
Fucoidan Protects against Acute Sulfoxaflor-Induced Hematological/Biochemical Alterations and Oxidative Stress in Male Mice

Abstract
:1. Introduction
2. Results
2.1. Changes in Hematological Parameters
2.2. Changes in Serum Biochemical Markers of Liver and Kidney Functions
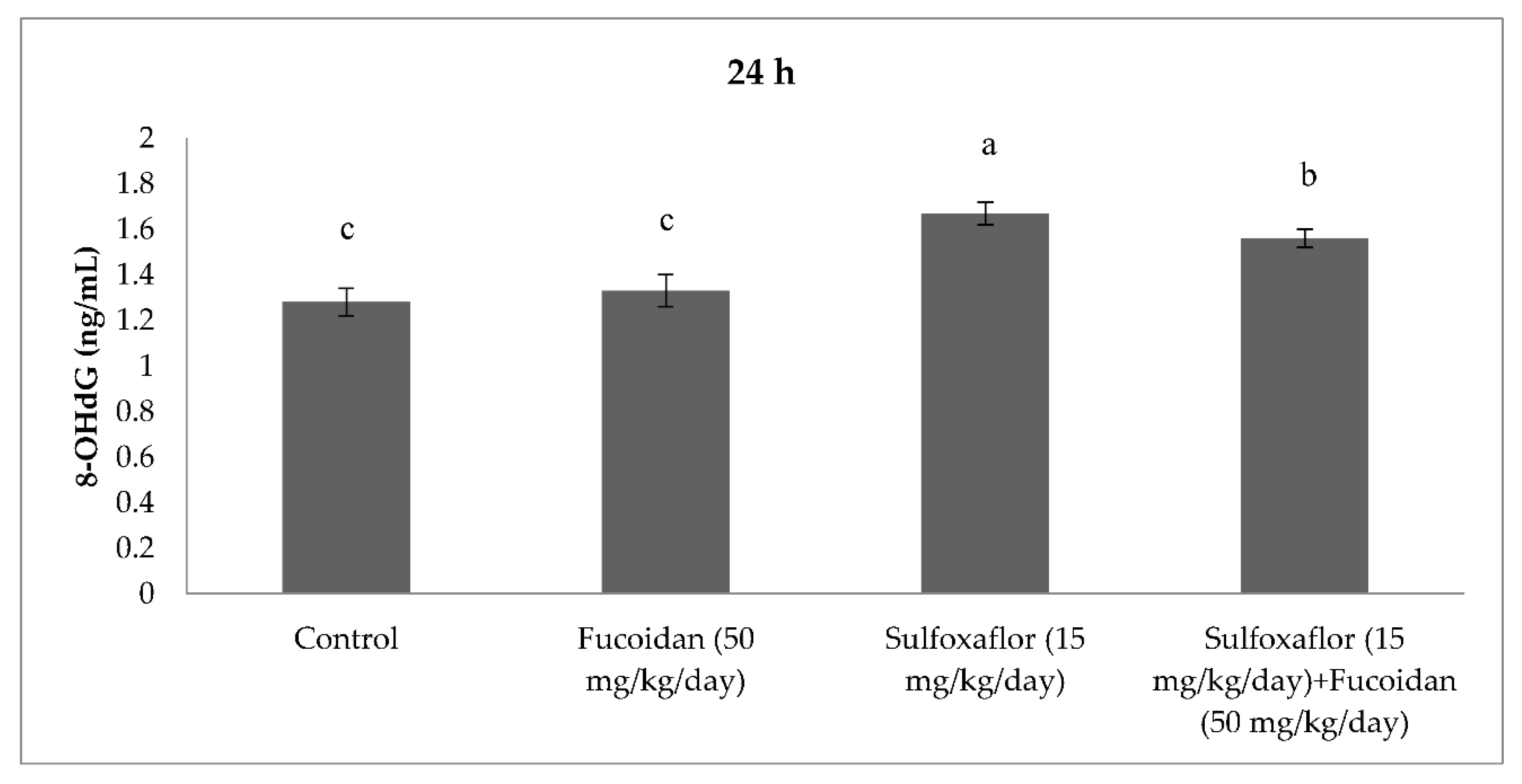
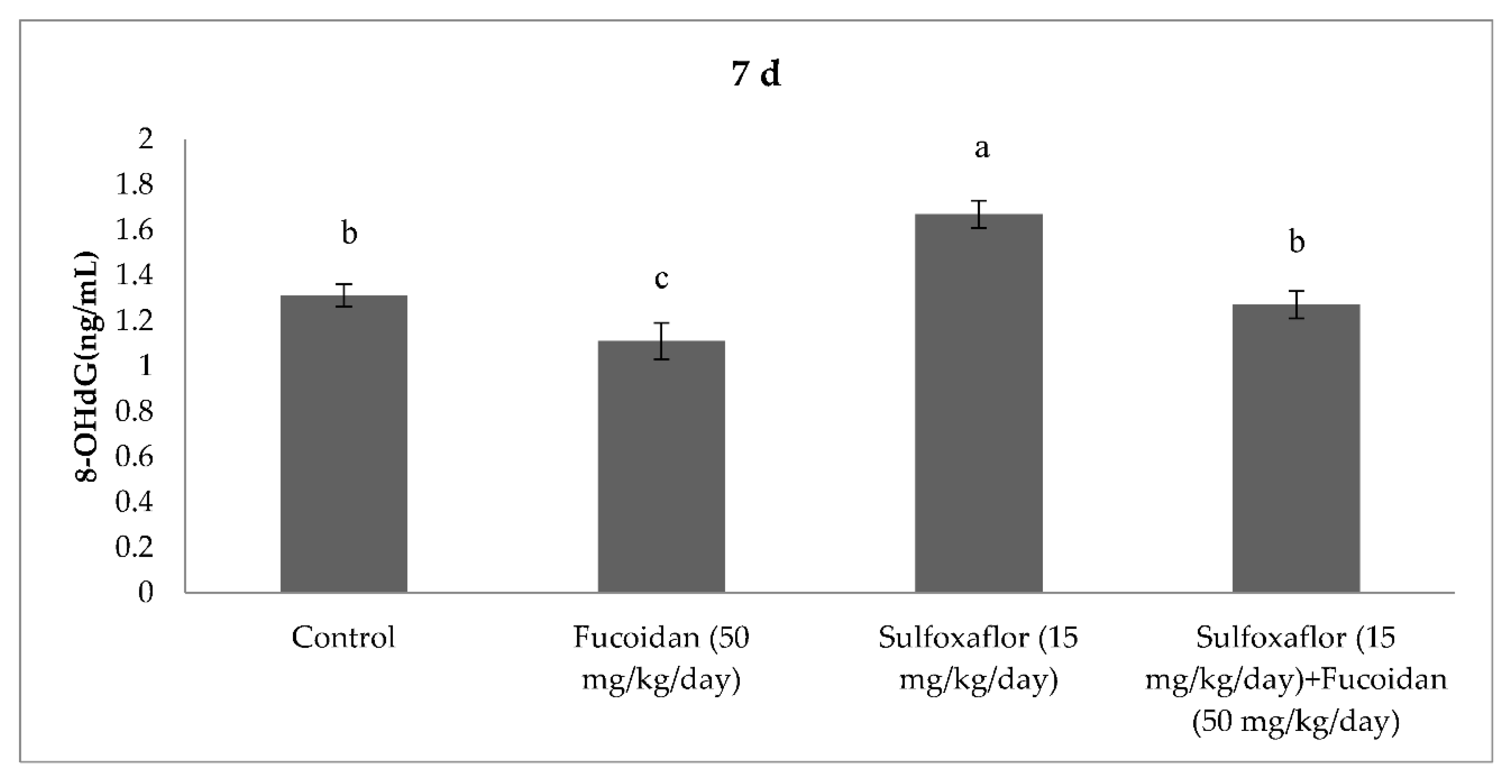
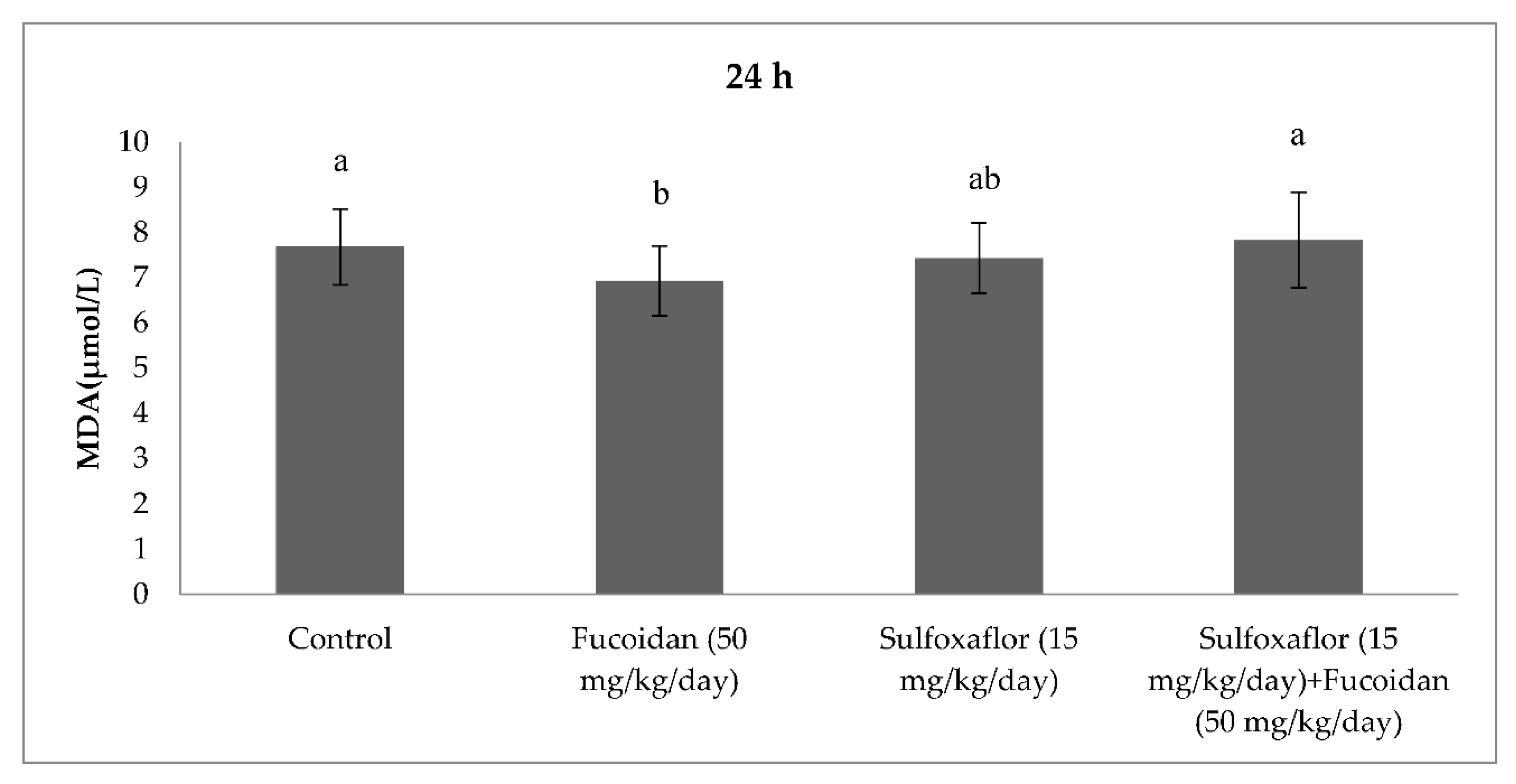
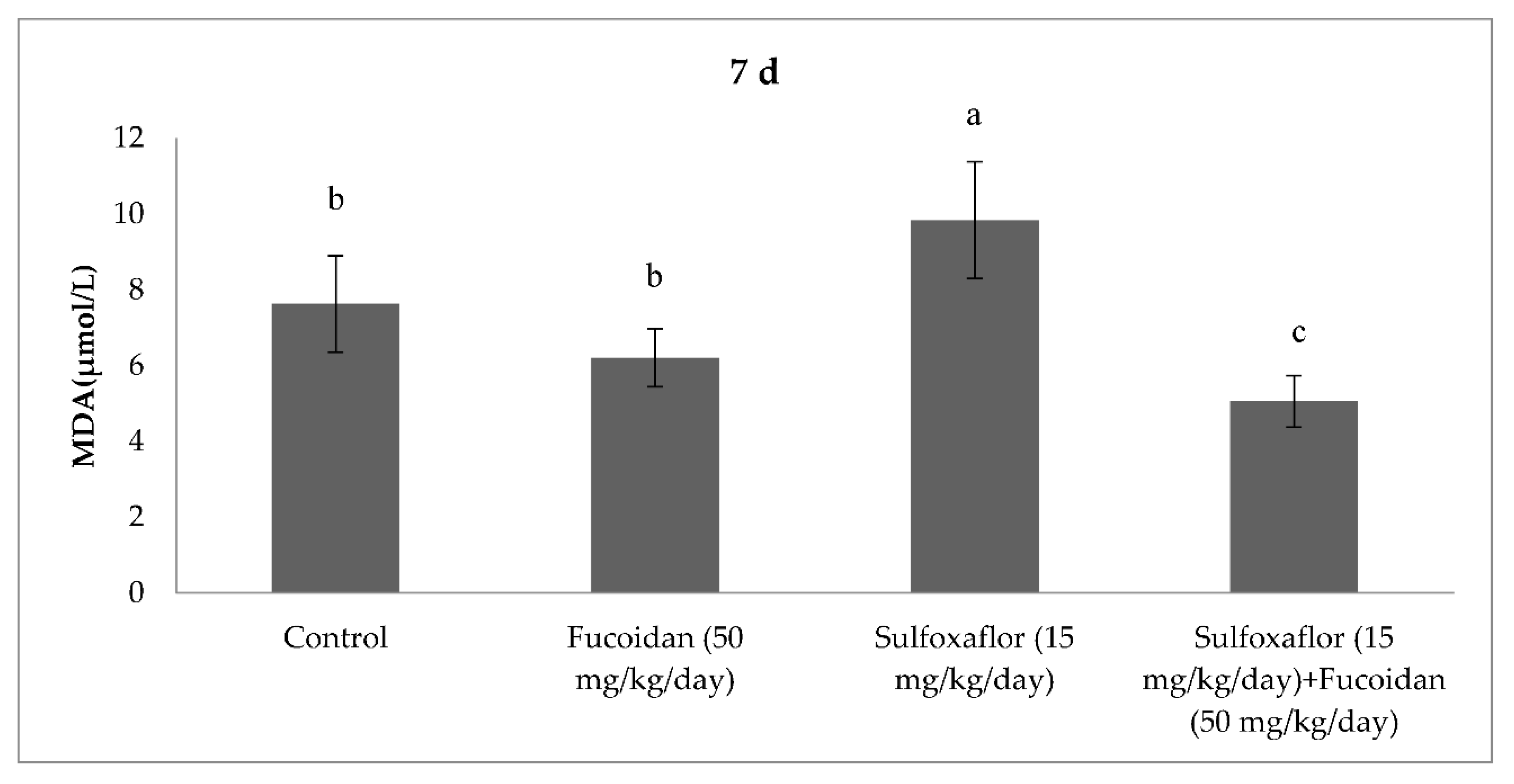
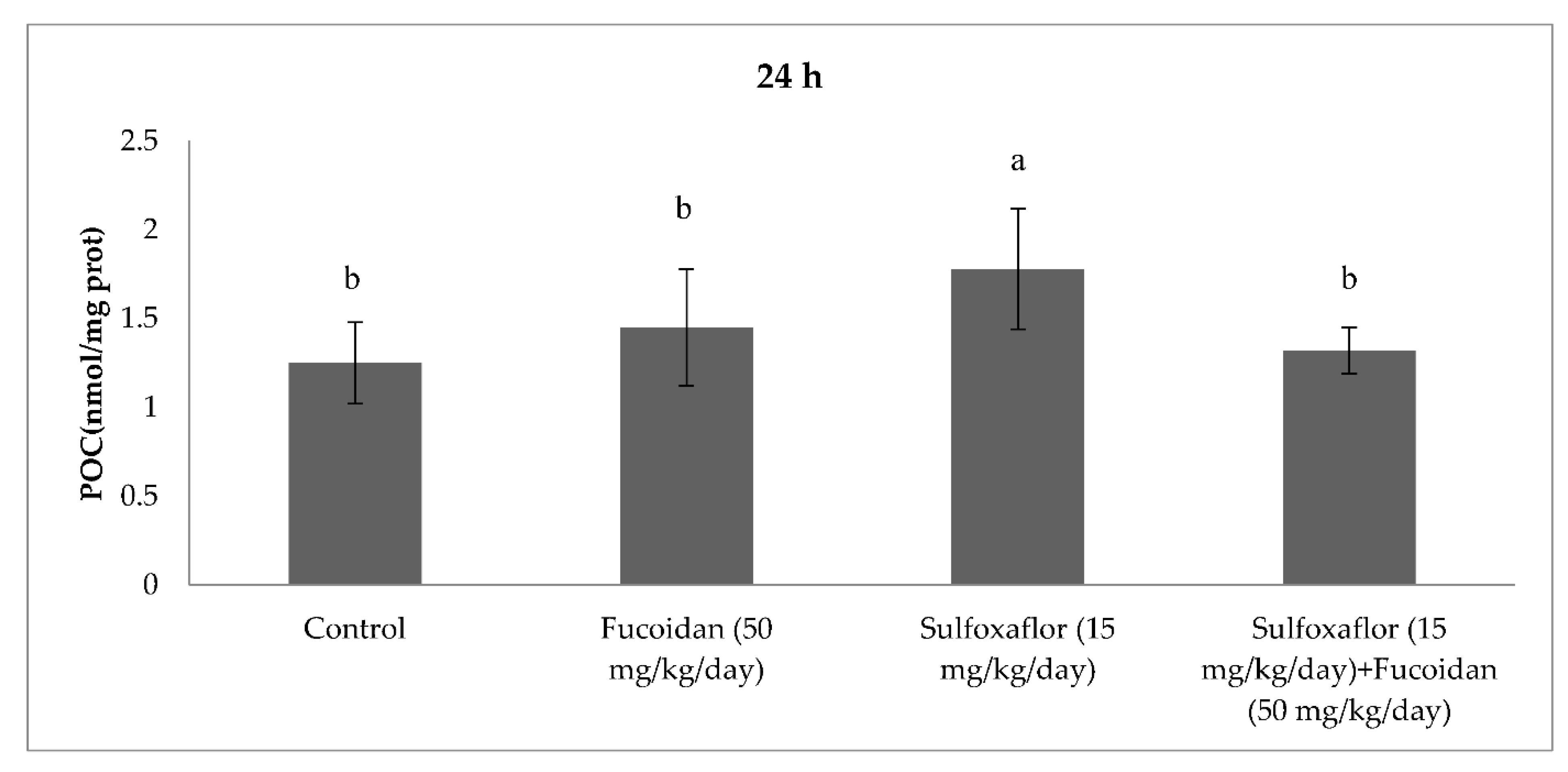
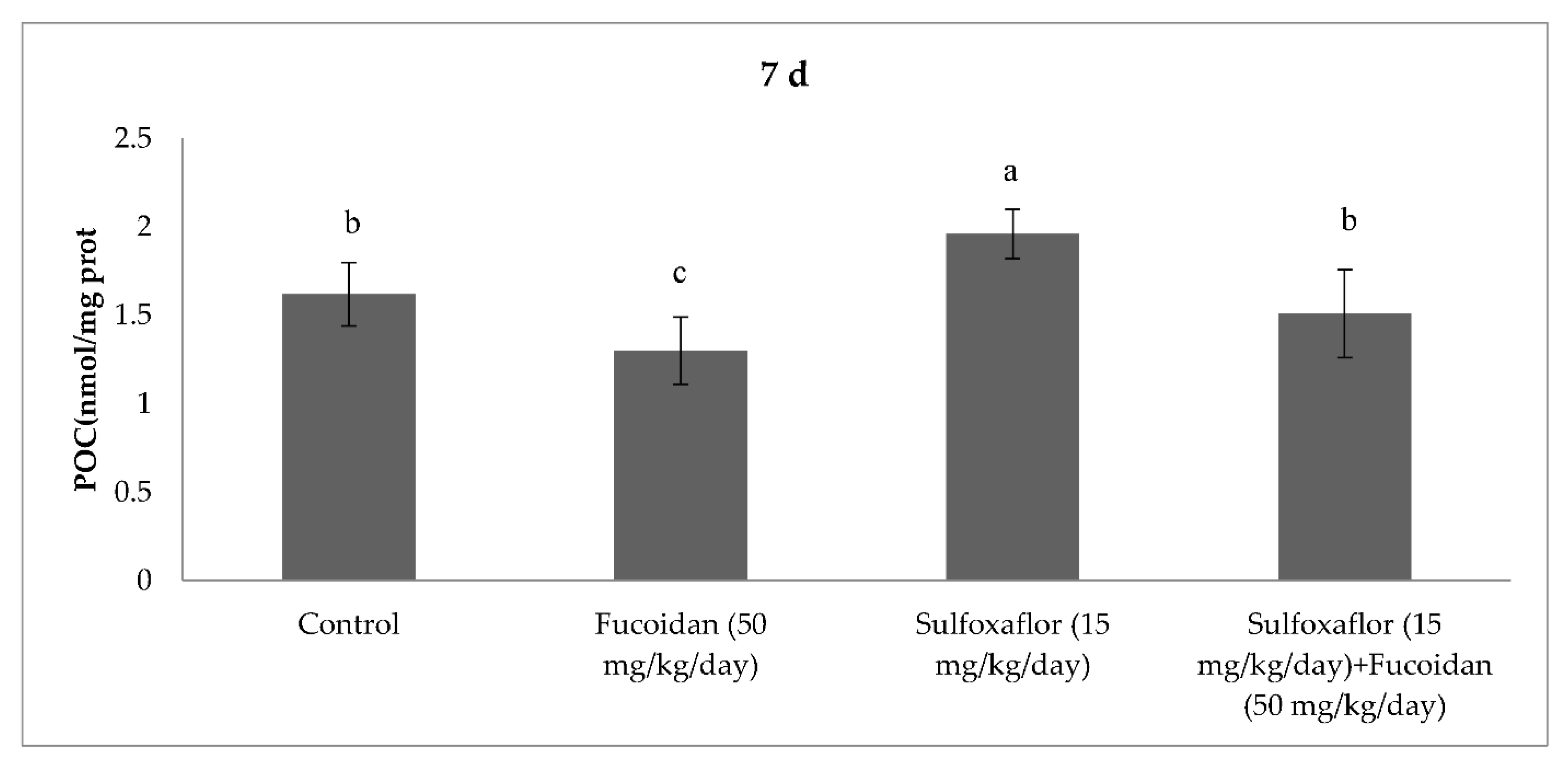
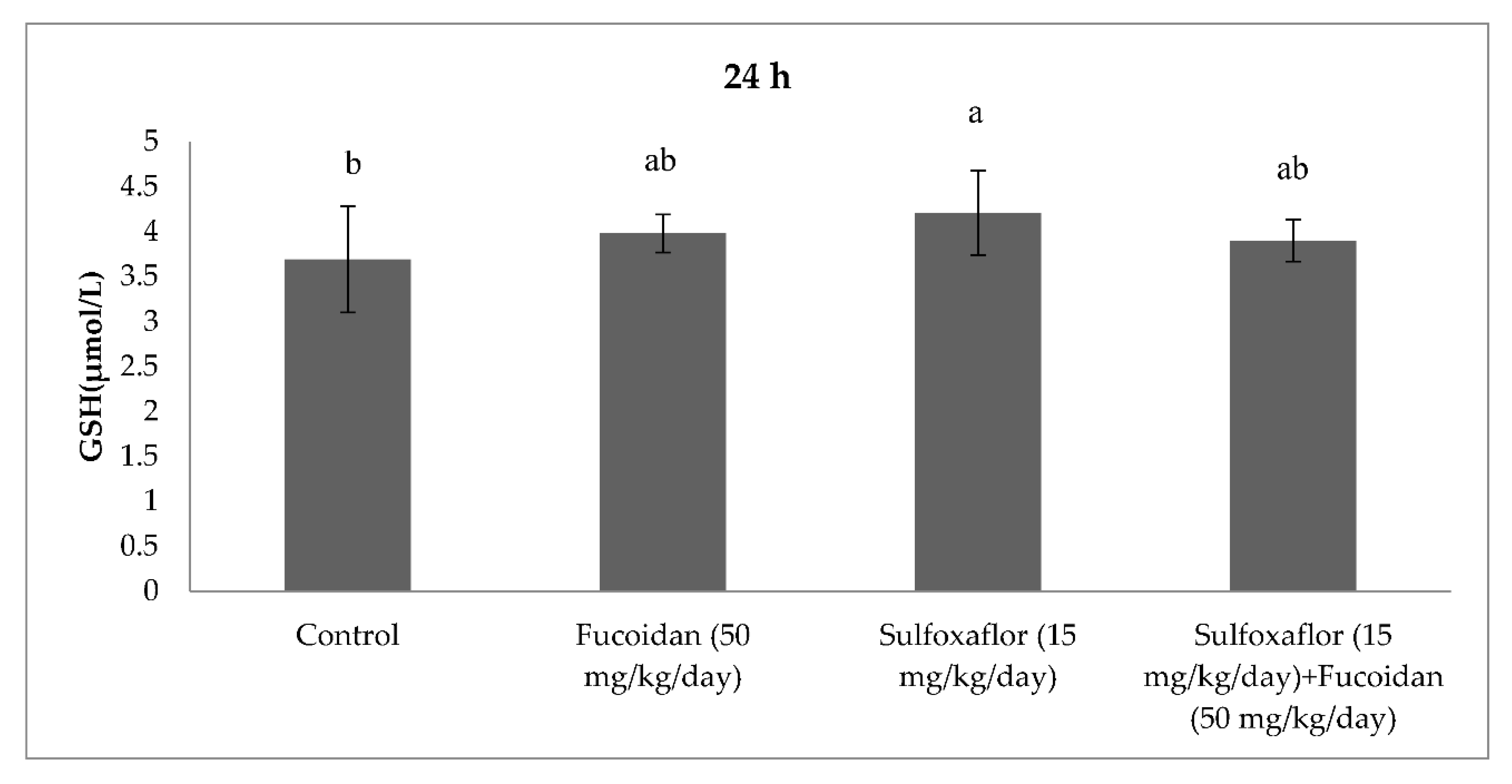
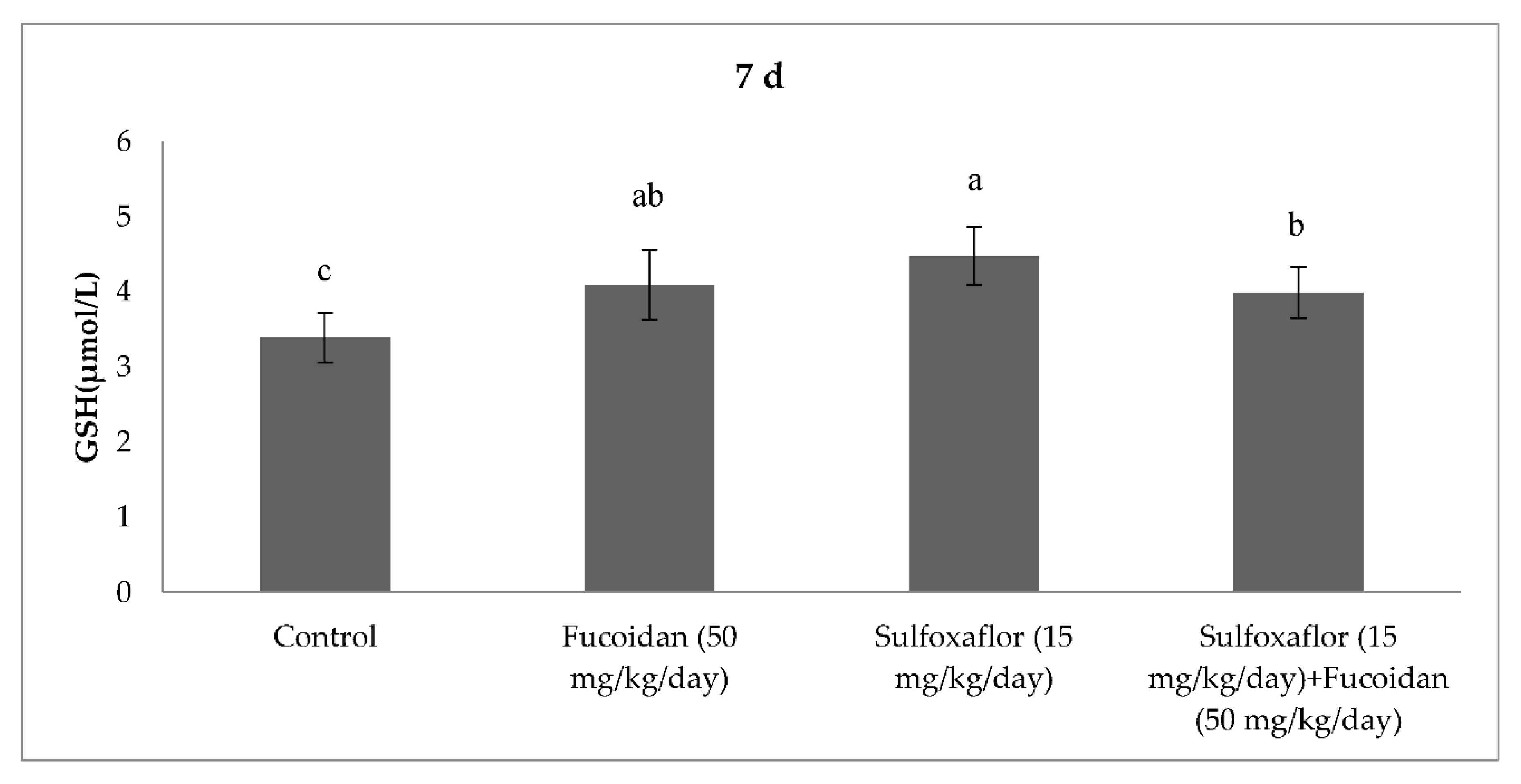
2.3. Changes in Serum Oxidative Stress and Antioxidant Markers
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals, Reagents, Kits
4.3. Experimental Design
4.4. Determination of Hematological Parameters
4.5. Determination of Biochemical Parameters in Serum
4.6. Determination of Oxidative Stress and Antioxidant Markers
4.7. Determination of Protein Levels
4.8. Statistical Analyzes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zymanczyk–Duda, E.; Szmigiel–Merena, B.; Brzezinska–Rodak, M. Natural antioxidants–properties and possible applications. J. Appl. Biotechnol. Bioeng. 