
Angiotensin-Converting Enzyme 2 (ACE2) in the Context of Respiratory Diseases and Its Importance in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection

 ,
,  ,
,  ,
,  , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
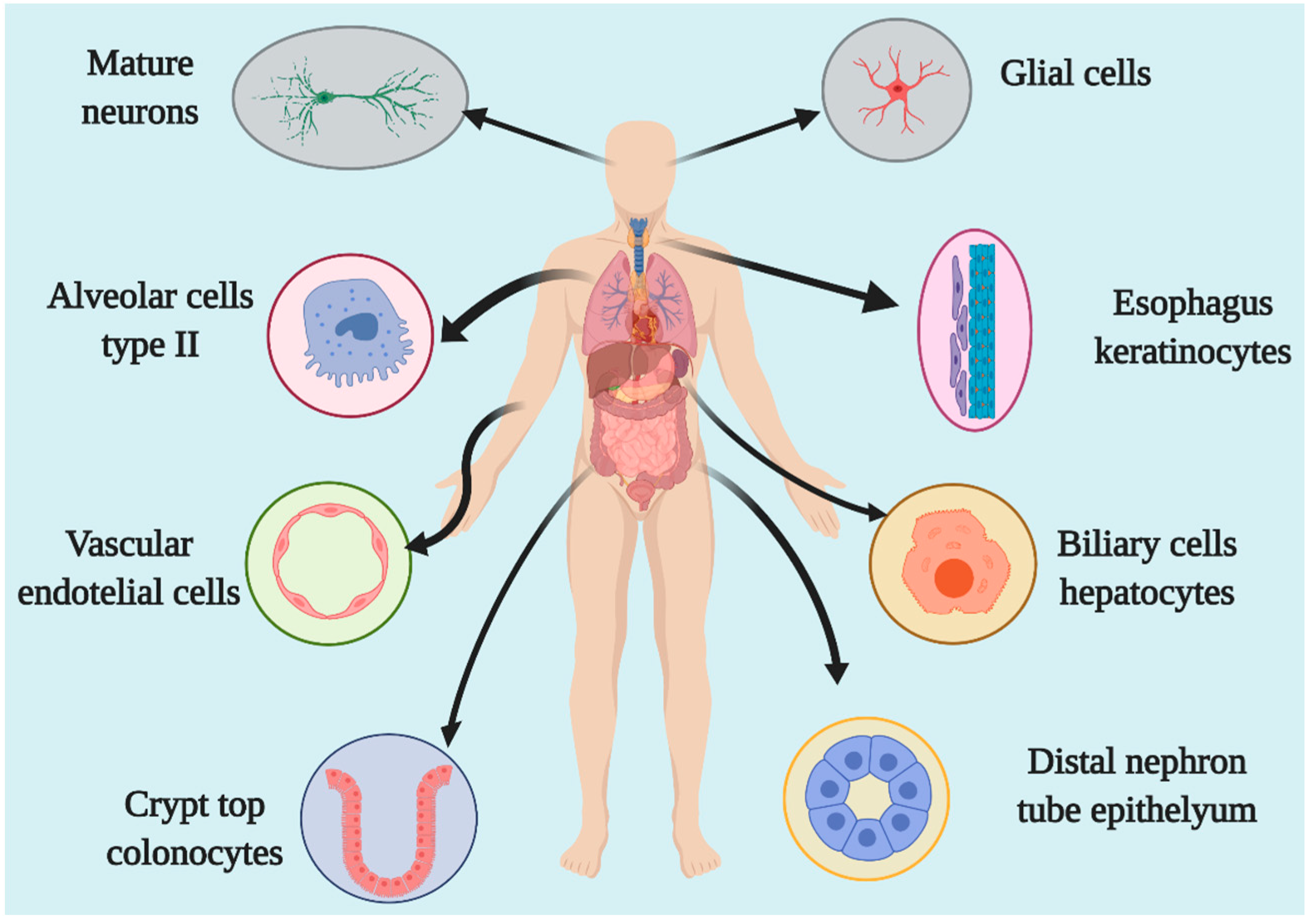
2. Location and Expression
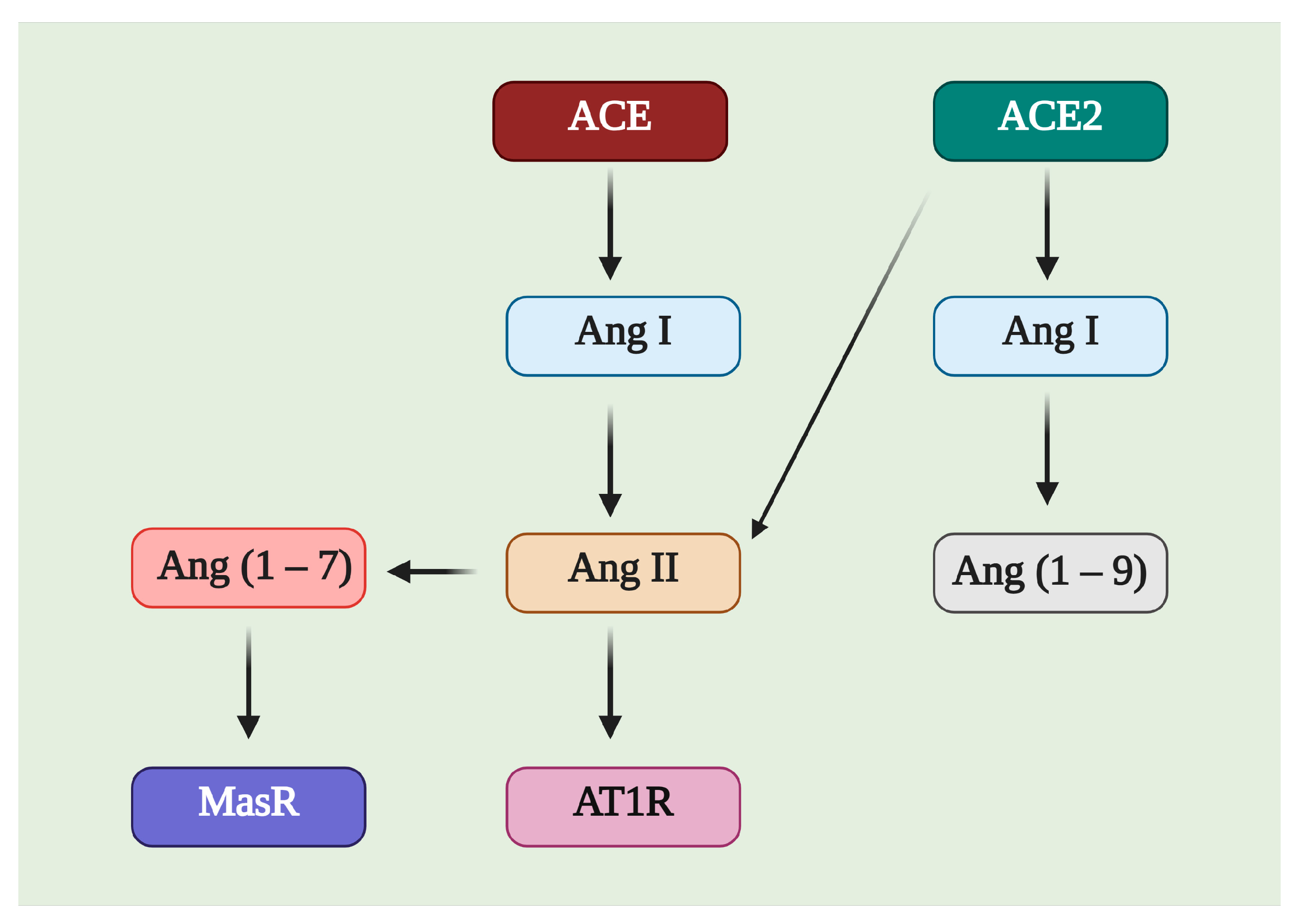
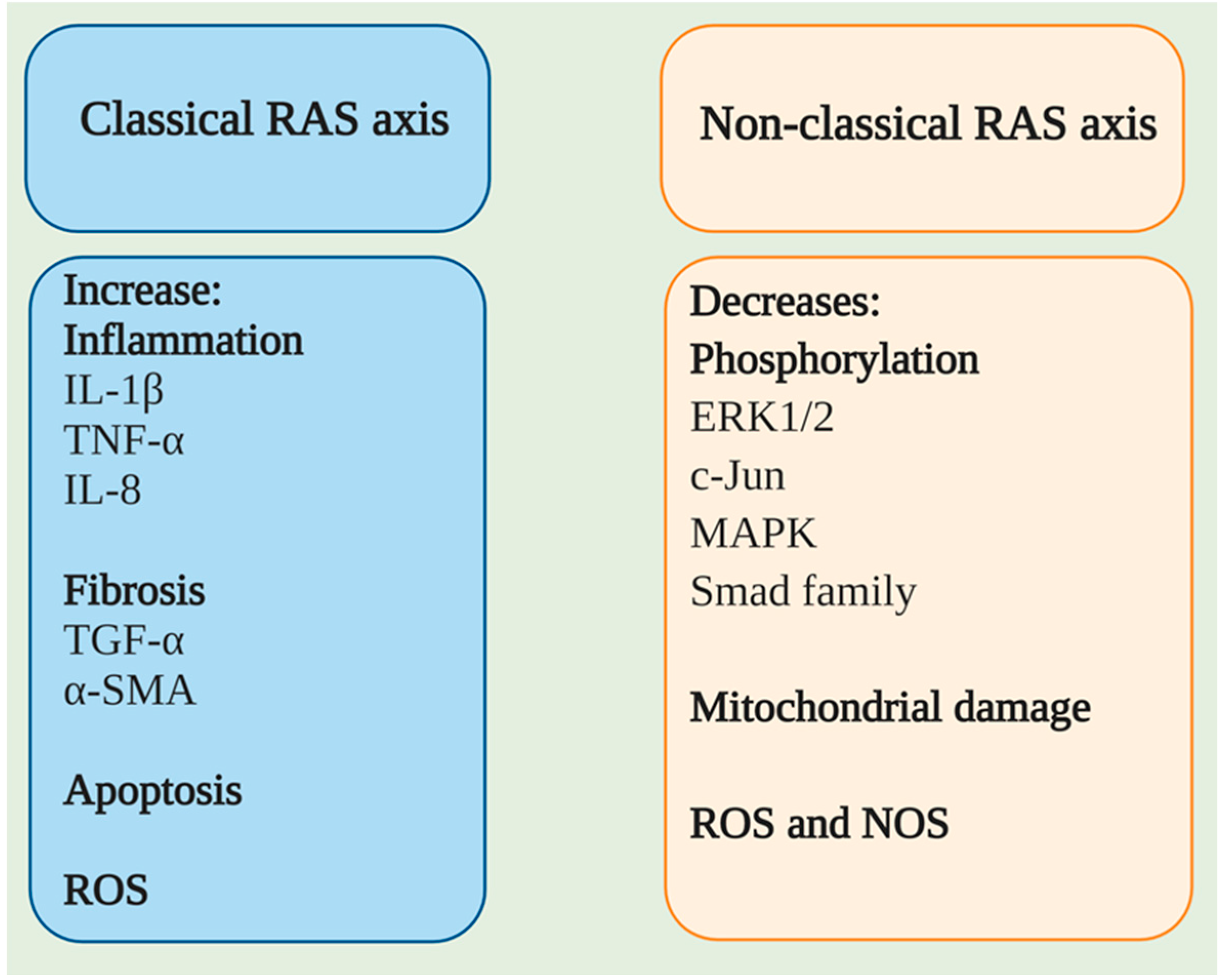
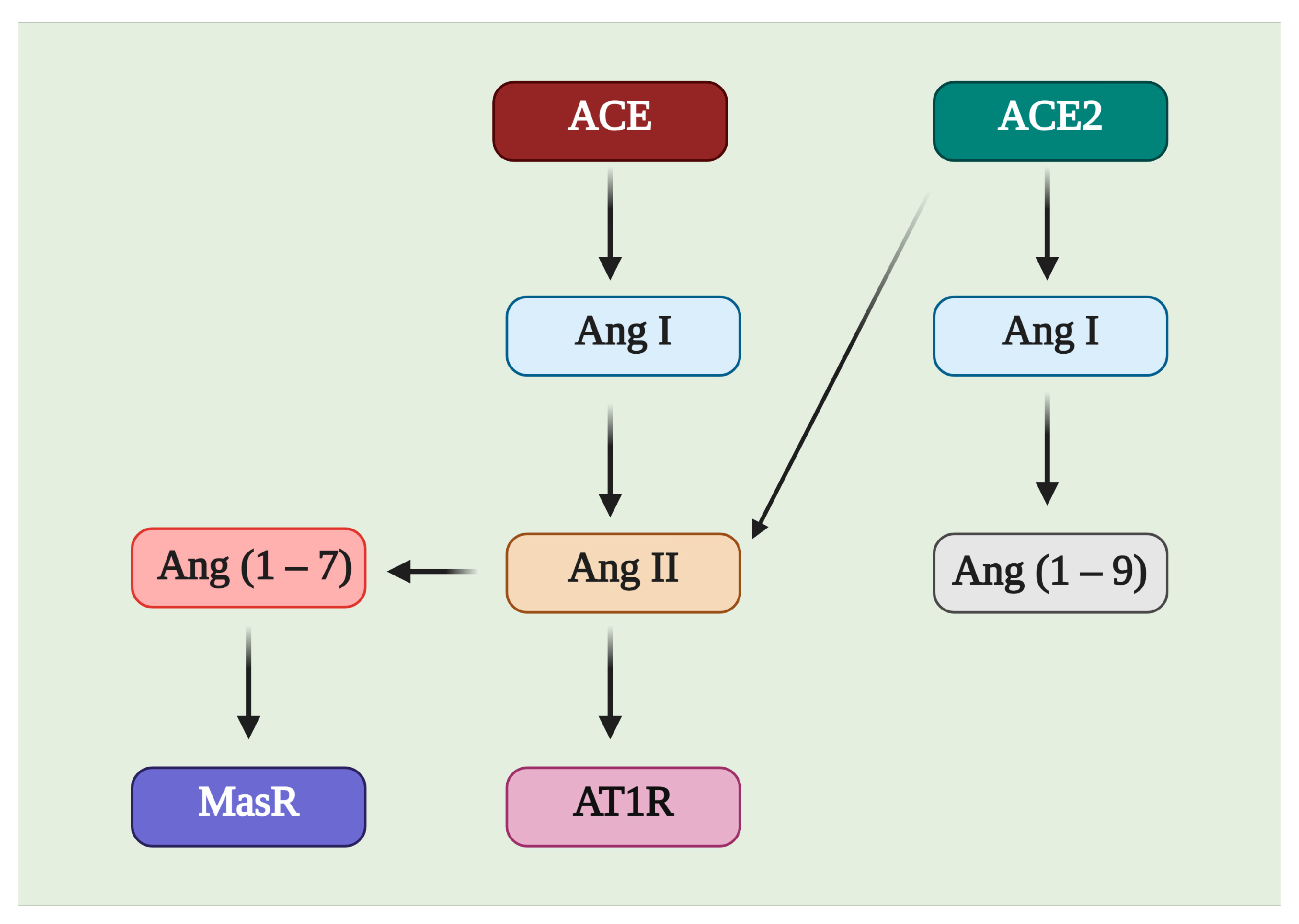
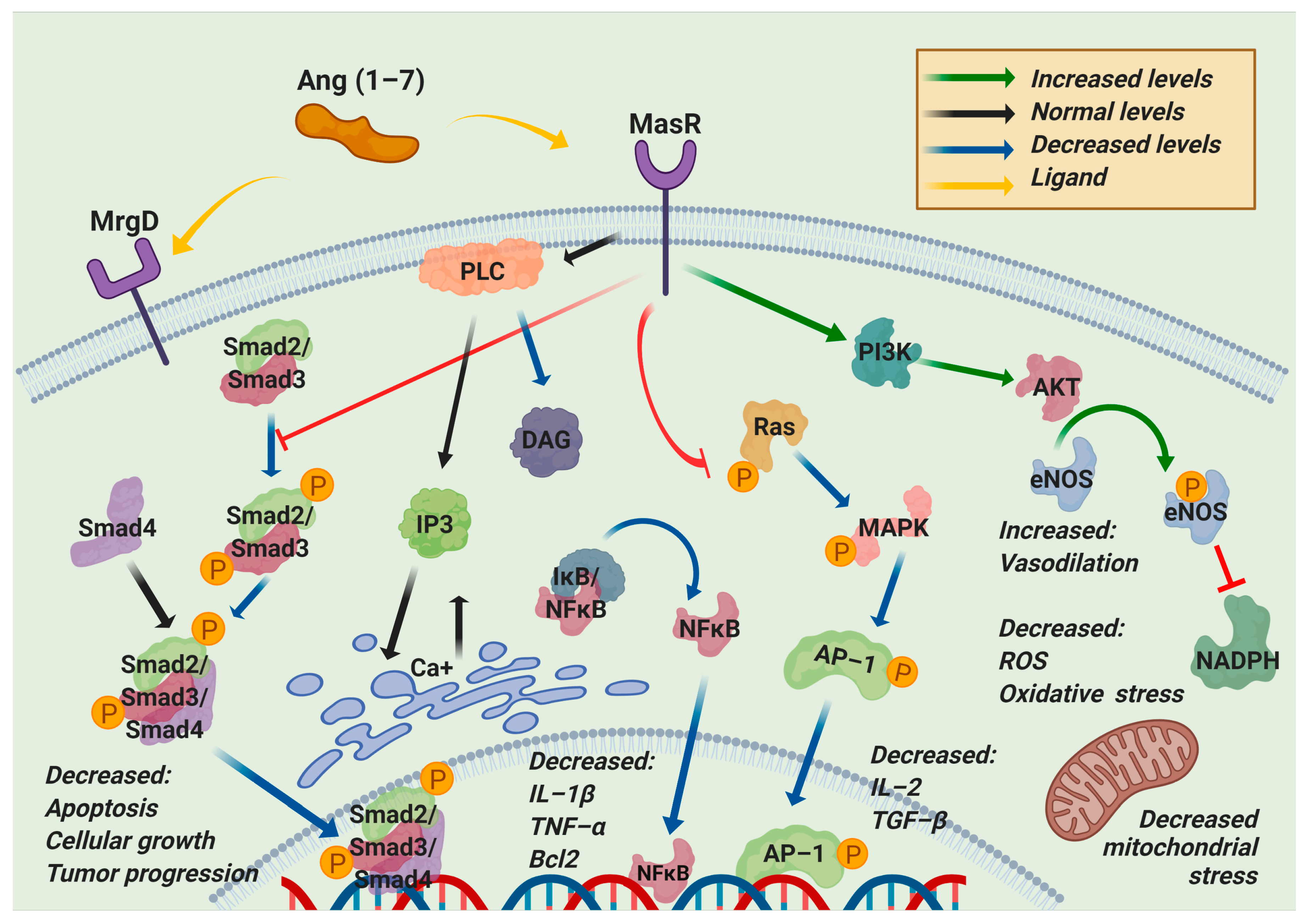
3. ACE2/Ang (1–7)/MasR Effector Axis
4. ACE2 in the Pathophysiology
4.1. Heart
4.2. Kidneys
4.3. Nervous System
4.4. Liver
4.5. Cancer
4.6. Other Conditions
5. ACE2 in Lung Conditions
SARS-CoV-2 Infection
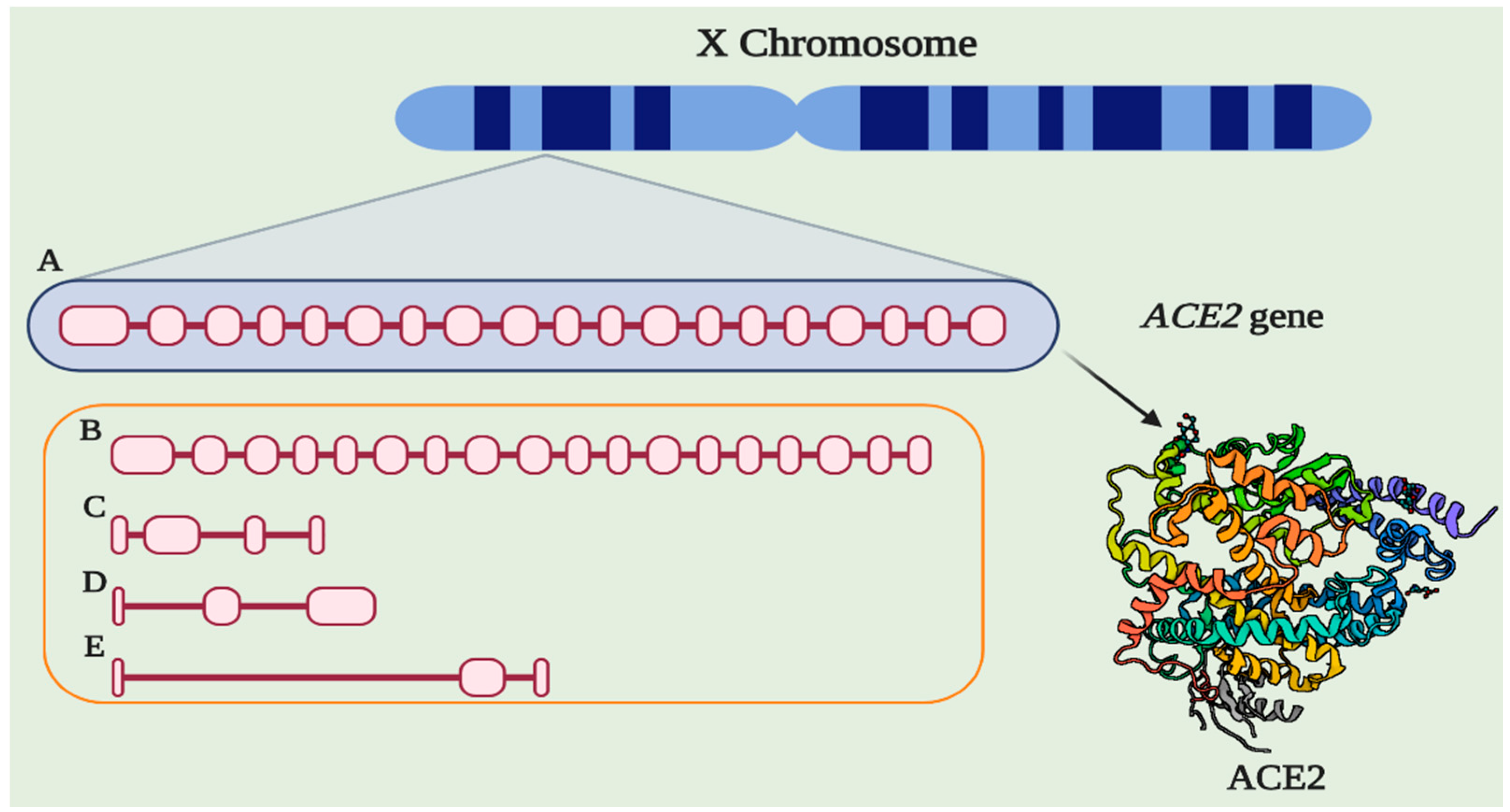
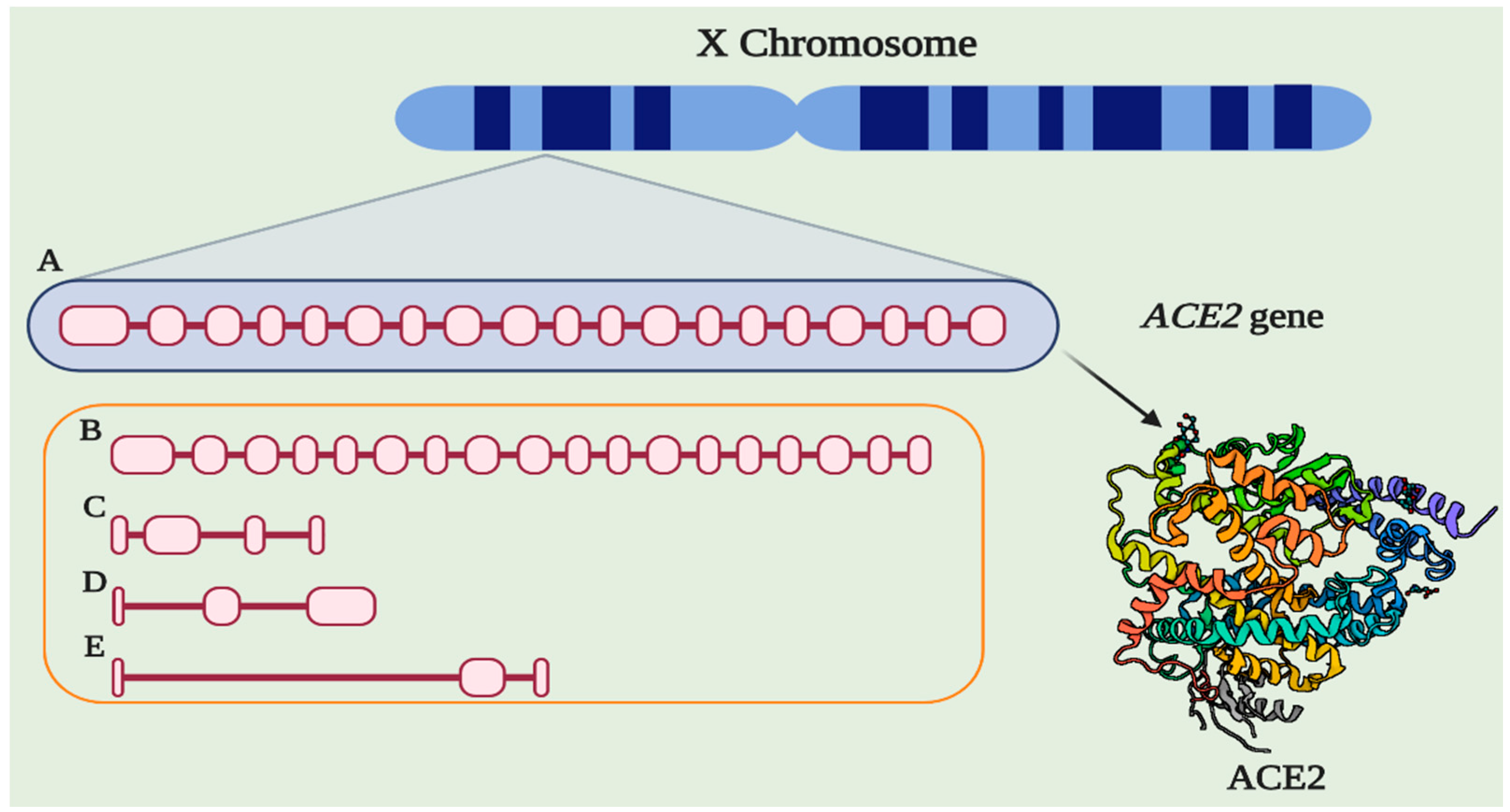
6. Genetic Aspects Related to ACE2
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gene. ACE2 (ENSG00000130234)—Summary—Homo Sapiens—Ensembl Genome Browser 99. Available online: https://www.ensembl.org/Homo_sapiens/Gene/Summary?db=core;g=ENSG00000130234;r=X:15561033-15602148 (accessed on 25 March 2020).
- Zisman, L.S.; Keller, R.S.; Weaver, B.; Lin, Q.; Speth, R.; Bristow, M.R.; Canver, C.C. Increased Angiotensin-(1–7)-Forming Activity in Failing Human Heart Ventricles: Evidence for Upregulation of the Angiotensin-Converting Enzyme Homologue ACE2. Circulation 2003, 108, 1707–1712. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.J.; Liu, H.; Wang, Z.H.; Liu, C.; Fan, J.Q.; Wang, Z.L.; Xu, Y.P.; Zhang, B.; Gyawali, L.; Li, Q.; et al. The Impact of Renal Denervation on the Progression of Heart Failure in a Canine Model Induced by Right Ventricular Rapid Pacing. Front. Physiol. 2020, 10, 1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukerjee, S.; Gao, H.; Xu, J.; Sato, R.; Zsombok, A.; Lazartigues, E. ACE2 and ADAM17 Interaction Regulates the Activity of Presympathetic Neurons. Hypertension 2019, 74, 1181–1191. [Google Scholar] [CrossRef]
- Sharma, N.; Malek, V.; Mulay, S.R.; Gaikwad, A.B. Angiotensin II Type 2 Receptor and Angiotensin-Converting Enzyme 2 Mediate Ischemic Renal Injury in Diabetic and Non-Diabetic Rats. Life Sci. 2019, 235, 116796. [Google Scholar] [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-Cell RNA-Seq Data Analysis on the Receptor ACE2 Expression Reveals the Potential Risk of Different Human Organs Vulnerable to 2019-NCoV Infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, A.M.; Khaleeq, A.; Ali, U.; Syeda, H. Evidence of the COVID-19 Virus Targeting the CNS: Tissue Distribution, Host–Virus Interaction, and Proposed Neurotropic Mechanisms. ACS Chem. Neurosci. 2020, 17, 995–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, C.E.; Miners, J.S.; Piva, G.; Willis, C.L.; Heard, D.M.; Kidd, E.J.; Good, M.A.; Kehoe, P.G. ACE2 Activation Protects against Cognitive Decline and Reduces Amyloid Pathology in the Tg2576 Mouse Model of Alzheimer’s Disease. Acta Neuropathol. 2020, 139, 485–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Garabelli, P.J.; Modrall, J.G.; Penninger, J.M.; Ferrario, C.M.; Chappell, M.C. Distinct Roles for Angiotensin-Converting Enzyme 2 and Carboxypeptidase A in the Processing of Angiotensins within the Murine Heart. Exp. Physiol. 2008, 93, 613–621. [Google Scholar] [CrossRef]
