
Protective Effects of Irbesartan, an Angiotensin Receptor Blocker with PPARγ Agonistic Activity, against Estradiol Benzoate-Induced Endometrial Hyperplasia and Atypia in Female Rats via Modulation of TNFα/Survivin Pathway



Abstract
:1. Introduction
2. Results
2.1. Effect of IRB on Uterine Weight, Malondialdehyde (MDA), and SOD
2.2. Histopathological and Immunohistochemical Evaluation
2.2.1. Macroscopic Examination
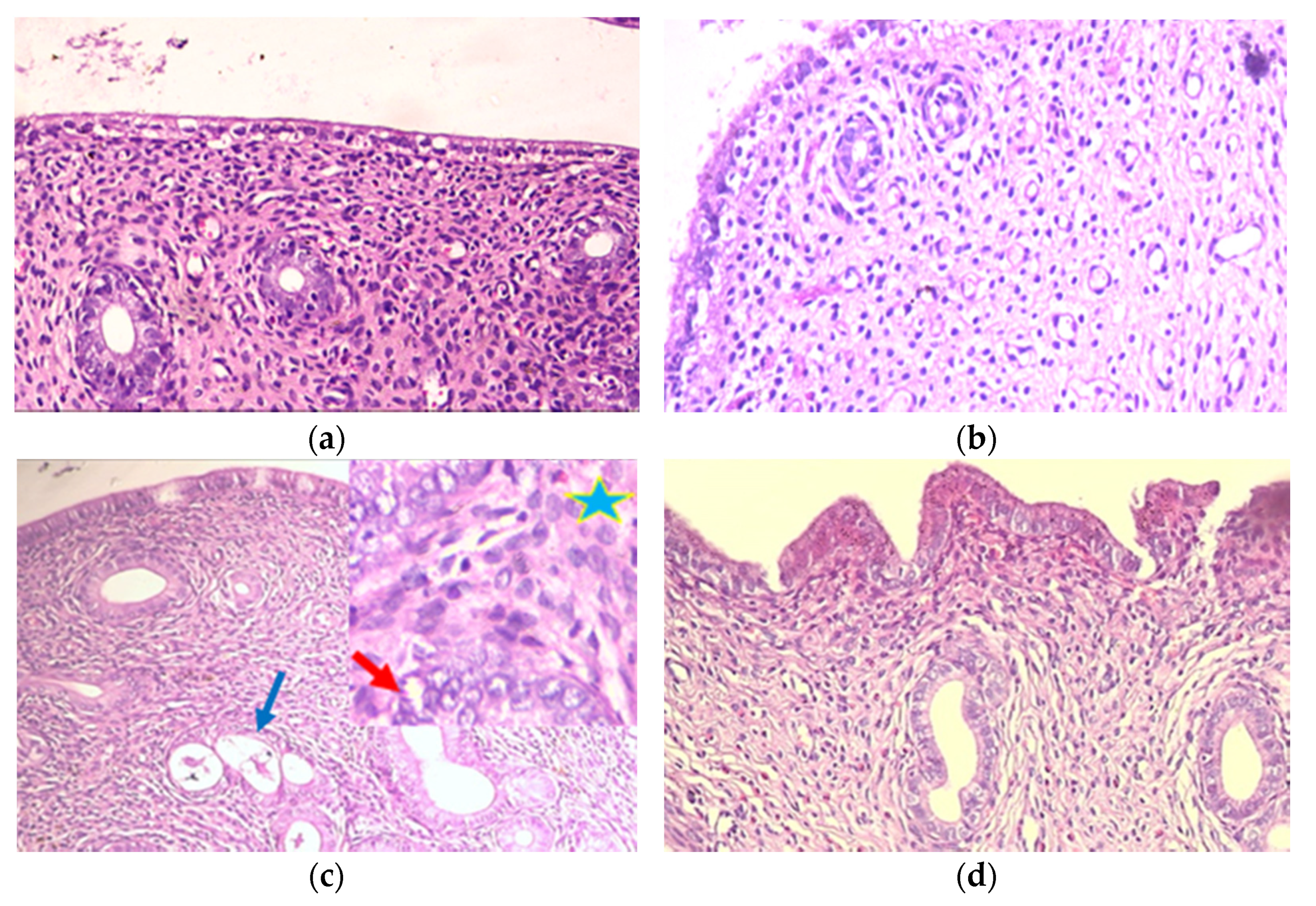
2.2.2. Histopathological Evaluation
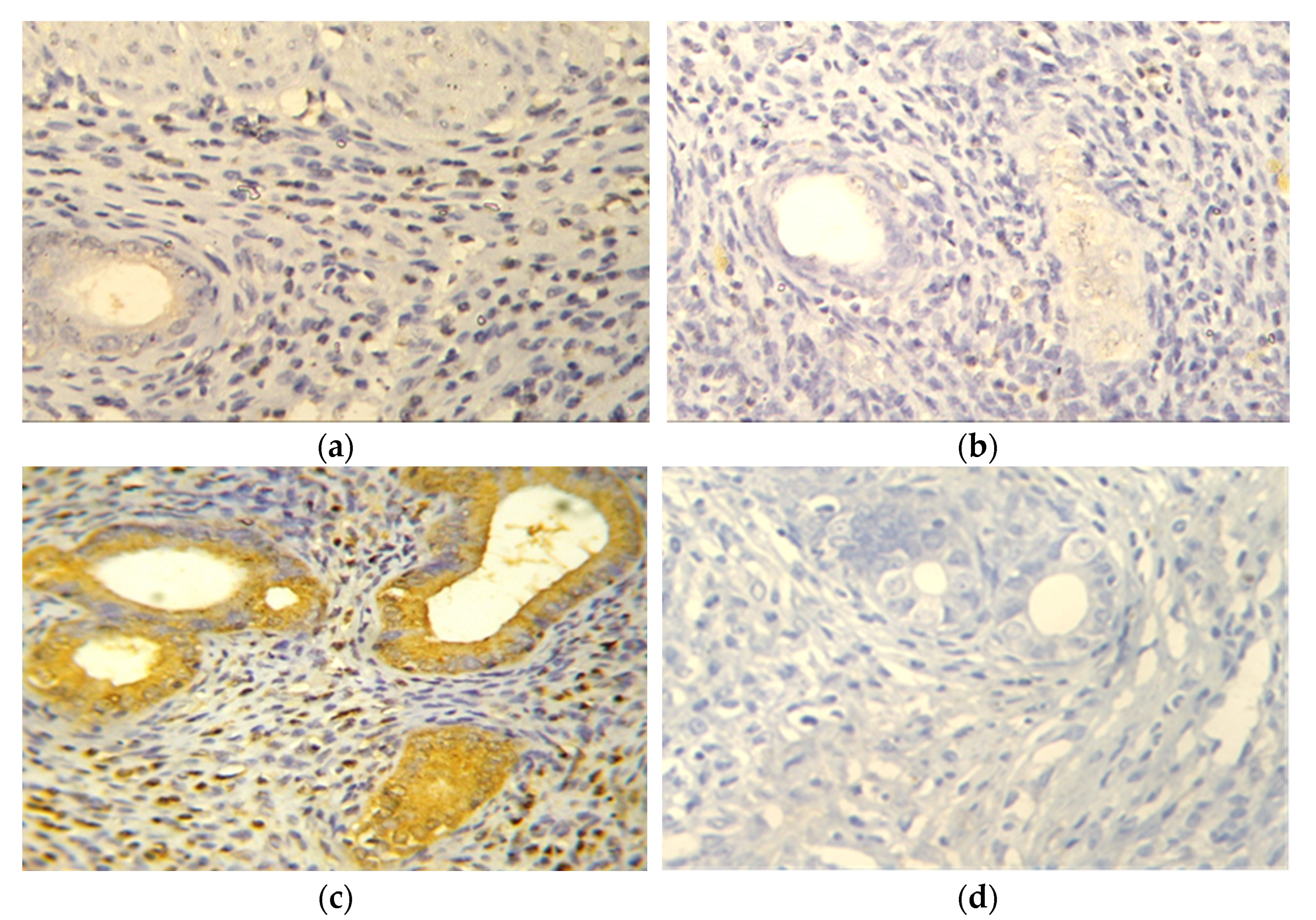
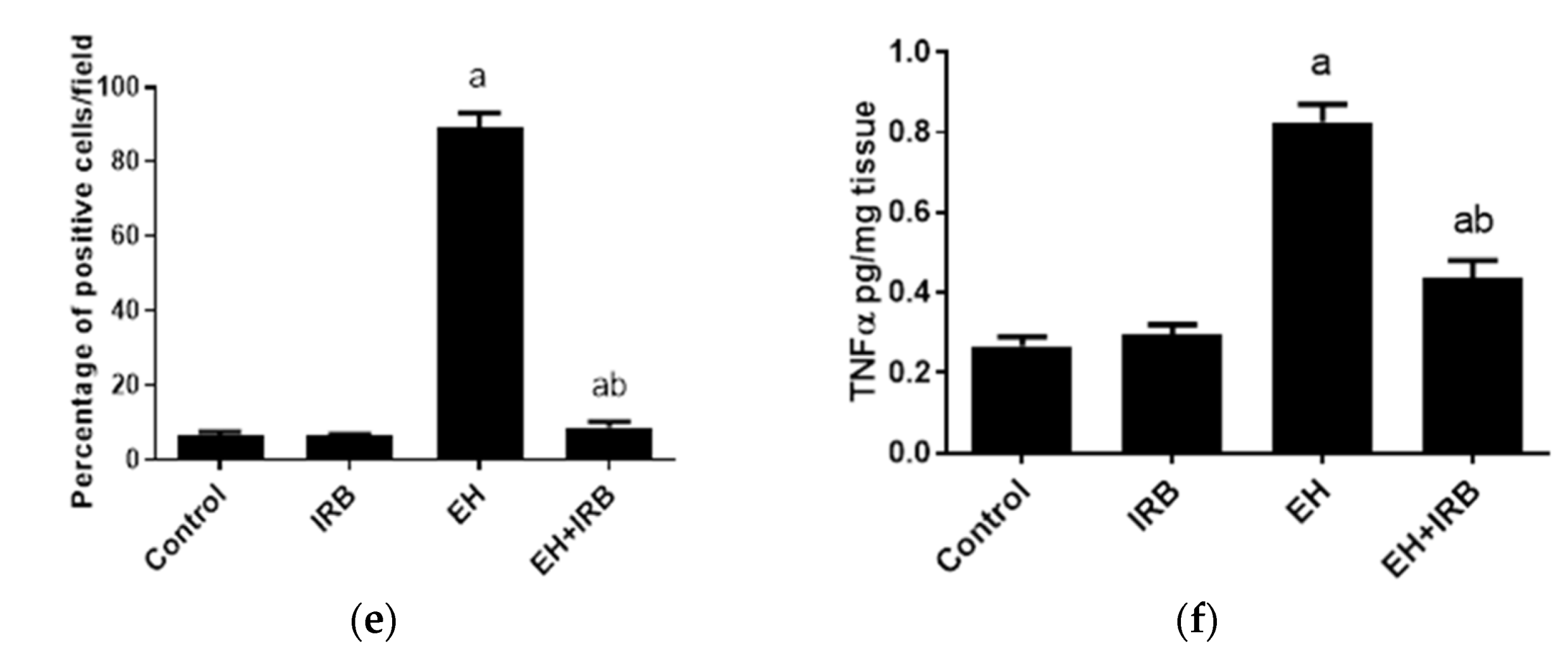
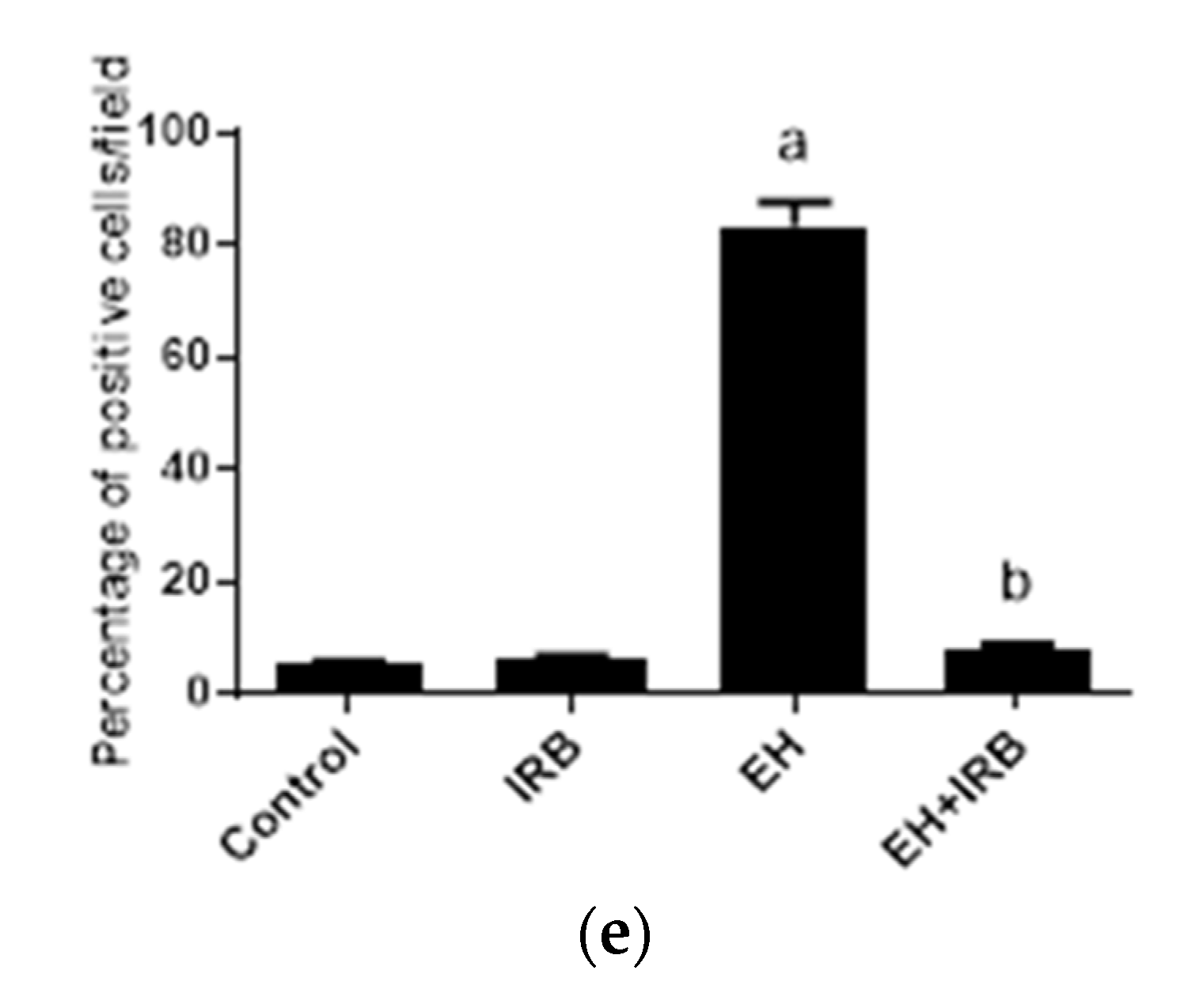
2.2.3. Evaluation of TNFα (Immunohistochemistry and ELISA)
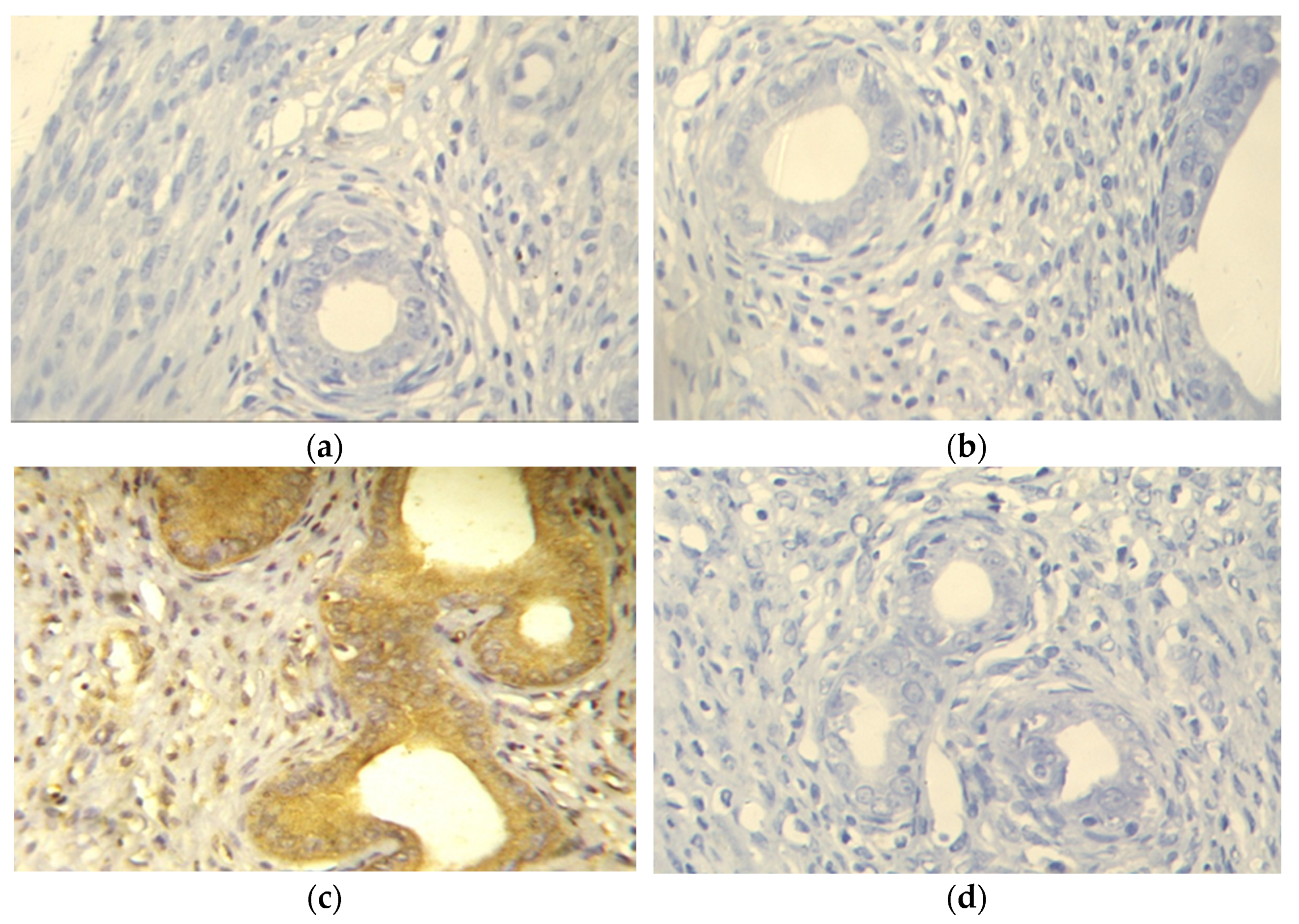
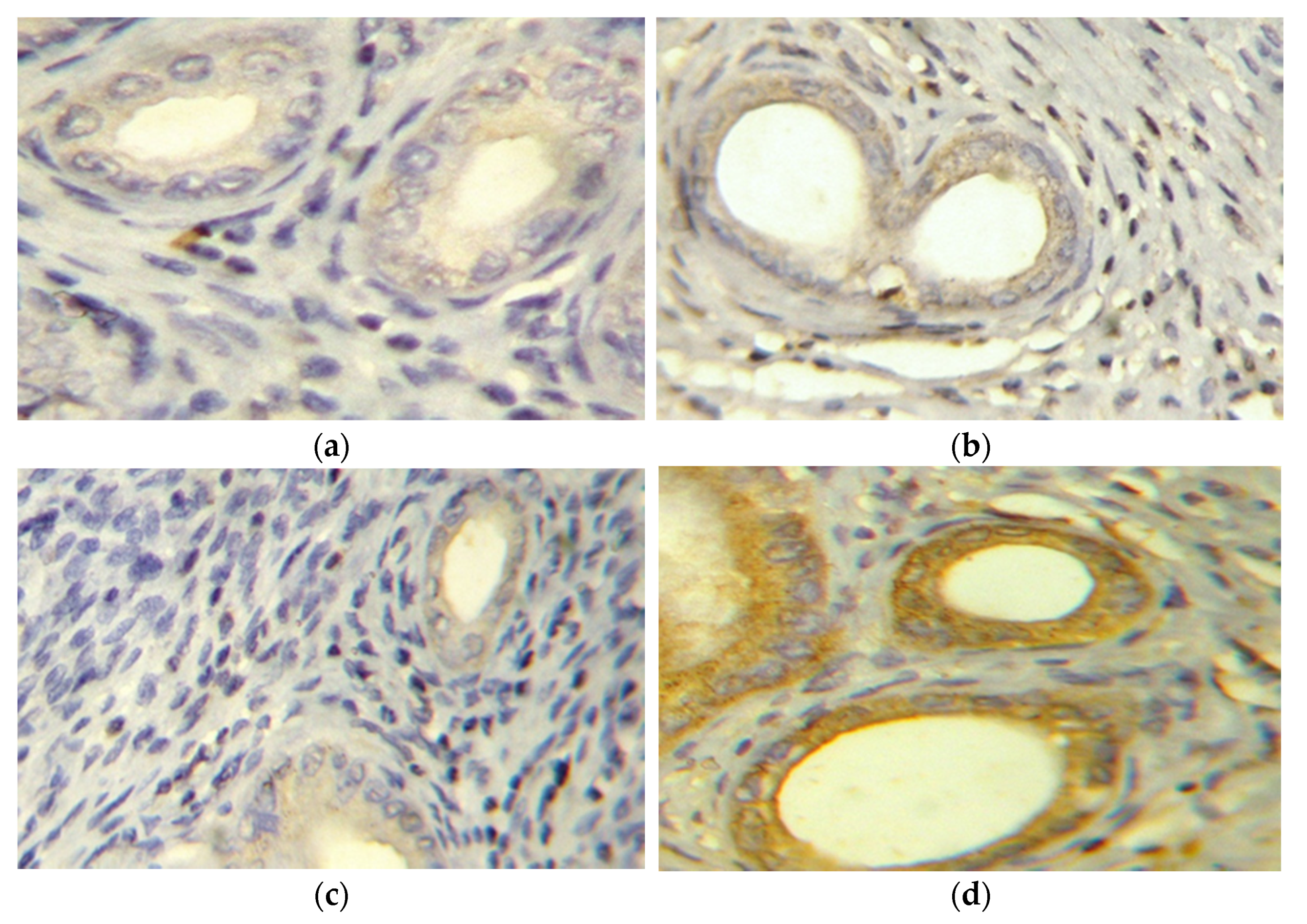
2.2.4. Evaluation of Survivin Expression
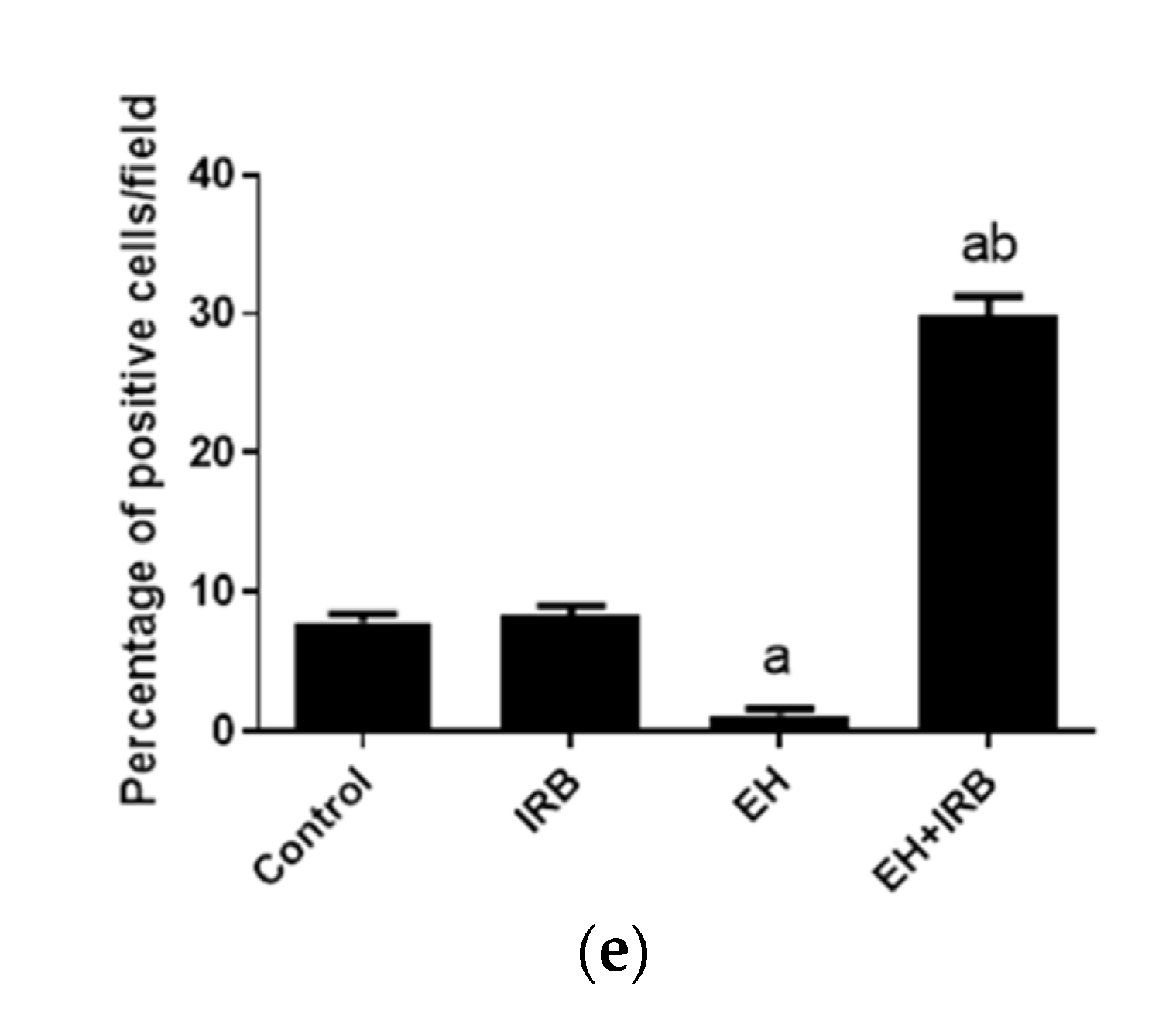
2.2.5. Evaluation of Cleaved Caspase 3 Expression
2.3. Effect of IRB on IL10 Gene Expression
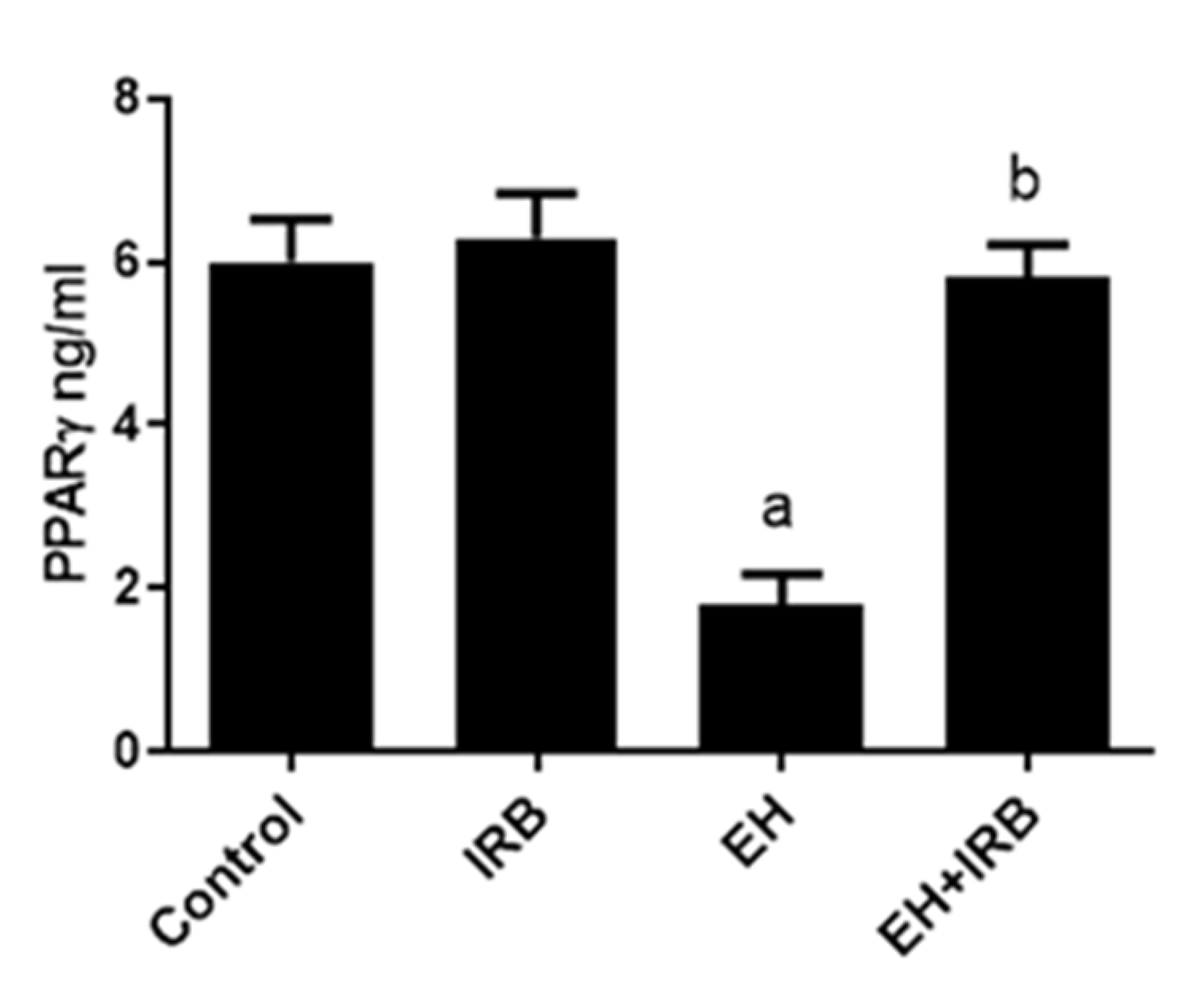
2.4. Effect of IRB on PPARγ Level
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Experimental Design
4.3. Preparation of Uterine Homogenate
4.4. Biochemical Analysis
4.4.1. Determination of Uterine MDA Level
4.4.2. Determination of Uterine SOD Activity
4.5. Macroscopic Examination and Histopathological Evaluation
4.6. Immunohistochemistry
4.7. Real-Time PCR
4.8. ELISA
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sengal, A.T.; Smith, D.; Rogers, R.; Snell, C.E.; Williams, E.D.; Pollock, P.M. Fibroblast Growth Factor Receptor 2 Isoforms Detected via Novel RNA ISH as Predictive Biomarkers for Progestin Therapy in Atypical Hyperplasia and Low-Grade Endometrial Cancer. Cancers 2021, 13, 1703. [Google Scholar] [CrossRef]
- Sobczuk, K.; Sobczuk, A. New classification system of endometrial hyperplasia WHO 2014 and its clinical implications. Prz. Menopauzalny 2017, 16, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Genome Atlas Research Network; Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; et al. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar]
- Modugno, F.; Ness, R.B.; Chen, C.; Weiss, N.S. Inflammation and endometrial cancer: A hypothesis. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2840–2847. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.O.; Stephens, N.D.; Qualls, C.R.; Fligelman, T.; Wang, T.; Lin, C.Y.; Burton, E.; Griffith, J.K.; Pollard, J.W. The clinical significance of inflammatory cytokines in primary cell culture in endometrial carcinoma. Mol. Oncol. 2013, 7, 41–54. [Google Scholar] [CrossRef]