2018, 5, 251–258. [Google Scholar]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The comparative analysis of antiviral activity of native and modified fucoidans from brown algae Fucus evanescens in vitro and in vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of bioactivities of fucoidan from the brown seaweed Fucus vesiculosus L. of the Barents sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.Y. Fucoidan as a marine anticancer agent in preclinical development. Mar. Drugs 2014, 12, 851–870. [Google Scholar] [CrossRef]
- Zorofchian Moghadamtousi, S.; Karimian, H.; Khanabdali, R.; Razavi, M.; Firoozinia, M.; Zandi, K.; Abdul Kadir, H. Anticancer and antitumor potential of fucoidan and fucoxanthin, two main metabolites isolated from brown algae. Sci. World J. 2014, 2014, 768323. [Google Scholar] [CrossRef]
- Luo, D.; Zhang, Q.; Wang, H.; Cui, Y.; Sun, Z.; Yang, J.; Zheng, Y.; Jia, J.; Yu, F.; Wang, X.; et al. Fucoidan protects against dopaminergic neuron death in vivo and in vitro. Eur. J. Pharmacol. 2009, 617, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Omar, H.E.D.M.; Eldien, H.M.S.; Badary, M.S.; Al–Khatib, B.Y.; Abd Elgaffar, S.K. The immunomodulating and antioxidant activity of fucoidan on the splenic tissue of rats treated with cyclosporine A. J. Basic Appl. Zool. 2013, 66, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.D.; Lee, S.R.; Kim, T.; Jang, S.A.; Kang, S.C.; Koo, H.J.; Sohn, E.; Bak, J.P.; Namkoong, S.; Kim, H.K.; et al. Fucoidan from Fucus vesiculosus protects against alcohol–induced liver damage by modulating inflammatory mediators in mice and HepG2 cells. Mar. Drugs 2015, 13, 1051–1067. