L | From 6 hpf to 96 hpf | 120 hpf | - No visible developmental malformation - Greater spontaneous movement for low dose | |||
| 9.38, 45 and 60 and 90 mg/L | From 6 hpf to 90 dpf | 90 dpf | - Changes in creatine kinase activity and creatinine concentration - Changes in heart tissue reflected by dissolution of cristae and vacuolation of mitochondria | |||
| 0, 9.38, 18.75, 37.5, 75 and 150 mg/L | From 6 hpf to 72 hpf | 48,60 and 72 hpf | - Decreased heart rate | |||
| 0, 18.75, 37.5, 75, 150, 300 and 450 mg/L | From 6 hpf to 120 hpf | 72 hpf120 hpf | - 72 hpf EC50 for hatching rate = 130.3 mg/L - 120 hpf EC50 for malformation rate = 135.1 mg/L - 120 hpf LC50 for malformation rate = 149.8 mg/L - Severe malformation | [38] | ||
| 0, 45, 60 and 90 mg/L | 60 dpf | 7, 14, 21 days | - Changes in creatine kinase activity and creatinine concentration | |||
| 9.38 mg/L | From 6 hpf to 6 days | 90 dpf | - Serious liver damage | [35] | ||
| 2.34, 9.38 and 37.5 mg/L | - Oxidative stress (SOD and GSH) | |||||
| Sulfonamides (binary mixtures) | Sulfamethoxazole, sulfapyridine, sulfadiazine, sulfameter and sulfamerazine | 0, 20, 40, 80 and 160 μg/L equi-toxic ratio | 24 and 168 hpf | Embryos | - Developmental toxicity - Cardiotoxicity | [93] |
| Sulfonamides, β-lactams, tetracyclines and quinolones | Sulfamonomethoxine cefotaxime sodium tetracycline enrofloxacin | 0.01, 1 and 100 μg/Lin equal proportions | 120 h | Embryos at 4 hpf | - No significant differences in the mortality - Decreased the body lengths - Changes in the mRNA transcription profiles | [83] |
| Macrolides, amphenicols and sulfonamides | Clarithromycin, florfenicol, sulfamethazine | 0.1 mg/L | 96 h | Embryo | - No visible morphological changes - Behavioral and metabolic effects | [40] |
| - | 5 dpf | |||||
| 72 h | Adult | |||||
| Macrolides, lincosamides, quinolones, sulfonamides, tetracyclines and other | Clarithromycin, erythromycin, roxithromycin, lincomycin, ciprofloxacin, enrofloxacin, norfloxacin, ofloxacin, sulfadiazine, sulfamethazine, sulfamethoxazole, trimethoprim, oxytetracycline, chlortetracycline and tetracycline | 0.1 and100 μg/L | 4 weeks | 150 dpf gravid fish | - Reproductive effects - An antibiotic transfer from exposed adult fish to their offspring - Gastrointestinal effects in zebrafish offspring | [116] |
| β-lactams, aminoglycosides and macrolides | Ampicillin, kanamycin and amphotericin B | Mixture of 100 μg/mL AMP, 5 μg/mL KAN 250 ng/mL AMB | 2 weeks | Adult male zebrafish | - Alter the intestinal microbiome - Decrease intestinal and hepatic inflammation - Decrease hepatic steatosis in zebrafish with SBS | [119] |
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Yang, C.; Song, G.; Lim, W. A review of the toxicity in fish exposed to antibiotics. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 237, 108840. [Google Scholar] [CrossRef]
- Kovaláková, P.; Cizmas, L.; McDonald, T.J.; Marsalek, B.; Feng, M.; Sharma, V.K. Occurrence and toxicity of antibiotics in the aquatic environment: A review. Chemosphere 2020, 251, 126351. [Google Scholar] [CrossRef]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, I.T.; Santos, L. Antibiotics in the aquatic environments: A review of the European scenario. Environ. Int. 2016, 94, 736–757. [Google Scholar] [CrossRef] [PubMed]
- Zhai, G.; Ma, J. Antibiotic Contamination: A Global Environment Issue. J. Bioremediation Biodegradation 2014, 5, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Rashidian, G.; Boldaji, J.; Rainis, S.; Prokić, M.; Faggio, C. Oregano (Origanum vulgare) Extract Enhances Zebrafish (Danio rerio) Growth Performance, Serum and Mucus Innate Immune Responses and Resistance against Aeromonas hydrophila Challenge. Animals 2021, 11, 299. [Google Scholar] [CrossRef] [PubMed]
- Gholampour, T.E.; Raieni, R.F.; Pouladi, M.; Larijani, M.; Pagano, M.; Faggio, C. The Dietary Effect of Vitex agnus-castus Hydroalcoholic Extract on Growth Performance, Blood Biochemical Parameters, Carcass Quality, Sex Ratio and Gonad Histology in Zebrafish (Danio rerio). Appl. Sci. 2020, 10, 1402. [Google Scholar] [CrossRef] [Green Version]