- Jabbour, H.N.; Sales, K.J.; Catalano, R.D.; Norman, J.E. Inflammatory pathways in female reproductive health and disease. Reproduction 2009, 138, 903–919. [Google Scholar] [CrossRef] [PubMed]
- Portt, L.; Norman, G.; Clapp, C.; Greenwood, M.; Greenwood, M.T. Anti-apoptosis and cell survival: A review. Biochim. Biophys. Acta 2011, 1813, 238–259. [Google Scholar] [CrossRef] [Green Version]
- Landskron, G.; De la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic inflammation and cytokines in the tumor microenvironment. J. Immunol. Res. 2014, 2014, 149185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altieri, D.C. Survivin and IAP proteins in cell-death mechanisms. Biochem. J. 2010, 430, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Sharma, A.K.; Dhingra, S.; Singal, P.K. Interplay of TNF-alpha and IL-10 in regulating oxidative stress in isolated adult cardiac myocytes. J. Mol. Cell. Cardiol. 2006, 41, 1023–1030. [Google Scholar] [CrossRef]
- Li, X.F.; Ahmed, A. Expression of angiotensin II and its receptor subtypes in endometrial hyperplasia: A possible role in dysfunctional menstruation. Lab. Investig. 1996, 75, 137–145. [Google Scholar] [PubMed]
- Delforce, S.J.; Lumbers, E.R.; Corbisier de Meaultsart, C.; Wang, Y.; Proietto, A.; Otton, G.; Scurry, J.; Verrills, N.M.; Scott, R.J.; Pringle, K.G. Expression of renin–angiotensin system (RAS) components in endometrial cancer. Endocr. Connect. 2017, 6, 9–19. [Google Scholar] [CrossRef]
- Piastowska-Ciesielska, A.W.; Płuciennik, E.; Wójcik-Krowiranda, K.; Bieńkiewicz, A.; Bednarek, A.; Ochędalski, T. Analysis of the expression of angiotensin II type 1 receptor and VEGF in endometrial adenocarcinoma with different clinicopathological characteristics. Tumour Biol. 2012, 33, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Shang, W.; Zhang, L.; Zhao, C.; Chen, W.; Zhang, Y.; Li, G. Effect of angiotensin-(1-7) and angiotensin II on the proliferation and activation of human endometrial stromal cells in vitro. Int. J. Clin. Exp. Pathol. 2015, 8, 8948–8957. [Google Scholar]