- Qi, F.; Qian, S.; Zhang, S.; Zhang, Z. Single Cell RNA Sequencing of 13 Human Tissues Identify Cell Types and Receptors of Human Coronaviruses. Biochem. Biophys. Res. Commun. 2020, 561, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Chappell, M.C. Biochemical Evaluation of the Renin-Angiotensin System: The Good, Bad, and Absolute? Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H137–H152. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhong, L.; Deng, J.; Peng, J.; Dan, H.; Zeng, X.; Li, T.; Chen, Q. High Expression of ACE2 Receptor of 2019-NCoV on the Epithelial Cells of Oral Mucosa. Int. J. Oral Sci. 2020, 12, 8. [Google Scholar] [CrossRef]
- Lukassen, S.; Lorenz Chua, R.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-CoV-2 Receptor ACE2 and TMPRSS2 Are Primarily Expressed in Bronchial Transient Secretory Cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, Y.H.; Jung, S.W.; Kim, D.J.; Park, S.H.; Song, S.J.; Jeong, K.H.; Moon, J.Y.; Ihm, C.-G.; Lee, T.W.; et al. Sex-Related Differences in the Intratubular Renin-Angiotensin System in Two-Kidney, One-Clip Hypertensive Rats. Am. J. Physiol. Ren. Physiol. 2019, 317, F670–F682. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; He, X.; Zhang, L.; Ran, Q.; Wang, J.; Xiong, A.; Wu, D.; Chen, F.; Sun, J.; Chang, C. Assessing ACE2 Expression Patterns in Lung Tissues in the Pathogenesis of COVID-J. J. Autoimmun. 2020, 112, 102463. [Google Scholar] [CrossRef] [PubMed]
- Radzikowska, U.; Ding, M.; Tan, G.; Zhakparov, D.; Peng, Y.; Wawrzyniak, P.; Wang, M.; Li, S.; Morita, H.; Altunbulakli, C.; et al. Distribution of ACE2, CD147, CD26, and Other SARS-CoV-2 Associated Molecules in Tissues and Immune Cells in Health and in Asthma, COPD, Obesity, Hypertension, and COVID-19 Risk Factors. Allergy 2020, 75, 2829–2845. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Wolf, J.; Auw-Haedrich, C.; Schlecht, A.; Boneva, S.; Lapp, T.; Horres, R.; Agostini, H.; Martin, G.; Reinhard, T.; et al. Expression of the COVID-19 Receptor ACE2 in the Human Conjunctiva. J. Med. Virol. 2020, 92, 2081–2086. [Google Scholar] [CrossRef]
- Serfozo, P.; Wysocki, J.; Gulua, G.; Schulze, A.; Ye, M.; Liu, P.; Jin, J.; Bader, M.; Myöhänen, T.; García-Horsman, J.A.; et al. Ang II (Angiotensin II) Conversion to Angiotensin-(1–7) in the Circulation Is POP (Prolyloligopeptidase)-Dependent and ACE2 (Angiotensin-Converting Enzyme 2)-Independent. Hypertension 2020, 75, 173–182. [Google Scholar] [CrossRef]
- Tao, M.-X.; Xue, X.; Gao, L.; Lu, J.-L.; Zhou, J.-S.; Jiang, T.; Zhang, Y.-D. Involvement of Angiotensin-(1–7) in the Neuroprotection of Captopril against Focal Cerebral Ischemia. Neurosci. Lett. 2018, 687, 16–21. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, S.; Li, T.; Yu, L.; Zhang, Y.; Zeng, H.; Qian, X.; Bi, J.; Lin, Y. ACE2 Inhibits Breast Cancer Angiogenesis via Suppressing the VEGFa/VEGFR2/ERK Pathway. J. Exp. Clin. Cancer Res. 2019, 38, 173. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, S.; Bihl, J. Exosome-Mediated Transfer of ACE2 (Angiotensin-Converting Enzyme 2) from Endothelial Progenitor Cells Promotes Survival and Function of Endothelial Cell. Oxid. Med. Cell. Longev. 2020, 2020, 4213541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, H.; Gao, F.; Xie, G.; Liu, Z. Angiotensin-Converting Enzyme 2 Inhibits Apoptosis of Pulmonary Endothelial Cells during Acute Lung Injury Through Suppressing MiR-4262. Cell. Physiol. Biochem. 2015, 37, 759–767. [Google Scholar] [CrossRef]