- Ahmadifar, E.; Sheikhzadeh, N.; Roshanaei, K.; Dargahi, N.; Faggio, C. Can dietary ginger (Zingiber officinale) alter biochemical and immunological parameters and gene expression related to growth, immunity and antioxidant system in zebrafish (Danio rerio)? Aquaculture 2019, 507, 341–348. [Google Scholar] [CrossRef]
- Sehonova, P.; Tokanova, N.; Hodkovicova, N.; Kroupova, H.K.; Tumova, J.; Blahova, J.; Marsalek, P.; Plhalova, L.; Doubkova, V.; Dobsikova, R.; et al. Oxidative stress induced by fluoroquinolone enrofloxacin in zebrafish (Danio rerio) can be ameliorated after a prolonged exposure. Environ. Toxicol. Pharmacol. 2019, 67, 87–93. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Capillo, G.; Paknejad, H.; Khalili, M.; Tabarraei, A.; Van Doan, H.; Spanò, N.; Faggio, C. Mucosal immune parameters, immune and antioxidant defence related genes expression and growth performance of zebrafish (Danio rerio) fed on Gracilaria gracilis powder. Fish Shellfish. Immunol. 2018, 83, 232–237. [Google Scholar] [CrossRef]
- Petrovici, A.; Strungaru, S.-A.; Nicoara, M.; Robea, M.A.; Solcan, C.; Faggio, C. Toxicity of Deltamethrin to Zebrafish Gonads Revealed by Cellular Biomarkers. J. Mar. Sci. Eng. 2020, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Zon, L.I.; Peterson, R.T. In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discov. 2005, 4, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Cassar, S.; Adatto, I.; Freeman, J.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Kalueff, A.V.; Echevarria, D.J.; Homechaudhuri, S.; Stewart, A.M.; Collier, A.D.; Kaluyeva, A.A.; Li, S.; Liu, Y.; Chen, P.; Wang, J.; et al. Zebrafish neurobehavioral phenomics for aquatic neuropharmacology and toxicology research. Aquat. Toxicol. 2016, 170, 297–309. [Google Scholar] [CrossRef]
- McGrath, P.; Li, C.-Q. Zebrafish: A predictive model for assessing drug-induced toxicity. Drug Discov. Today 2008, 13, 394–401. [Google Scholar] [CrossRef]
- Fiorino, E.; Sehonova, P.; Plhalova, L.; Blahova, J.; Svobodova, Z.; Faggio, C. Effects of glyphosate on early life stages: Comparison between Cyprinus carpio and Danio rerio. Environ. Sci. Pollut. Res. 2018, 25, 8542–8549. [Google Scholar] [CrossRef]
- Plhalova, L.; Blahova, J.; Divisova, L.; Enevova, V.; Di Tocco, F.C.; Faggio, C.; Tichy, F.; Vecerek, V.; Svobodova, Z. The effects of subchronic exposure to NeemAzal T/S on zebrafish (Danio rerio). Chem. Ecol. 2017, 34, 199–210. [Google Scholar] [CrossRef]
- Bartoskova, M.; Dobsikova, R.; Stancova, V.; Zivna, D.; Blahova, J.; Marsalek, P.; Zelníckova, L.; Bartos, M.; Di Tocco, F.C.; Faggio, C. Evaluation of ibuprofen toxicity for zebrafish (Danio rerio) targeting on selected biomarkers of oxidative stress. Neuro Endocrinol. Lett. 2013, 34, 102–108. [Google Scholar] [PubMed]
- Plhalova, L.; Sehonova, P.; Blahova, J.; Doubkova, V.; Tichy, F.; Faggio, C.; Berankova, P.; Svobodova, Z. Evaluation of Tramadol Hydrochloride Toxicity to Juvenile Zebrafish—Morphological, Antioxidant and Histological Responses. Appl. Sci. 2020, 10, 2349. [Google Scholar] [CrossRef] [Green Version]
- Faria, M.; Prats, E.; Ramírez, J.R.R.; Bellot, M.; Bedrossiantz, J.; Pagano, M.; Valls, A.; Gomez-Canela, C.; Porta, J.M.; Mestres, J.; et al. Androgenic activation, impairment of the monoaminergic system and altered behavior in zebrafish larvae exposed to environmental concentrations of fenitrothion. Sci. Total. Environ. 2021, 775, 145671. [Google Scholar] [CrossRef]
- Ni, H.; Peng, L.; Gao, X.; Ji, H.; Ma, J.; Li, Y.; Jiang, S. Effects of maduramicin on adult zebrafish (Danio rerio): Acute toxicity, tissue damage and oxidative stress. Ecotoxicol. Environ. Saf. 2019, 168, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.; McDonough, S.; Ladewig, J.C.; Soares, A.M.; Nogueira, A.J.; Domingues, I. Effects of oxytetracycline and amoxicillin on development and biomarkers activities of zebrafish (Danio rerio). Environ. Toxicol. Pharmacol. 2013, 36, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.L.; Vasconcelos, F.F.P.; Wessler, L.B.; Lemos, I.S.; Candiotto, G.; Lin, J.; Matias, M.B.D.; Rico, E.P.; Streck, E.L. Exposure to a high dose of amoxicillin causes behavioral changes and oxidative stress in young zebrafish. Metab. Brain Dis. 2020, 35, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.R.; Tacão, M.; Machado, A.L.; Golovko, O.; Zlabek, V.; Domingues, I.; Henriques, I. Long-term effects of oxytetracycline exposure in zebrafish: A multi-level perspective. Chemosphere 2019, 222, 333–344. [Google Scholar] [CrossRef]