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.W.; Lee, H.S.; Jung, K.H.; Lee, H.; Hong, S.S. Protective effect of fucoidan against acetaminophen–induced liver injury. Arch. Pharm. Res. 2012, 35, 1099–1105. [Google Scholar] [CrossRef]
- AlKahtane, A.A.; Abushouk, A.I.; Mohammed, E.T.; ALNasser, M.; Alarifi, S.; Ali, D.; Alessia, M.S.; Almeer, R.S.; AlBasher, G.; Alkahtani, S.; et al. Fucoidan alleviates microcystin-LR-induced hepatic, renal, and cardiac oxidative stress and inflammatory injuries in mice. Environ. Sci. Pollut. Res. Int. 2020, 27, 2935–2944. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Abushouk, A.I.; Bahbah, E.I.; Bungău, S.G.; Alyousif, M.S.; Aleya, L.; Alkahtani, S. Fucoidan protects against subacute diazinon-induced oxidative damage in cardiac, hepatic, and renal tissues. Environ. Sci. Pollut. Res. Int. 2020, 27, 11554–11564. [Google Scholar] [CrossRef] [PubMed]
- Mahgoub, H.A.; El–Adl, M.A.M.; Martyniuk, C.J. Fucoidan ameliorates acute and sub–chronic in vivo toxicity of the fungicide cholorothalonil in Oreochromis niloticus (Nile tilapia). Comp. Biochem. Phys. C 2021, 245, 109035. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Peana, M.; Maes, M.; Dadar, M.; Severin, B. The glutathione system in Parkinson’s disease and its progression. Neurosci. Biobehav. Rev. 2021, 120, 470–478. [Google Scholar] [CrossRef]