- Gunarathne, L.S.; Angus, P.W.; Herath, C.B. Blockade of Mas Receptor or Mas-Related G-Protein Coupled Receptor Type D Reduces Portal Pressure in Cirrhotic but Not in Non-Cirrhotic Portal Hypertensive Rats. Front. Physiol. 2019, 10, 1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.S.; Kim, I.J.; Kim, C.S.; Ma, S.K.; Scholey, J.W.; Kim, S.W.; Bae, E.H. Angiotensin-[1–7] Attenuates Kidney Injury in Experimental Alport Syndrome. Sci. Rep. 2020, 10, 4225. [Google Scholar] [CrossRef] [PubMed]
- Hekmat, A.S.; Zare, N.; Moravej, A.; Meshkibaf, M.H.; Javanmardi, K. Effect of Prolonged Infusion of Alamandine on Cardiovascular Parameters and Cardiac ACE2 Expression in a Rat Model of Renovascular Hypertension. Biol. Pharm. Bull. 2019, 42, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Ali, R.M.; Al-Shorbagy, M.Y.; Helmy, M.W.; El-Abhar, H.S. Role of Wnt4/β-Catenin, Ang II/TGFβ, ACE2, NF-κB, and IL-18 in Attenuating Renal Ischemia/Reperfusion-Induced Injury in Rats Treated with Vit D and Pioglitazone. Eur. J. Pharmacol. 2018, 831, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Stoll, D.; Yokota, R.; Sanches Aragão, D.; Casarini, D.E. Both Aldosterone and Spironolactone Can Modulate the Intracellular ACE/ANG II/AT1 and ACE2/ANG (1–7)/MAS Receptor Axes in Human Mesangial Cells. Physiol. Rep. 2019, 7, e14105. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Hu, B.; Qu, H.; Wang, L.; Huang, X.; Li, M.; Zhang, M. Upregulation of Angiotensin Converting Enzyme 2 by Shear Stress Reduced Inflammation and Proliferation in Vascular Endothelial Cells. Biochem. Biophys. Res. Commun. 2020, 525, 812–818. [Google Scholar] [CrossRef]
- Wu, H.T.; Chuang, Y.W.; Huang, C.P.; Chang, M.H. Loss of Angiotensin Converting Enzyme II (ACE2) Accelerates the Development of Liver Injury Induced by Thioacetamide. Exp. Anim. 2018, 67, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Kangussu, L.M.; De Almeida, T.C.S.; Prestes, T.R.R.; De Andrade De Maria, M.L.; Da Silva Filha, R.; Vieira, M.A.R.; Silva, A.C.S.E.; Ferreira, A.J. Beneficial Effects of the Angiotensin-Converting Enzyme 2 Activator Dize in Renovascular Hypertension. Protein Pept. Lett. 2019, 26, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Dang, Z.; Su, S.; Jin, G.; Nan, X.; Ma, L.; Li, Z.; Lu, D.; Ge, R. Tsantan Sumtang Attenuated Chronic Hypoxia-Induced Right Ventricular Structure Remodeling and Fibrosis by Equilibrating Local ACE-AngII-AT1R/ACE2-Ang 1–7-Mas Axis in Rat. J. Ethnopharmacol. 2020, 250, 112470. [Google Scholar] [CrossRef]
- Li, S.; Zhao, W.; Tao, Y.; Liu, C. Fugan Wan Alleviates Hepatic Fibrosis by Inhibiting ACE/Ang II/AT-1R Signaling Pathway and Enhancing ACE2/Ang 1–7/Mas Signaling Pathway in Hepatic Fibrosis Rat Models. Am. J. Transl. Res. 2020, 12, 592–601. [Google Scholar] [PubMed]
- Ramchand, J.; Patel, S.K.; Kearney, L.G.; Matalanis, G.; Farouque, O.; Srivastava, P.M.; Burrell, L.M. Plasma ACE2 Activity Predicts Mortality in Aortic Stenosis and Is Associated With Severe Myocardial Fibrosis. JACC Cardiovasc. Imaging 2020, 13, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, J.; Schulze, A.; Batlle, D. Novel Variants of Angiotensin Converting Enzyme-2 of Shorter Molecular Size to Target the Kidney Renin Angiotensin System. Biomolecules 2019, 9, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.A.; Gallardo, M.; Cespedes, C.; Vio, C.P. Potassium Intake Prevents the Induction of the Renin-Angiotensin System and Increases Medullary ACE2 and COX-2 in the Kidneys of Angiotensin II-Dependent Hypertensive Rats. Front. Pharmacol. 