- Blahova, J.; Cocilovo, C.; Plhalova, L.; Svobodova, Z.; Faggio, C. Embryotoxicity of atrazine and its degradation products to early life stages of zebrafish (Danio rerio). Environ. Toxicol. Pharmacol. 2020, 77, 103370. [Google Scholar] [CrossRef]
- Aderemi, A.O. Developmental anomalies and oxidative stress responses in zebrafish (danio rerio) following embryonic exposure to human pharmaceuticals. J. Toxicol. Environ. Health 2020, 5, 109–125. [Google Scholar]
- Wang, X.; Zheng, Y.; Zhang, Y.; Li, J.; Zhang, H.; Wang, H. Effects of β-diketone antibiotic mixtures on behavior of zebrafish (Danio rerio). Chemosphere 2016, 144, 2195–2205. [Google Scholar] [CrossRef]
- Minski, V.T.; Garbinato, C.; Thiel, N.; Siebel, A.M. Erythromycin in the aquatic environment: Deleterious effects on the initial development of zebrafish. J. Toxicol. Environ. Heal. Part A 2021, 84, 56–66. [Google Scholar] [CrossRef]
- Yin, X.; Wang, H.; Zhang, Y.; Dahlgren, R.A.; Zhang, H.; Shi, M.; Gao, M.; Wang, X. Toxicological Assessment of Trace β-Diketone Antibiotic Mixtures on Zebrafish (Danio rerio) by Proteomic Analysis. PLoS ONE 2014, 9, e102731. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ling, Y.; Huang, W.; Sun, L.; Li, Y.; Wang, C.; Zhang, Y.; Wang, X.; Dahlgren, R.A.; Wang, H. Regulatory mechanisms of miR-96 and miR-184 abnormal expressions on otic vesicle development of zebrafish following exposure to β-diketone antibiotics. Chemosphere 2019, 214, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lin, J.; Huili, W.; Zhang, H.; Haifeng, Z.; Wang, X.; Dahlgren, R.A.; Wang, H. Effects of β-diketone antibiotics on F1-zebrafish (Danio rerio) based on high throughput miRNA sequencing under exposure to parents. Chemosphere 2016, 164, 41–51. [Google Scholar] [CrossRef]
- Han, E.; Oh, K.H.; Park, S.; Rah, Y.C.; Park, H.-C.; Koun, S.; Choi, J. Analysis of behavioral changes in zebrafish (Danio rerio) larvae caused by aminoglycoside-induced damage to the lateral line and muscles. NeuroToxicology 2020, 78, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.D.; Pereira, T.C.B.; Altenhofen, S.; Nabinger, D.D.; Ferreira, P.M.D.A.; Bogo, M.R.; Bonan, C.D. Antibiotic drugs alter zebrafish behavior. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 242, 108936. [Google Scholar] [CrossRef]
- Wang, H. Chronic toxicological effects of β-diketone antibiotics on Z ebrafish (D anio rerio) using transcriptome profiling of deep sequencing. Environ. Toxicol. 2016, 31, 1357–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Huang, X.; Xie, Y.; Song, M.; Zhu, K.; Ding, S. Macrolides induce severe cardiotoxicity and developmental toxicity in zebrafish embryos. Sci. Total. Environ. 2019, 649, 1414–1421. [Google Scholar] [CrossRef]
- Zhang, Y. Toxicity assessment of combined fluoroquinolone and tetracycline exposure in zebrafish (D anio rerio). Environ. Toxicol. 2016, 31, 736–750. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Zang, L.; Zhang, Y.; Zhang, Y.; Wang, X.; Ai, W.; Ding, N.; Wang, H. Joint toxicity of fluoroquinolone and tetracycline antibiotics to zebrafish (Danio rerio) based on biochemical biomarkers and histopathological observation. J. Toxicol. Sci. 2017, 42, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Li, P. Soil Behaviour of the Veterinary Drugs Lincomycin, Monensin, and Roxarsone and Their Toxicity on Environmental Or-ganisms. Molecules 2019, 24, 4465. [Google Scholar] [CrossRef] [Green Version]
- Ryan, B. Sub-lethal pharmaceutical hazard tracking in adult zebrafish using untargeted LC–MS environmental metabolomics. J. Hazard. Mater. 2017, 339, 63–72. [Google Scholar]
- Zhou, L.; Limbu, S.M.; Qiao, F.; Du, Z.-Y.; Zhang, M. Influence of Long-Term Feeding Antibiotics on the Gut Health of Zebrafish. Zebrafish 2018, 15, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Keerthisinghe, T.P.; Wang, F.; Wang, M.; Yang, Q.; Li, J.; Yang, J.; Xi, L.; Dong, W.; Fang, M. Long-term exposure to TET increases body weight of juvenile zebrafish as indicated in host metabolism and gut microbiome. Environ. Int. 2020, 139, 105705. [Google Scholar] [CrossRef]