- Wang, X.; Anadón, A.; Wu, Q.; Qiao, F.; Ares, I.; Martínez-Larrañaga, M.R.; Yuan, Z.; Martínez, M.A. Mechanism of neonicotinoid toxicity: Impact on Oxidative stress and metabolism. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 471–507. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.B.; Mukhopadhayay, S.K.; Sar, T.K.; Ganguly, S. Acetamiprid induces toxicity in mice under experimental conditions with prominent effect on the hematobiochemical parameters. J. Drug Metab. Toxicol. 2012, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Chakroun, S.; Ezzi, L.; Grissa, I.; Kerkeni, E.; Neffati, F.; Bhouri, R.; Sallem, A.; Najjar, M.F.; Hassine, M.; Mehdi, M.; et al. Hematological, biochemical, and toxicopathic effects of subchronic acetamiprid toxicity in Wistar rats. Environ. Sci. Pollut. R 2016, 23, 25191–25199. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R.; Schindler, M.; Alfred, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef]
- Gibbons, D.; Morrissey, C.; Mineau, P. A review of the direct and indirect effects of neonicotinoids and fipronil on vertebrate wildlife. Environ. Sci. Pollut. R 2015, 22, 103–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El–Gendy, K.S.; Aly, N.M.; Mahmoud, F.H.; Kenawy, A.; El–Sebae, A.K.H. The role of vitamin C as antioxidant in protection of oxidative stress induced by imidacloprid. Food Chem. Toxicol. 2010, 48, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, U.; Srivastava, M.; Srivastava, L. Toxicological impact of technical imidacloprid on ovarian morphology, hormones and antioxidant enzymes in female rats. Food Chem. Toxicol. 2011, 49, 3086–3089. [Google Scholar] [CrossRef] [PubMed]
- Khaldoun–Oularbi, H.; Bouzid, N.; Boukreta, S.; Makhlouf, C.; Derriche, F.; Djennas, N. Thiamethoxam Actara® induced alterations in kidney liver cerebellum and hippocampus of male rats. J. Xenobiot. 2017, 7, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, K.J.; Santana, M.B.; Do Nascimento, J.L.; Picanço–Diniz, D.L.; Maués, L.A.; Santos, S.N.; Ferreira, V.M.; Alfonso, M.; Durán, R.; Faro, L.R. Behavioral and biochemical effects of neonicotinoid thiamethoxam on the cholinergic system in rats. Ecotoxicol. Environ. Saf. 2010, 73, 101–107. [Google Scholar] [CrossRef]
- Zhu, Y.; Loso, M.R.; Watson, G.B.; Sparks, T.C.; Rogers, R.B.; Huang, J.X.; Gerwick, B.C.; Babcock, J.M.; Kelley, D.; Hegde, V.B.; et al. Discovery and characterization of sulfoxaflor, a novel insecticide targeting sap-feeding pests. J. Agric. Food Chem. 2011, 59, 2950–2957. [Google Scholar] [CrossRef]