2019, 10, 1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamel, A.S.; Abdelkader, N.F.; Abd El-Rahman, S.S.; Emara, M.; Zaki, H.F.; Khattab, M.M. Stimulation of ACE2/ANG(1–7)/Mas Axis by Diminazene Ameliorates Alzheimer’s Disease in the D-Galactose-Ovariectomized Rat Model: Role of PI3K/Akt Pathway. Mol. Neurobiol. 2018, 55, 8188–8202. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, N.F.; Abd El-Latif, A.M.; Khattab, M.M. Telmisartan/17β-Estradiol Mitigated Cognitive Deficit in an Ovariectomized Rat Model of Alzheimer’s Disease: Modulation of ACE1/ACE2 and AT1/AT2 Ratio. Life Sci. 2020, 245, 117388. [Google Scholar] [CrossRef]
- Cui, C.; Xu, P.; Li, G.; Qiao, Y.; Han, W.; Geng, C.; Liao, D.; Yang, M.; Chen, D.; Jiang, P. Vitamin D Receptor Activation Regulates Microglia Polarization and Oxidative Stress in Spontaneously Hypertensive Rats and Angiotensin II-Exposed Microglial Cells: Role of Renin-Angiotensin System. Redox Biol. 2019, 26, 101295. [Google Scholar] [CrossRef] [PubMed]
- Guan, G.W.; Gao, L.; Wang, J.W.; Wen, X.J.; Mao, T.H.; Peng, S.W.; Zhang, T.; Chen, X.M.; Lu, F.M. Exploring the Mechanism of Liver Enzyme Abnormalities in Patients with Novel Coronavirus-Infected Pneumonia. Chin. J. Hepatol. 2020, 28, E002. [Google Scholar] [CrossRef]
- Rajapaksha, I.G.; Gunarathne, L.S.; Asadi, K.; Cunningham, S.C.; Sharland, A.; Alexander, I.E.; Angus, P.W.; Herath, C.B. Liver-Targeted Angiotensin Converting Enzyme 2 Therapy Inhibits Chronic Biliary Fibrosis in Multiple Drug-Resistant Gene 2-Knockout Mice. Hepatol. Commun. 2019, 3, 1656–1673. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Hu, Z.; Chen, Q.; Huang, Y.; Mao, Z.; Xu, F.; Zhou, X. BML-111 Equilibrated ACE-AngII-AT1R and ACE2-Ang-(1–7)-Mas Axis to Protect Hepatic Fibrosis in Rats. Prostaglandins Other Lipid Mediat. 2017, 131, 75–82. [Google Scholar] [CrossRef]
- Casey, S.; Schierwagen, R.; Mak, K.Y.; Klein, S.; Uschner, F.; Jansen, C.; Praktiknjo, M.; Meyer, C.; Thomas, D.; Herath, C.; et al. Activation of the Alternate Renin-Angiotensin System Correlates with the Clinical Status in Human Cirrhosis and Corrects Post Liver Transplantation. J. Clin. Med. 2019, 8, 419. [Google Scholar] [CrossRef] [Green Version]
- Narayan, S.S.; Lorenz, K.; Ukkat, J.; Hoang-Vu, C.; Trojanowicz, B. Angiotensin Converting Enzymes ACE and ACE2 in Thyroid Cancer Progression. Neoplasma 2019, 67, 402–409. [Google Scholar] [CrossRef]
- Yamagata, R.; Nemoto, W.; Nakagawasai, O.; Takahashi, K.; Tan-No, K. Downregulation of Spinal Angiotensin Converting Enzyme 2 Is Involved in Neuropathic Pain Associated with Type 2 Diabetes Mellitus in Mice. Biochem. Pharmacol. 2020, 174, 113825. [Google Scholar] [CrossRef]
- Wong, M.H.; Chapin, O.C.; Johnson, M.D. LPS-Stimulated Cytokine Production in Type I Cells Is Modulated by the Renin-Angiotensin System. Am. J. Respir. Cell Mol. Biol. 2012, 46, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zheng, J.; Yan, Y.; Ruan, Z.; Su, Y.; Wang, J.; Huang, H.; Zhang, Y.; Wang, W.; Gao, J.; et al. Angiotensin-Converting Enzyme 2 Regulates Autophagy in Acute Lung Injury through AMPK/MTOR Signaling. Arch. Biochem. Biophys. 2019, 672, 108061. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Lin, Q.; Qin, X.; Ruan, Z.; Zhou, J.; Lin, Z.; Su, Y.; Zheng, J.; Liu, Z. ACE2 Antagonizes VEGFa to Reduce Vascular Permeability during Acute Lung Injury. Cell. Physiol. Biochem. 