- Almeida, A.R.; Domingues, I.; Henriques, I. Zebrafish and water microbiome recovery after oxytetracycline exposure. Environ. Pollut. 2021, 272, 116371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cheng, J.; Xin, Q. Effects of tetracycline on developmental toxicity and molecular responses in zebrafish (Danio rerio) embryos. Ecotoxicology 2015, 24, 707–719. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, S.; Chen, J.; Jiang, J.; Xie, Q.; Quan, X. Biological uptake and depuration of sulfadiazine and sulfamethoxazole in common carp (Cyprinus carpio). Chemosphere 2015, 120, 592–597. [Google Scholar] [CrossRef]
- Song, C.; Zhang, C.; Fan, L.; Qiu, L.; Wu, W.; Meng, S.; Hu, G.; Kamira, B.; Chen, J. Occurrence of antibiotics and their impacts to primary productivity in fishponds around Tai Lake, China. Chemosphere 2016, 161, 127–135. [Google Scholar] [CrossRef]
- Ben, W.; Zhu, B.; Yuan, X.; Zhang, Y.; Yang, M.; Qiang, Z. Occurrence, removal and risk of organic micropollutants in wastewater treatment plants across China: Comparison of wastewater treatment processes. Water Res. 2018, 130, 38–46. [Google Scholar] [CrossRef]
- Brown, K.D.; Kulis, J.; Thomson, B.; Chapman, T.H.; Mawhinney, D.B. Occurrence of antibiotics in hospital, residential, and dairy effluent, municipal wastewater, and the Rio Grande in New Mexico. Sci. Total. Environ. 2006, 366, 772–783. [Google Scholar] [CrossRef]
- Seifrtová, M. Determination of fluoroquinolone antibiotics in hospital and municipal wastewaters in Coimbra by liquid chroma-tography with a monolithic column and fluorescence detection. Anal. Bioanal. Chem. 2008, 391, 799–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, X.; Tian, H.; Wei, Q.; Liu, B.; Bao, G.; Liao, M.; Peng, J.; Huang, X.; Wang, L. High through-put determination of 28 veterinary antibiotic residues in swine wastewater by one-step dispersive solid phase extraction sample cleanup coupled with ultra-performance liquid chromatography-tandem mass spectrometry. Chemosphere 2019, 230, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Li, D. Determination and fate of oxytetracycline and related compounds in oxytetracycline production wastewater and the re-ceiving river. Environ. Toxicol. Chem. Int. J. 2008, 27, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Q.; Zhang, J.; Dong, J.; Yan, H.; Chen, C.; Feng, R. Characterization and source identification of tetracycline antibiotics in the drinking water sources of the lower Yangtze River. J. Environ. Manag. 2019, 244, 13–22. [Google Scholar] [CrossRef]
- Hentschel, D.M.; Park, K.M.; Cilenti, L.; Zervos, A.S.; Drummond, I.; Bonventre, J.V. Acute renal failure in zebrafish: A novel system to study a complex disease. Am. J. Physiol. Physiol. 2005, 288, F923–F929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Qin, W.; Zhang, J.-P.; Hu, C.-Q. Antibiotic Toxicity and Absorption in Zebrafish Using Liquid Chromatography-Tandem Mass Spectrometry. PLoS ONE 2015, 10, e0124805. [Google Scholar] [CrossRef]
- Ibrahim, M. Chapter 4—Current trends of antimicrobials used in food animals and aquaculture. In Antibiotics and Antimicrobial Resistance Genes in the Environment; Hashmi, M.Z., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 39–69. [Google Scholar]
- Larsson, D.G.J. Antibiotics in the environment. Upsala J. Med Sci. 2014, 119, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.; Roy, K. Risk assessment for ecotoxicity of pharmaceuticals–an emerging issue. Expert Opin. Drug Safety 2012, 11, 235–274. [Google Scholar] [CrossRef]
- Minguez, L.; Pedelucq, J.; Farcy, E.; Ballandonne, C.; Budzinski, H.; Halm-Lemeille, M.-P. Toxicities of 48 pharmaceuticals and their freshwater and marine environmental assessment in northwestern France. Environ. Sci. Pollut. Res. 2016, 23, 4992–5001. [Google Scholar] [CrossRef]
- Straub, J.O. Aquatic environmental risk assessment for human use of the old antibiotic sulfamethoxazole in Europe. Environ. Toxicol. Chem. 2015, 35, 767–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.; Ji, K.; Kho, Y.; Kim, P.-G.; Park, K.; Kim, K.; Kim, Y.; Kim, K.-T.; Choi, K. Effects of chronic exposure to cefadroxil and cefradine on Daphnia magna and Oryzias latipes. Chemosphere 2017, 185, 844–851. [Google Scholar] [CrossRef]