- LeBaron, M.J.; Geter, D.R.; Rasoulpour, R.J.; Gollapudi, B.B.; Thomas, J.; Murray, J.; Kan, H.L.; Wood, A.J.; Elcombe, C.; Vardy, A.; et al. An integrated approach for prospectively investigating a mode-of-action for rodent liver effects. Toxicol. Appl. Pharmacol. 2013, 270, 164–173. [Google Scholar] [CrossRef]
- Rasoulpour, R.J.; Terry, C.; LeBaron, M.J.; Stebbins, K.; Ellis-Hutchings, R.G.; Billington, R. Mode-of-action and human relevance framework analysis for rat Leydig cell tumors associated with sulfoxaflor. Crit. Rev. Toxicol. 2014, 2, 25–44. [Google Scholar] [CrossRef]
- Rasoulpour, R.J.; Ellis-Hutchings, R.G.; Terry, C.; Millar, N.S.; Zablotny, C.L.; Gibb, A.V.; Marshall, T.; Collins, E.W.; Carney, E.W.; Billington, R. A novel mode-of-action mediated by the fetal muscle nicotinic acetylcholine receptor resulting in developmental toxicity in rats. Toxicol. Sci. 2012, 127, 522–534. [Google Scholar] [CrossRef] [Green Version]
- Piner Benli, P.; Celik, M. Glutathione and its dependent enzymes’ modulatory responses to neonicotinoid insecticide sulfoxaflor-induced oxidative damage in zebrafish in vivo. Sci. Prog. 2021, 104, 36850421102836. [Google Scholar] [CrossRef]