2016, 38, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.I.; Tsai, C.H.; Sun, Y.L.; Hsieh, W.Y.; Lin, Y.C.; Chen, C.Y.; Lin, C.S. Instillation of Particulate Matter 2.5 Induced Acute Lung Injury and Attenuated the Injury Recovery in ACE2 Knockout Mice. Int. J. Biol. Sci. 2018, 14, 253–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.; Liu, Y. Osthole Alleviates Bleomycin-Induced Pulmonary Fibrosis via Modulating Angiotensin-Converting Enzyme 2/Angiotensin-(1–7) Axis and Decreasing Inflammation Responses in Rats. Biol. Pharm. Bull. 2016, 39, 457–465. [Google Scholar] [CrossRef]
- Min, F.; Gao, F.; Li, Q.; Liu, Z. Therapeutic Effect of Human Umbilical Cord Mesenchymal Stem Cells Modifed by Angiotensin-Converting Enzyme 2 Gene on Bleomycin-Induced Lung Fibrosis Injury. Mol. Med. Rep. 2015, 11, 2387–2396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Zhang, X.; Xu, H.; Gao, X.M.; Zhang, G.Z.; Zhang, H.; Yang, F. Dynamic Variation of RAS on Silicotic Fibrosis Pathogenesis in Rats. Curr. Med. Sci. 2019, 39, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.N.; Xu, H.; Gao, X.M.; Zhang, G.Z.; Zhang, X.; Yang, F. Protective Effect of Angiotensin (1–7) on Silicotic Fibrosis in Rats. Biomed. Environ. Sci. 2019, 32, 419–426. [Google Scholar] [CrossRef]
- Sandoval, J.; Del Valle-Mondragón, L.; Masso, F.; Zayas, N.; Pulido, T.; Teijeiro, R.; Gonzalez-Pacheco, H.; Olmedo-Ocampo, R.; Sisniega, C.; Paez-Arenas, A.; et al. Angiotensin Converting Enzyme 2 and Angiotensin (1–7) Axis in Pulmonary Arterial Hypertension. Eur. Respir. J. 2020, 56, 1902416. [Google Scholar] [CrossRef]
- Kou, Y.L.; Zhang, P.P.; Wang, H.Y.; Zhang, J.B.; Tan, X.S.; Huang, C.; Zhang, M. Protective Effect of Angiotensin Converting Enzyme 2 (ACE2) against Chronic Intermittent Hypoxia-Induced Pulmonary Oxidative Stress Injury in Rats. J. Sichuan Univ. Med. Sci. Ed. 2016, 47, 43–48. [Google Scholar]
- Xiao, H.L.; Zhao, L.X.; Yang, J.; Tong, N.; An, L.; Liu, Q.T.; Xie, M.R.; Li, C.S. Association between ACE2/ACE Balance and Pneumocyte Apoptosis in a Porcine Model of Acute Pulmonary Thromboembolism with Cardiac Arrest. Mol. Med. Rep. 2018, 17, 4221–4228. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, J.; Chen, J.; Luo, Q.; Zhang, Q.; Zhang, H. Vitamin D Alleviates Lipopolysaccharide-Induced Acute Lung Injury via Regulation of the Renin-Angiotensin System. Mol. Med. Rep. 2017, 16, 7432–7438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Gao, Y.; Zhao, W.; Yu, G.; Jin, F. ACE-2/ANG1–7 Ameliorates ER Stress-Induced Apoptosis in Seawater Aspiration-Induced Acute Lung Injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L1015–L1027. [Google Scholar] [CrossRef] [PubMed]
- Supé, S.; Kohse, F.; Gembardt, F.; Kuebler, W.M.; Walther, T. Therapeutic Time Window for Angiotensin-(1–7) in Acute Lung Injury. Br. J. Pharmacol. 2016, 173, 1618–1628. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Xie, Z.; Li, T.; Zhang, S.; Lai, C.; Zhu, P.; Wang, K.; Han, L.; Duan, Y.; Zhao, Z.; et al. Angiotensin-Converting Enzyme 2 Inhibits Lung Injury Induced by Respiratory Syncytial Virus. Sci. Rep. 2016, 6, 19840. [Google Scholar] [CrossRef]
- Yang, P.; Gu, H.; Zhao, Z.; Wang, W.; Cao, B.; Lai, C.; Yang, X.; Zhang, L.Y.; Duan, Y.; Zhang, S.; et al. Angiotensin-Converting Enzyme 2 (ACE2) Mediates Influenza H7N9 Virus-Induced Acute Lung Injury. Sci. Rep. 2014, 4, 7027. [Google Scholar] [CrossRef] [Green Version]