- Matysiak-Burzyńska, Z.E.; Nowakowska, M.; Domińska, K.; Kowalska, K.; Płuciennik, E.; Piastowska-Ciesielska, A.W. Silencing of angiotensin receptor 1 interferes with angiotensin II oncogenic activity in endometrial cancer. J. Cell. Biochem. 2018, 119, 9110–9121. [Google Scholar] [CrossRef] [PubMed]
- Ziaja, M.; Urbanek, K.A.; Kowalska, K.; Piastowska-Ciesielska, A.W. Angiotensin II and Angiotensin Receptors 1 and 2-Multifunctional System in Cells Biology, What Do We Know? Cells 2021, 10, 381. [Google Scholar] [CrossRef] [PubMed]
- Koyama, N.; Nishida, Y.; Ishii, T.; Yoshida, T.; Furukawa, Y.; Narahara, H. Telmisartan induces growth inhibition, DNA double-strand breaks and apoptosis in human endometrial cancer cells. PLoS ONE 2014, 9, e93050. [Google Scholar] [CrossRef]
- Kinoshita, J.; Fushida, S.; Harada, S.; Yagi, Y.; Fujita, H.; Kinami, S.; Ninomiya, I.; Fujimura, T.; Kayahara, M.; Yashiro, M.; et al. Local angiotensin II-generation in human gastric cancer: Correlation with tumor progression through the activation of ERK1/2, NF-kappaB and survivin. Int. J. Oncol. 2009, 34, 1573–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Ning, W.; Xie, D.; Guo, L.; DuBois, R.N. Peroxisome proliferator-activated receptor delta confers resistance to peroxisome proliferator-activated receptor gamma-induced apoptosis in colorectal cancer cells. Oncogene 2012, 31, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Aljahdali, I.; Ling, X. Cancer therapeutics using survivin BIRC5 as a target: What can we do after over two decades of study? J. Exp. Clin. Cancer Res. 2019, 38, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, E.A.; Pitts, T.P.; Tandberg, K.R.; Winder, P.L.; Wright, A.E. Discovery of Survivin Inhibitors Part 1: Screening the Harbor Branch Pure Compound Library. Mar. Drugs 2021, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Ota, K.; Ito, K.; Suzuki, T.; Saito, S.; Tamura, M.; Hayashi, S.; Okamura, K.; Sasano, H.; Yaegashi, N. Peroxisome proliferator-activated receptor gamma and growth inhibition by its ligands in uterine endometrial carcinoma. Clin. Cancer Res. 2006, 12, 4200–4208. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Gong, W.; Lu, L.; Chen, J.; Lu, Z.; Li, H.; Liu, W.; Liu, Y.; Wang, M.; Hu, R.; et al. Irbesartan ameliorates hyperlipidemia and liver steatosis in type 2 diabetic db/db mice via stimulating PPAR-gamma, AMPK/Akt/mTOR signaling and autophagy. Int. Immunopharmacol. 