- Piner Benli, P.; Kaya, M.; Coskun, C. Fucoidan modulated oxidative stress and caspase-3 mRNA expression induced by sulfoxaflor in the brain of mice. Neurotox. Res. 2021, 39, 1908–1919. [Google Scholar] [CrossRef]
- Kataria, S.K.; Chhillar, A.K.; Kumar, A.; Tomar, M.; Malik, V. Cytogenetic and hematological alterations induced by acute oral exposure of imidacloprid in female mice. Drug Chem. Toxicol. 2016, 39, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, Q.; Song, J. Toxicological evaluation of fucoidan extracted from Laminaria japonica in Wistar rats. Food Chem. Toxicol. 2005, 43, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Phull, A.R.; Majid, M.; Haq, I.U.; Khan, M.R.; Kim, S.J. In vitro and in vivo evaluation of anti-arthritic, antioxidant efficacy of fucoidan from Undaria pinnatifida (Harvey) Suringar. Int. J. Biol. Macromol. 2017, 97, 468–480. [Google Scholar] [CrossRef]
- Ramu, S.; Murali, A.; Narasimhaiah, G.; Jayaraman, A. Toxicological evaluation of Sargassum wightii greville derived fucoidan in wistar rats: Hematological, biochemical and histopathological evidences. Toxicol. Rep. 2020, 23, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Jeun, J.; Houng, S.J.; Jun, H.J.; Kweon, D.K.; Lee, S.J. Toxicological evaluation of fucoidan from Undaria pinnatifidain vitro and in vivo. Phytother. Res. 2010, 24, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Murray, J.A.; Saghir, S.A.; Yano, B.L. Xr-208: 90-Day Dietary Toxicity Study in CD-1 Mice; Report of Toxicology & Environmental Research and Consulting; The Dow Chemical Company: Midland, MI, USA, 2010. [Google Scholar]