- Glowacka, I.; Bertram, S.; Herzog, P.; Pfefferle, S.; Steffen, I.; Muench, M.O.; Simmons, G.; Hofmann, H.; Kuri, T.; Weber, F.; et al. Differential Downregulation of ACE2 by the Spike Proteins of Severe Acute Respiratory Syndrome Coronavirus and Human Coronavirus NL. J. Virol. 2010, 84, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Menachery, V.D.; Yount, B.L.; Debbink, K.; Agnihothram, S.; Gralinski, L.E.; Plante, J.A.; Graham, R.L.; Scobey, T.; Ge, X.Y.; Donaldson, E.F.; et al. A SARS-like Cluster of Circulating Bat Coronaviruses Shows Potential for Human Emergence. Nat. Med. 2015, 21, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.T.; Serrano, M.L.; Pujol, F.H.; Rangel, H.R. Role of Changes in SARS-CoV-2 Spike Protein in the Interaction with the Human ACE2 Receptor: An in Silico Analysis. EXCLI J. 2020, 19, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhao, Y.-B.; Wang, Q.; Li, J.-Y.; Zhou, Z.-J.; Liao, C.-H.; Ge, X.-Y. Predicting the Angiotensin Converting Enzyme 2 (ACE2) Utilizing Capability as the Receptor of SARS-CoV-2. Microbes Infect. 2020, 22, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Zhou, Y.; Du, L. Characterization of the Receptor-Binding Domain (RBD) of 2019 Novel Coronavirus: Implication for Development of RBD Protein as a Viral Attachment Inhibitor and Vaccine. Cell. Mol. Immunol. 2020, 17, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, J.; Lu, Y.; Jin, X.; Zhang, L. Spike Protein Recognition of Mammalian ACE2 Predicts the Host Range and an Optimized ACE2 for SARS-CoV-2 Infection. Biochem. Biophys. Res. Commun. 2020, 526, 165–169. [Google Scholar] [CrossRef]
- Zhang, P.; Leu, J.I.J.; Murphy, M.E.; George, D.L.; Marmorstein, R. Crystal Structure of the Stress-Inducible Human Heat Shock Protein 70 Substrate-Binding Domain in Complex with Peptide Substrate. PLoS ONE 2014, 9, e103518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.-Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Woudenberg, T.; Pelleau, S.; Anna, F.; Attia, M.; Donnadieu, F.; Gravet, A.; Lohmann, C.; Seraphin, H.; Guiheneuf, R.; Delamare, C.; et al. Collateral Effects of COVID-19 Pandemic Emergency Response on Worldwide Immunizations. EBioMedicine 2021, 70, 103495. [Google Scholar] [CrossRef]
- Reche, P.A. Potential Cross-Reactive Immunity to SARS-CoV-2 From Common Human Pathogens and Vaccines. Front. Immunol. 2020, 11, 2694. [Google Scholar] [CrossRef]
- Urbán, S.; Paragi, G.; Burián, K.; McLean, G.R.; Virok, D.P. Identification of Similar Epitopes between Severe Acute Respiratory Syndrome Coronavirus-2 and Bacillus Calmette–Guérin: Potential for Cross-Reactive Adaptive Immunity. Clin. Transl. Immunol. 2020, 9, e1227. [Google Scholar] [CrossRef]
- Lee, C.H.; Pinho, M.P.; Buckley, P.R.; Woodhouse, I.B.; Ogg, G.; Simmons, A.; Napolitani, G.; Koohy, H. Potential CD8+ T Cell Cross-Reactivity Against SARS-CoV-2 Conferred by Other Coronavirus Strains. Front. Immunol. 2020, 11, 2878. [Google Scholar] [CrossRef]