2017, 42, 176–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Al-Nami, M.S. Irbesartan Attenuates Gentamicin-induced Nephrotoxicity in Rats through Modulation of Oxidative Stress and Endogenous Antioxidant Capacity. Int. J. Prev. Med. 2020, 11, 16. [Google Scholar] [PubMed]
- Helal, M.G.; Samra, Y.A. Irbesartan mitigates acute liver injury, oxidative stress, and apoptosis induced by acetaminophen in mice. J. Biochem. Mol. Toxicol. 2020, 34, e22447. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, L.; Ding, Y. Effects of irbesartan on phenotypic alterations in monocytes and the inflammatory status of hypertensive patients with left ventricular hypertrophy. BMC Cardiovasc. Disord. 2021, 21, 194. [Google Scholar] [CrossRef] [PubMed]
- Pejić, S.; Todorović, A.; Stojiljković, V.; Kasapović, J.; Pajović, S.B. Antioxidant enzymes and lipid peroxidation in endometrium of patients with polyps, myoma, hyperplasia and adenocarcinoma. Reprod. Biol. Endocrinol. 2009, 7, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Gupta, S.; So, J.Y.; Wall, B.; Wahler, J.; Smolarek, A.K.; Sae-Tan, S.; Soewono, K.Y.; Yu, H.; Lee, M.J.; Thomas, P.E.; et al. Tocopherols inhibit oxidative and nitrosative stress in estrogen-induced early mammary hyperplasia in ACI rats. Mol. Carcinog. 2015, 54, 916–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refaie, M.M.M.; El-Hussieny, M. The role of interleukin-1b and its antagonist (diacerein) in estradiol benzoate-induced endometrial hyperplasia and atypia in female rats. Fundam. Clin. Pharmacol. 2017, 31, 438–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olowofolahan, A.O.; Oyebode, O.T.; Olorunsogo, O.O. Methyl palmitate reversed estradiol benzoate-induced endometrial hyperplasia in female rats. Toxicol. Mech. Methods 2021, 31, 43–52. [Google Scholar] [CrossRef]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. The Copper-zinc superoxide dismutase activity in selected diseases. Eur. J. Clin. Investig. 2019, 49, e13036. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Kim, Y.G.; Jeong, K.H.; Lee, S.H.; Lee, T.W.; Ihm, C.G.; Moon, J.Y. Angiotensin II-induced mitochondrial Nox4 is a major endogenous source of oxidative stress in kidney tubular cells. PLoS ONE 2012, 7, e39739. [Google Scholar] [CrossRef] [Green Version]