- Kim, J.-W.; Ishibashi, H.; Yamauchi, R.; Ichikawa, N.; Takao, Y.; Hirano, M.; Koga, M.; Arizono, K. Acute toxicity of pharmaceutical and personal care products on freshwater crustacean (Thamnocephalus platyurus) and fish (Oryzias latipes). J. Toxicol. Sci. 2009, 34, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.E.; Abdel-Daim, M.M. Modulating Effects of Spirulina platensis against Tilmicosin-Induced Cardiotoxicity in Mice. Cell J. 2015, 17, 137–144. [Google Scholar]
- Rodrigues, S.; Antunes, S.C.; Correia, A.; Nunes, B. Rainbow trout (Oncorhynchus mykiss) pro-oxidant and genotoxic responses following acute and chronic exposure to the antibiotic oxytetracycline. Ecotoxicology 2016, 26, 104–117. [Google Scholar] [CrossRef]
- Chen, M.; Zhao, H.; Wang, Y.; Bekele, T.G.; Liu, W.; Chen, J. Uptake and depuration of eight fluoroquinolones (FQs) in common carp (Cyprinus carpio). Ecotoxicol. Environ. Saf. 2019, 180, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, N.; Hashmi, I. Assessment of immunohematological, hematological and biochemical responses in cultivable fish Cyprinus carpio exposed to an antibiotic sulfamethoxazole (SMX). J. Water Heal. 2021, 19, 108–119. [Google Scholar] [CrossRef]
- Liu, J.; Lu, G.; Wu, D.; Yan, Z. A multi-biomarker assessment of single and combined effects of norfloxacin and sulfamethoxazole on male goldfish (Carassius auratus). Ecotoxicol. Environ. Saf. 2014, 102, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-J.; Choi, K.H.; Kim, M.Y.; Kim, P.G. Endocrine disruption induced by some sulfa drugs and tetracyclines on Oryzias latipes. J. Environ. Health Sci. 2006, 32, 227–234. [Google Scholar]
- Li, Y.; Zhang, J. The Effect of Acute Erythromycin Exposure on the Swimming Ability of Zebrafish (Danio rerio) and Medaka (Oryzias latipes). Int. J. Environ. Res. Public Heal. 2020, 17, 3389. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.-J.; Hou, J.-H.; Kuo, T.-F.; Lai, H.-T. Toxicity of the veterinary sulfonamide antibiotic sulfamonomethoxine to five aquatic organisms. Environ. Toxicol. Pharmacol. 2014, 38, 874–880. [Google Scholar] [CrossRef]
- Rodrigues, S.; Antunes, S.C.; Correia, A.; Golovko, O.; Žlábek, V.; Nunes, B. Assessment of toxic effects of the antibiotic erythromycin on the marine fish gilthead seabream (Sparus aurata L.) by a multi-biomarker approach. Chemosphere 2019, 216, 234–247. [Google Scholar] [CrossRef]
- Rodrigues, S.; Antunes, S.; Correia, A.; Nunes, B. Ecotoxicological evaluation of gilthead seabream (Sparus aurata) exposed to the antibiotic oxytetracycline using a multibiomarker approach. Mar. Environ. Res. 2018, 141, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.; Antunes, S.C.; Nunes, B.; Correia, A.T. Histopathological effects in gills and liver of Sparus aurata following acute and chronic exposures to erythromycin and oxytetracycline. Environ. Sci. Pollut. Res. 2019, 26, 15481–15495. [Google Scholar] [CrossRef]
- Wang, N. Adverse effects of enrofloxacin when associated with environmental stress in Tra catfish (Pangasianodon hy-pophthalmus). Chemosphere 2009, 77, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Varó, I.; Navarro, J.C.; Rigos, G.; Del Ramo, J.; Calduch-Giner, J.A.; Hernández, A.; Pertusa, J.; Torreblanca, A. Proteomic evaluation of potentiated sulfa treatment on gilthead sea bream (Sparus aurata L.) liver. Aquaculture 2013, 376-379, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Ton, C.; Parng, C. The use of zebrafish for assessing ototoxic and otoprotective agents. Hear. Res. 2005, 208, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Deeti, S.; O’Farrell, S.; Kennedy, B.N. Early safety assessment of human oculotoxic drugs using the zebrafish visualmotor response. J. Pharmacol. Toxicol. Methods 2014, 69, 1–8. [Google Scholar] [CrossRef]