- Guo, E.; Guo, H. CD8 T Cell Epitope Generation toward the Continually Mutating SARS-CoV-2 Spike Protein in Genetically Diverse Human Population: Implications for Disease Control and Prevention. PLoS ONE 2020, 15, e0239566. [Google Scholar] [CrossRef]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A Crucial Role of Angiotensin Converting Enzyme 2 (ACE2) in SARS Coronavirus-Induced Lung Injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Guzzi, P.H.; Mercatelli, D.; Ceraolo, C.; Giorgi, F.M. Master Regulator Analysis of the SARS-CoV-2/Human Interactome. J. Clin. Med. 2020, 9, 982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Sun, C. Conjunctiva Is Not a Preferred Gateway of Entry for SARS-CoV-2 to Infect Respiratory Tract. J. Med. Virol. 2020, 92, 1410–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Li, R.; Lu, Z.; Huang, Y. Does Comorbidity Increase the Risk of Patients with COVID-19: Evidence from Meta-Analysis. Aging 2020, 12, 6049. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Wang, Y.; Wang, G.-Q. Organ-Protective Effect of Angiotensin-Converting Enzyme 2 and Its Effect on the Prognosis of COVID-19. J. Med. Virol. 2020, 92, 726–730. [Google Scholar] [CrossRef]
- Yang, J.; Zheng, Y.; Gou, X.; Pu, K.; Chen, Z.; Guo, Q.; Ji, R.; Wang, H.; Wang, Y.; Zhou, Y. Prevalence of Comorbidities and Its Effects in Patients Infected with SARS-CoV-2: A Systematic Review and Meta-Analysis. Int. J. Infect. Dis. 2020, 94, 91–95. [Google Scholar] [CrossRef]
- Hamet, P.; Pausova, Z.; Attaoua, R.; Hishmih, C.; Haloui, M.; Shin, J.; Paus, T.; Abrahamowicz, M.; Gaudet, D.; Santucci, L.; et al. SARS-CoV-2 Receptor ACE2 Gene Is Associated with Hypertension and Severity of COVID 19: Interaction with Sex, Obesity, and Smoking. Am. J. Hypertens. 2021, 34, 367–376. [Google Scholar] [CrossRef]
- Brake, S.J.; Barnsley, K.; Lu, W.; McAlinden, K.D.; Eapen, M.S.; Sohal, S.S. Smoking Upregulates Angiotensin-Converting Enzyme-2 Receptor: A Potential Adhesion Site for Novel Coronavirus SARS-CoV-2 (COVID-19). J. Clin. Med. 2020, 9, 841. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Meng, M.; Kumar, R.; Wu, Y.; Huang, J.; Lian, N.; Deng, Y.; Lin, S. The Impact of COPD and Smoking History on the Severity of COVID-19: A Systemic Review and Meta-Analysis. J. Med. Virol. 2020, 92, 1915–1921. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Zhang, X.; Li, R.; Zheng, M.; Yang, S.; Dai, L.; Wu, A.; Hu, C.; Huang, Y.; Xie, M.; et al. Overexpression of the SARS-CoV-2 Receptor ACE2 Is Induced by Cigarette Smoke in Bronchial and Alveolar Epithelia. J. Pathol. 2021, 253, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.C.; Sausville, E.; Girish, V.; Yuan, M.L.; Vasudevan, A.; John, K.M.; Sheltzer, J.M. Cigarette Smoke Exposure and Inflammatory Signaling Increase the Expression of the SARS-CoV-2 Receptor ACE2 in the Respiratory Tract. Dev. Cell 2020, 53, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Cava, C.; Bertoli, G.; Castiglioni, I. In Silico Discovery of Candidate Drugs against COVID-19. Viruses 2020, 12, 404. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.L.; Yang, J.M.; Sun, Y.P.; Su, G.H. Inhibitors of RAS Might Be a Good Choice for the Therapy of COVID-19 Pneumonia. Chin. J. Tuberc. Respir. Dis. 2020, 43, 219–222. [Google Scholar] [CrossRef]
- Danser, A.H.J.; Epstein, M.; Batlle, D. Renin-Angiotensin System Blockers and the COVID-19 Pandemic: At Present There Is No Evidence to Abandon Renin-Angiotensin System Blockers. Hypertension 2020, 75, 1382–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellinghaus, D. Genomewide Association Study of Severe COVID-19 with Respiratory Failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Huang, S.; Gao, R.; Zhou, Y.; Lai, C.; Li, Z.; Xian, W.; Qian, X.; Li, Z.; Huang, Y.; et al. Initial Whole-Genome Sequencing and Analysis of the Host Genetic Contribution to COVID-19 Severity and Susceptibility. Cell Discov. 2020, 6, 83. [Google Scholar] [CrossRef]