- Xing, L.; Fang, J.; Zhu, B.; Wang, L.; Chen, J.; Wang, Y.; Huang, J.; Wang, H.; Yao, X. Astragaloside IV protects against podocyte apoptosis by inhibiting oxidative stress via activating PPARgamma-Klotho-FoxO1 axis in diabetic nephropathy. Life Sci. 2021, 269, 119068. [Google Scholar] [CrossRef]
- Kubyshkin, A.V.; Aliev, L.L.; Fomochkina, I.I.; Kovalenko, Y.P.; Litvinova, S.V.; Filonenko, T.G.; Lomakin, N.V.; Kubyshkin, V.A.; Karapetian, O.V. Endometrial hyperplasia-related inflammation: Its role in the development and progression of endometrial hyperplasia. Inflamm. Res. 2016, 65, 785–794. [Google Scholar] [CrossRef]
- Mariappan, N.; Soorappan, R.N.; Haque, M.; Sriramula, S.; Francis, J. TNF-alpha-induced mitochondrial oxidative stress and cardiac dysfunction: Restoration by superoxide dismutase mimetic Tempol. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2726–H2737. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Ono, M.; Shono, T.; Izumi, H.; Ishibashi, T.; Suzuki, H.; Kuwano, M. Involvement of interleukin-8, vascular endothelial growth factor, and basic fibroblast growth factor in tumor necrosis factor alpha-dependent angiogenesis. Mol. Cell. Biol. 1997, 17, 4015–4023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purohit, A.; Newman, S.P.; Reed, M.J. The role of cytokines in regulating estrogen synthesis: Implications for the etiology of breast cancer. Breast Cancer Res. 2002, 4, 65–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelzaher, W.Y.; Bahaa, H.A.; Toni, N.D.M.; Sanad, A.S. Mechanisms underlying the protective effect of montelukast in prevention of endometrial hyperplasia in female rats. Int. Immunopharmacol. 2018, 62, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, G.T.; Zhdanov, A.V.; Davydova, M.P.; Slukina, T.V.; Chernukha, G.E.; Samoilova, T.E.; Smetnik, V.P. Disorders in cytokine gene expression in endometrial hyperplasia and effect of hormone therapy. Bull. Exp. Biol. Med. 2005, 139, 235–237. [Google Scholar] [CrossRef]
- Abdel-Raheem, I.T.; Omran, G.A.; Katary, M.A. Irbesartan, an angiotensin II receptor antagonist, with selective PPAR-gamma-modulating activity improves function and structure of chemotherapy-damaged ovaries in rats. Fundam. Clin. Pharmacol. 2015, 29, 286–298. [Google Scholar] [CrossRef]