- Owens, K.N.; Coffin, A.B.; Hong, L.S.; Bennett, K.O.; Rubel, E.W.; Raible, D.W. Response of mechanosensory hair cells of the zebrafish lateral line to aminoglycosides reveals distinct cell death pathways. Hear. Res. 2009, 253, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pindling, S. Dysbiosis and early mortality in zebrafish larvae exposed to subclinical concentrations of streptomycin. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Shao, W.; Zhong, D.; Jiang, H.; Han, Y.; Yin, Y.; Li, R.; Qian, X.; Chen, D.; Jing, L. A new aminoglycoside etimicin shows low nephrotoxicity and ototoxicity in zebrafish embryos. J. Appl. Toxicol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zheng, Y.; Zhang, J.; Hu, C. Neurobehavioral Effects of Cephalosporins: Assessment of Locomotors Activity, Motor and Sensory Development in Zebrafish. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- He, J.-H.; Guo, S.-Y.; Zhu, F.; Zhu, J.-J.; Chen, Y.-X.; Huang, C.-J.; Gao, J.-M.; Dong, Q.-X.; Xuan, Y.-X.; Li, C.-Q. A zebrafish phenotypic assay for assessing drug-induced hepatotoxicity. J. Pharmacol. Toxicol. Methods 2013, 67, 25–32. [Google Scholar] [CrossRef]
- Zhang, M.-Q.; Chen, B.; Zhang, J.-P.; Chen, N.; Liu, C.-Z.; Hu, C.-Q. Liver toxicity of macrolide antibiotics in zebrafish. Toxicology 2020, 441, 152501. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Liu, X.; Yang, F.; Li, R.; Xiong, Y.; Fu, C.; Li, G.; Liu, S.; Zheng, C. Single and joint toxic effects of four antibiotics on some metabolic pathways of zebrafish (Danio rerio) larvae. Sci. Total. Environ. 2020, 716, 137062. [Google Scholar] [CrossRef]
- Yan, Z.; Lu, G.; Ye, Q.; Liu, J. Long-term effects of antibiotics, norfloxacin, and sulfamethoxazole, in a partial life-cycle study with zebrafish (Danio rerio): Effects on growth, development, and reproduction. Environ. Sci. Pollut. Res. 2016, 23, 18222–18228. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Liu, J.; He, S.; Shen, W.; Wei, C.; Li, K.; Zhang, Y.; Yue, J.; Yang, Z. Effects of norfloxacin exposure on neurodevelopment of zebrafish (Danio rerio) embryos. NeuroToxicology 2019, 72, 85–94. [Google Scholar] [CrossRef]
- Han, Y.; Ma, Y.; Yao, S.; Zhang, J.; Hu, C. In vivo and in silico evaluations of survival and cardiac developmental toxicity of quinolone antibiotics in zebrafish embryos (Danio rerio). Environ. Pollut. 2021, 277, 116779. [Google Scholar] [CrossRef]
- Nadal, A.L.; Peggs, D.; Wiegertjes, G.; Brugman, S. Exposure to Antibiotics Affects Saponin Immersion-Induced Immune Stimulation and Shift in Microbial Composition in Zebrafish Larvae. Front. Microbiol. 2018, 9, 2588. [Google Scholar] [CrossRef]
- Zhao, H.; Quan, W.; Bekele, T.G.; Chen, M.; Zhang, X.; Qu, B. Effect of copper on the accumulation and elimination kinetics of fluoroquinolones in the zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2018, 156, 135–140. [Google Scholar] [CrossRef]
- Shen, R.; Yu, Y.; Lan, R.; Yu, R.; Yuan, Z.; Xia, Z. The cardiovascular toxicity induced by high doses of gatifloxacin and ciprofloxacin in zebrafish. Environ. Pollut. 2019, 254, 112861. [Google Scholar] [CrossRef]
- Tokanová, N.; Dobšíková, R.; Doubková, V.; Blahová, J.; Svobodová, Z.; Maršálek, P. The effect of sulfamethoxazole on oxidative stress indices in zebrafish (Danio rerio). Drug Chem. Toxicol. 2021, 44, 58–63. [Google Scholar] [CrossRef]
- Zhou, L.; Limbu, S.M.; Shen, M.; Zhai, W.; Qiao, F.; He, A.; Du, Z.-Y.; Zhang, M. Environmental concentrations of antibiotics impair zebrafish gut health. Environ. Pollut. 2018, 235, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wei, T.; Wu, X.; Zhong, H.; Qiu, W.; Zheng, Y. Early exposure to environmental levels of sulfamethoxazole triggers immune and inflammatory response of healthy zebrafish larvae. Sci. Total. Environ. 2020, 703, 134724. [Google Scholar] [CrossRef]
- Hamid, N.; Junaid, M.; Pei, D.-S. Individual and combined mechanistic toxicity of sulfonamides and their implications for ecological risk assessment in the Three Gorges Reservoir Area (TGRA), China. J. Hazard. Mater. 2020, 382, 121106. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Yang, Q.; Jiang, W.; Lu, J.; Xiang, Z.; Guo, R.; Chen, J. Integrated toxic evaluation of sulfamethazine on zebrafish: Including two lifespan stages (embryo-larval and adult) and three exposure periods (exposure, post-exposure and re-exposure). Chemosphere 2018, 195, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Li, X.; Qiu, Y.; Zeng, X.; Yu, X.; Wang, W.; Yi, X.; Huang, L. Low-dose effects on thyroid disruption in zebrafish by long-term exposure to oxytetracycline. Aquat. Toxicol. 2020, 227, 105608. [Google Scholar] [CrossRef]