- Hu, J.; Li, C.; Wang, S.; Li, T.; Zhang, H. Genetic Variants Are Identified to Increase Risk of COVID-19 Related Mortality from UK Biobank Data. Hum. Genom. 2021, 15, 10. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, T.; Li, Y.; Guan, T.; Lai, Y.; Shen, Y.; Zeyaweiding, A.; Maimaiti, T.; Li, F.; Zhao, H.; et al. Association of ACE2 Polymorphisms with Susceptibility to Essential Hypertension and Dyslipidemia in Xinjiang, China. Lipids Health Dis. 2018, 17, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, L.J.; Cañadas-Garre, M.; Cappa, R.C.; Maxwell, A.P.; McKnight, A.J. Genetic Associations between Genes in the Renin-Angiotensin-Aldosterone System and Renal Disease: A Systematic Review and Meta-Analysis. BMJ Open 2019, 9, e026777. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Li, L.; Feng, Z.; Wan, S.; Huang, P.; Sun, X.; Wen, F.; Huang, X.; Ning, G.; Wang, W. Comparative Genetic Analysis of the Novel Coronavirus (2019-NCoV/SARS-CoV-2) Receptor ACE2 in Different Populations. Cell Discov. 2020, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Jabeen, N.; Raza, F.; Shabbir, S.; Baig, A.A.; Amanullah, A.; Aziz, B. Structural Variations in Human ACE2 May Influence Its Binding with SARS-CoV-2 Spike Protein. J. Med. Virol. 2020, 92, 1580–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambrocio-Ortiz, E.; Pérez-Rubio, G.; Del Ángel-Pablo, A.D.; Buendía-Roldán, I.; Chávez-Galán, L.; Hernández-Zenteno, R.d.J.; Ramírez-Venegas, A.; Rojas-Serrano, J.; Mejía, M.; Pérez-Padilla, R.; et al. Angiotensin-Converting Enzyme 2 (ACE2) in the Context of Respiratory Diseases and Its Importance in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection. Pharmaceuticals 2021, 14, 805. https://doi.org/10.3390/ph14080805
Ambrocio-Ortiz E, Pérez-Rubio G, Del Ángel-Pablo AD, Buendía-Roldán I, Chávez-Galán L, Hernández-Zenteno RdJ, Ramírez-Venegas A, Rojas-Serrano J, Mejía M, Pérez-Padilla R, et al. Angiotensin-Converting Enzyme 2 (ACE2) in the Context of Respiratory Diseases and Its Importance in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection. Pharmaceuticals. 2021; 14(8):805. https://doi.org/10.3390/ph14080805
Chicago/Turabian StyleAmbrocio-Ortiz, Enrique, Gloria Pérez-Rubio, Alma D. Del Ángel-Pablo, Ivette Buendía-Roldán, Leslie Chávez-Galán, Rafael de Jesús Hernández-Zenteno, Alejandra Ramírez-Venegas, Jorge Rojas-Serrano, Mayra Mejía, Rogelio Pérez-Padilla, and et al. 2021. "Angiotensin-Converting Enzyme 2 (ACE2) in the Context of Respiratory Diseases and Its Importance in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection" Pharmaceuticals 14, no. 8: 805. https://doi.org/10.3390/ph14080805
APA StyleAmbrocio-Ortiz, E., Pérez-Rubio, G., Del Ángel-Pablo, A. D., Buendía-Roldán, I., Chávez-Galán, L., Hernández-Zenteno, R. d. J., Ramírez-Venegas, A., Rojas-Serrano, J., Mejía, M., Pérez-Padilla, R., Guadarrama-Pérez, C., & Falfán-Valencia, R. (2021). Angiotensin-Converting Enzyme 2 (ACE2) in the Context of Respiratory Diseases and Its Importance in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection. Pharmaceuticals, 14(8), 805. https://doi.org/10.3390/ph14080805