- Huang, G.; Cheng, P.; Ding, L.; Wang, L.; Hu, J.; Zhang, Y.; Cai, G.; Chen, M.; Shen, A.; Gao, S. Protective effect of Xin-Ji-Er-Kang on cardiovascular remodeling in high salt-induced hypertensive mice. Exp. Ther. Med. 2019, 17, 1551–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakshmi, S.P.; Reddy, A.T.; Banno, A.; Reddy, R.C. Airway Epithelial Cell Peroxisome Proliferator-Activated Receptor γ Regulates Inflammation and Mucin Expression in Allergic Airway Disease. J. Immunol. 2018, 201, 1775–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erkanli, S.; Kayaselcuk, F.; Kuscu, E.; Bagis, T.; Bolat, F.; Haberal, A.; Demirhan, B. Expression of survivin, PTEN and p27 in normal, hyperplastic, and carcinomatous endometrium. Int. J. Gynecol. Cancer 2006, 16, 1412–1418. [Google Scholar] [CrossRef]
- He, F.; Zhang, W.; Zhang, H. Apoptotic signaling pathways in uteri of rats with endometrial hyperplasia induced by ovariectomy combined with estrogen. Gynecol. Obstet. Invest. 2013, 76, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, H.; Takagi, H.; Oh, H.; Suzuma, K.; Suzuma, I.; Miyamoto, N.; Uemura, A.; Watanabe, D.; Murakami, T.; Sugaya, T.; et al. Phosphatidylinositol 3-kinase/Akt regulates angiotensin II-induced inhibition of apoptosis in microvascular endothelial cells by governing survivin expression and suppression of caspase-3 activity. Circ. Res. 2004, 94, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Amaya, K.; Ohta, T.; Kitagawa, H.; Kayahara, M.; Takamura, H.; Fujimura, T.; Nishimura, G.; Shimizu, K.; Miwa, K. Angiotensin II activates MAP kinase and NF-kappaB through angiotensin II type I receptor in human pancreatic cancer cells. Int. J. Oncol. 2004, 25, 849–856. [Google Scholar]
- Bafford, R.; Sui, X.X.; Wang, G.; Conte, M. Angiotensin II and tumor necrosis factor-alpha upregulate survivin and Kruppel-like factor 5 in smooth muscle cells: Potential relevance to vein graft hyperplasia. Surgery 2006, 140, 289–296. [Google Scholar] [CrossRef]
- Li, Z.; Yan, Z.; Xu, C.; Dong, Y.; Xiong, Y.; Dai, Y. Acetylshikonin attenuates angiotensin II-induced proliferation and motility of human brain smooth muscle cells by inhibiting Wnt/beta-catenin signaling. Hum. Cell 2018, 31, 242–250. [Google Scholar] [CrossRef]