- Yu, X.; Wu, Y.; Deng, M.; Liu, Y.; Wang, S.; He, X.; Allaire-Leung, M.; Wan, J.; Zou, Y.; Yang, C.; et al. Tetracycline antibiotics as PI3K inhibitors in the Nrf2-mediated regulation of antioxidative stress in zebrafish larvae. Chemosphere 2019, 226, 696–703. [Google Scholar] [CrossRef]
- Rosenberg, C.R.; Fang, X.; Allison, K.R. Potentiating aminoglycoside antibiotics to reduce their toxic side effects. PLoS ONE 2020, 15, e0237948. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.B.T.; Rahman, N.; Khan, R.A. A review on antibiotic resistance and way of combating antimicrobial resistance. GSC Biol. Pharm. Sci. 2021, 14, 087–097. [Google Scholar] [CrossRef]
- Eliopoulos, G.M.; Drusano, G.L.; Ambrose, P.G.; Bhavnani, S.M.; Bertino, J.S.; Nafziger, A.N.; Louie, A. Back to the Future: Using Aminoglycosides Again and How to Dose Them Optimally. Clin. Infect. Dis. 2007, 45, 753–760. [Google Scholar] [CrossRef]
- Anita, A.B.R. Spectroscopic analysis of netilmicin by derivatization using quality by design for bulk dosage forms. Global Trends Pharm Sci. 2020, 11, 8705–8712. [Google Scholar]
- Chen, C.; Chen, Y.; Wu, P.; Chen, B. Update on new medicinal applications of gentamicin: Evidence-based review. J. Formos. Med Assoc. 2014, 113, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riaz, L. Chapter2: Antibiotics Use in Hospitals and Their Presence in the Associated Waste. In Antibiotics and Antimicrobial Resistance Genes. Environmental Occurrence and Treatment Technologies; Hashmi, M.Z., Ed.; Springer: Berlin, Germany, 2020; pp. 27–49. [Google Scholar]
- Thakuria, B.; Lahon, K. The Beta Lactam Antibiotics as an Empirical Therapy in a Developing Country: An Update on Their Current Status and Recommendations to Counter the Resistance against Them. J. Clin. Diagn. Res. 2013, 7, 1207–1214. [Google Scholar] [CrossRef]
- Sauberan, J.B.; Bradley, J.S. 292 Antimicrobial agents. In Principles and Practice of Pediatric Infectious Diseases, 5th ed.; Long, S.S., Prober, C.G., Fischer, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1499–1531. [Google Scholar]
- Foster, T.J. Can β-Lactam Antibiotics Be Resurrected to Combat MRSA? Trends Microbiol. 2019, 27, 26–38. [Google Scholar] [CrossRef]
- Pham, T.D.M.; Ziora, Z.M.; Blaskovich, M.A.T. Quinolone antibiotics. MedChemComm 2019, 10, 1719–1739. [Google Scholar] [CrossRef]
- Cheong, M.S.; Seo, K.H.; Chohra, H.; Yoon, Y.E.; Choe, H.; Kantharaj, V.; Lee, Y.B. Influence of Sulfonamide Contamination Derived from Veterinary Antibiotics on Plant Growth and Development. Antibiotics 2020, 9, 456. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Cheng, L.; Zheng, Y.; Xu, J. Sediment and salinity effects on the bioaccumulation of sulfamethoxazole in zebrafish (Danio rerio). Chemosphere 2017, 180, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Bakker, E.P. Ionophore Antibiotics. In Mechanism of Action of Antibacterial Agents; Hahn, F.E., Ed.; Springer: Berlin/Heidelberg, Germany, 1979; pp. 67–97. [Google Scholar]
- Kart, A.; Bilgili, A. Ionophore antibiotics: Toxicity, mode of action and neurotoxic aspect of carboxylic ionophores. J. Anim. Vet. Adv. 2008, 7, 748–751. [Google Scholar]
- Jijie, R.; Solcan, G.; Nicoara, M.; Micu, D.; Strungaru, S.-A. Antagonistic effects in zebrafish (Danio rerio) behavior and oxidative stress induced by toxic metals and deltamethrin acute exposure. Sci. Total. Environ. 2020, 698, 134299. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, H.; Liu, J.; Lin, J.; Zeng, A.; Ai, W.; Wang, X.; Dahlgren, R.A.; Wang, H. Immunotoxicity of β-Diketone Antibiotic Mixtures to Zebrafish (Danio rerio) by Transcriptome Analysis. PLoS ONE 2016, 11, e0152530. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, J.; Zhang, Y.; Wang, X.; Li, W.; Zhang, H.; Wang, H.; Zhang, Y. Screening on the differentially expressed miRNAs in zebrafish ( Danio rerio ) exposed to trace β-diketone antibiotics and their related functions. Aquat. Toxicol. 2016, 178, 27–38. [Google Scholar] [CrossRef]