- Abdel-Hamid, H.A.; Zenhom, N.M.; Toni, N.D. Melatonin reduced endometrial hyperplasia induced by estradiol in female albino rats. Gen. Physiol. Biophys. 2019, 38, 63–71. [Google Scholar] [CrossRef]
- Ager, E.I.; Wen, S.W.; Chan, J.; Chong, W.W.; Neo, J.H.; Christophi, C. Altered efficacy of AT1R-targeted treatment after spontaneous cancer cell-AT1R upregulation. BMC Cancer 2011, 11, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clasen, R.; Schupp, M.; Foryst-Ludwig, A.; Sprang, C.; Clemenz, M.; Krikov, M.; Thöne-Reineke, C.; Unger, T.; Kintscher, U. PPARgamma-activating angiotensin type-1 receptor blockers induce adiponectin. Hypertension 2005, 46, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Michalczyk, K.; Niklas, N.; Rychlicka, M.; Cymbaluk-Płoska, A. The Influence of Biologically Active Substances Secreted by the Adipose Tissue on Endometrial Cancer. Diagnostics 2021, 11, 494. [Google Scholar] [CrossRef] [PubMed]
- Afzal, S.; Sattar, M.A.; Johns, E.J.; Abdulla, M.H.; Akhtar, S.; Hashmi, F.; Abdullah, N.A. Interaction between irbesartan, peroxisome proliferator-activated receptor (PPAR-γ), and adiponectin in the regulation of blood pressure and renal function in spontaneously hypertensive rats. J. Physiol. Biochem. 2016, 72, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- de Matos, F.R.; de Moraes, M.; Nonaka, C.F.; de Souza, L.B.; de Almeida Freitas, R. Immunoexpression of TNF-α and TGF-β in central and peripheral giant cell lesions of the jaws. J. Oral Pathol. Med. 2012, 41, 194–199. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Meng, X.; Du, Z.X.; Fang, C.Q.; Liu, G.L.; Wang, H.Q.; Deng, W.W. Significance of survivin, caspase-3, and VEGF expression in thyroid carcinoma. Clin. Exp. Med. 2009, 9, 207–213. [Google Scholar] [CrossRef]
- Gonzalez, P.; Burgaya, F.; Acarin, L.; Peluffo, H.; Castellano, B.; Gonzalez, B. Interleukin-10 and interleukin-10 receptor-I are upregulated in glial cells after an excitotoxic injury to the postnatal rat brain. J. Neuropathol. Exp. Neurol. 2009, 68, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Strøm, C.C.; Aplin, M.; Ploug, T.; Christoffersen, T.E.; Langfort, J.; Viese, M.; Galbo, H.; Haunsø, S.; Sheikh, S.P. Expression profiling reveals differences in metabolic gene expression between exercise-induced cardiac effects and maladaptive cardiac hypertrophy. FEBS J. 2005, 272, 2684–2695. [Google Scholar] [CrossRef]