- Wang, X.; Ma, Y.; Liu, J.; Yin, X.; Zhang, Z.; Wang, C.; Li, Y.; Wang, H. Reproductive toxicity of β-diketone antibiotic mixtures to zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2017, 141, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, J.; Li, F.; Zhang, C.; Li, J.; Wang, C.; Dahlgren, R.A.; Zhang, H.; Wang, H. Screening and functional identification of lncRNAs under β-diketone antibiotic exposure to zebrafish (Danio rerio) using high-throughput sequencing. Aquat. Toxicol. 2017, 182, 214–225. [Google Scholar] [CrossRef]
- Qiu, W.; Fang, M.; Magnuson, J.T.; Greer, J.B.; Chen, Q.; Zheng, Y.; Xiong, Y.; Luo, S.; Zheng, C.; Schlenk, D. Maternal exposure to environmental antibiotic mixture during gravid period predicts gastrointestinal effects in zebrafish offspring. J. Hazard. Mater. 2020, 399, 123009. [Google Scholar] [CrossRef]
- Wang, H.; Che, B.; Duan, A.; Mao, J.; Dahlgren, R.A.; Zhang, M.; Zhang, H.; Zeng, A.; Wang, X. Toxicity evaluation of β-diketone antibiotics on the development of embryo-larval zebrafish (Danio rerio). Environ. Toxicol. 2014, 29, 1134–1146. [Google Scholar] [CrossRef] [PubMed]
- Bielen, A.; Simatovic, A.; Kosić-Vukšić, J.; Senta, I.; Ahel, M.; Babić, S.; Jurina, T.; Plaza, J.J.G.; Milaković, M.; Udiković-Kolić, N. Negative environmental impacts of antibiotic-contaminated effluents from pharmaceutical industries. Water Res. 2017, 126, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Maselli, K.M.; Gee, K.; Isani, M.; Fode, A.; Schall, K.A.; Grikscheit, T.C. Broad-spectrum antibiotics alter the microbiome, increase intestinal fxr, and decrease hepatic steatosis in zebrafish short bowel syndrome. Am. J. Physiol. Liver Physiol. 2020, 319, G212–G226. [Google Scholar] [CrossRef] [PubMed]



| Changes in F0 Female (n = 20 Individuals) | Antibiotic Concentrations | ||
|---|---|---|---|
| Control | 1 μg/L | 100 μg/L | |
| Body weight (g) | 0.51 ± 0.016 | 0.53 ± 0.015 | 0.56 ± 0.025 |
| Body length (cm) | 3.58 ± 0.029 | 3.55 ± 0.032 | 3.66 ± 0.031* |
| Intestinal weight (g) | 0.02 ± 0.001 | 0.025 ± 0.002 | 0.024 ± 0.001 |
| Ovary weight (g) | 0.07 ±0.004 | 0.084 ± 0.005 | 0.087 ± 0.004 |
| Changes in F1 embryo and larval | Control | 1 μg/L | 100 μg/L |
| Egg production (number per parent, 20 individuals) | 490.6 ± 23.09 | 442 ± 134.51 | 397.3 ± 31.39 |
| Egg death rate at birth (% 0 hpf, 3 biological replicates) | 5.09 ± 2.05 | 6.11 ± 3.05 | 17.76 ± 3.3 * |
| Fertilization rate (% 4 hpf, 3 biological replicates) | 73.2 ± 0.73 | 78.6 ± 1.77 | 74.7 ± 2.25 |
| Egg death rate at 120 hpf (%, 3 biological replicates) | 1.7 ± 0.13 | 2 ± 0.26 | 5.9 ± 0.94 * |
| Hatching rate (%, 72 hpf, 3 biological replicates) | 94.5 ± 2.94 | 92.1 ± 6 | 89.4 ± 3.29 |
| F1 body length (mm, 120 hpf, 20 individuals) | 3.91 ± 0.02 | 3.88 ± 0.02 | 3.92 ± 0.02 |
| Displacement distance (mm, 0–10 min, 120 hpf, 20 individuals) | 582.7 ± 106.2 | 678.4 ± 109 | 465.4 ± 58.6 |
| Antibiotic | Concentration (μg/L) | ||
|---|---|---|---|
| Winter | Spring | ||
| 1 | Azithromycin | 2137 | 3776 |
| n-desmethyl azithromycin | 2341 | 5660 | |
| Erythromycin | 2009 | 1069 | |
| Summer | Autumn | ||
| 2 | Sulfadiazine | 7.1 | 3 |
| Sulfamethazine | 231 | 6.7 | |
| Trimethoprim | 5.4 | 1 | |
| Enrofloxacin | 4.3 | 3.6 | |
| Oxytetracycline | 29 | 10 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jijie, R.; Mihalache, G.; Balmus, I.-M.; Strungaru, S.-A.; Baltag, E.S.; Ciobica, A.; Nicoara, M.; Faggio, C. Zebrafish as a Screening Model to Study the Single and Joint Effects of Antibiotics . Pharmaceuticals 2021, 14, 578. https://doi.org/10.3390/ph14060578
Jijie R, Mihalache G, Balmus I-M, Strungaru S-A, Baltag ES, Ciobica A, Nicoara M, Faggio C. Zebrafish as a Screening Model to Study the Single and Joint Effects of Antibiotics . Pharmaceuticals. 2021; 14(6):578. https://doi.org/10.3390/ph14060578
Chicago/Turabian StyleJijie, Roxana, Gabriela Mihalache, Ioana-Miruna Balmus, Stefan-Adrian Strungaru, Emanuel Stefan Baltag, Alin Ciobica, Mircea Nicoara, and Caterina Faggio. 2021. "Zebrafish as a Screening Model to Study the Single and Joint Effects of Antibiotics " Pharmaceuticals 14, no. 6: 578. https://doi.org/10.3390/ph14060578
APA StyleJijie, R., Mihalache, G., Balmus, I.-M., Strungaru, S.-A., Baltag, E. S., Ciobica, A., Nicoara, M., & Faggio, C. (2021). Zebrafish as a Screening Model to Study the Single and Joint Effects of Antibiotics . Pharmaceuticals, 14(6), 578. https://doi.org/10.3390/ph14060578