- VanGuilder, H.D.; Vrana, K.E.; Freeman, W.M. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques 2008, 44, 619–626. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Uterine Weight (g) | MDA (nmol/g Tissue) | SOD (U/g Tissue) |
|---|---|---|---|
| Control | 1.0 ± 0.05 | 27.0 ± 2.7 | 3701 ± 374 |
| IRB | 0.9 ± 0.05 | 26.7 ± 2.4 | 4050 ± 275 |
| EH | 5.3 ± 0.20 a | 87.2 ± 5.2 a | 1440 ± 150 a |
| EH + IRB | 2.0 ± 0.06 ab | 33.7 ± 2.1 b | 2463 ± 238 ab |
| Group | Glandular Irregularity and Crowdedness | Epithelial Hypertrophy and Stratification | Focal Atypical Cellular Changes | Leukocytic Cell Infiltration |
|---|---|---|---|---|
| Control | - | - | - | + |
| IRB | - | - | - | + |
| EH | +++ | +++ | +++ | +++ |
| EH + IRB | + | + | - | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morsy, M.A.; Abdelraheem, W.M.; El-Hussieny, M.; Refaie, M.M.M. Protective Effects of Irbesartan, an Angiotensin Receptor Blocker with PPARγ Agonistic Activity, against Estradiol Benzoate-Induced Endometrial Hyperplasia and Atypia in Female Rats via Modulation of TNFα/Survivin Pathway. Pharmaceuticals 2021, 14, 649. https://doi.org/10.3390/ph14070649
Morsy MA, Abdelraheem WM, El-Hussieny M, Refaie MMM. Protective Effects of Irbesartan, an Angiotensin Receptor Blocker with PPARγ Agonistic Activity, against Estradiol Benzoate-Induced Endometrial Hyperplasia and Atypia in Female Rats via Modulation of TNFα/Survivin Pathway. Pharmaceuticals. 2021; 14(7):649. https://doi.org/10.3390/ph14070649
Chicago/Turabian StyleMorsy, Mohamed A., Wedad M. Abdelraheem, Maram El-Hussieny, and Marwa M. M. Refaie. 2021. "Protective Effects of Irbesartan, an Angiotensin Receptor Blocker with PPARγ Agonistic Activity, against Estradiol Benzoate-Induced Endometrial Hyperplasia and Atypia in Female Rats via Modulation of TNFα/Survivin Pathway" Pharmaceuticals 14, no. 7: 649. https://doi.org/10.3390/ph14070649
APA StyleMorsy, M. A., Abdelraheem, W. M., El-Hussieny, M., & Refaie, M. M. M. (2021). Protective Effects of Irbesartan, an Angiotensin Receptor Blocker with PPARγ Agonistic Activity, against Estradiol Benzoate-Induced Endometrial Hyperplasia and Atypia in Female Rats via Modulation of TNFα/Survivin Pathway. Pharmaceuticals, 14(7), 649. https://doi.org/10.3390/ph14070649