- Thomas, J.; Dryzga, M.D.; Saghir, S.A.; McClymont, E.L.; Quast, J.F. Xr-208: 4-Week Repeated Dose Dietary Toxicity Study in crl: CD1(icr) Mice; Report of Toxicology & Environmental Research and Consulting; The Dow Chemical Company: Midland, MI, USA, 2008. [Google Scholar]
- Bhardwaj, S.; Srivastava, M.K.; Kapoor, U.; Srivastava, L.P. A 90 days oral toxicity of imidacloprid in female rats: Morphological, biochemical and histopathological evaluations. Food Chem. Toxicol. 2010, 48, 1185–1190. [Google Scholar] [CrossRef]
- Zhang, J.J.; Wang, Y.; Xiang, H.Y.; Li, M.X.; Li, W.H.; Ma, K.G.; Wang, X.Z.; Zhang, J.H. Oxidative stress: Role in acetamiprid-induced impairment of the male mice reproductive system. Agric. Sci. China. 2011, 10, 786–796. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abdeen, A.; Jalouli, M.; Abdelkader, A.; Megahed, A.; Alkahtane, A.; Almeer, R.; Alhoshani, N.M.; Al-Johani, N.S.; Alkahtani, S.; et al. Fucoidan supplementation modulates hepato-renal oxidative stress and DNA damage induced by aflatoxin B1 intoxication in rats. Sci. Total Environ. 2021, 768, 144781. [Google Scholar] [CrossRef]
- Aleissa, M.S.; Alkahtani, S.; Abd Eldaim, M.A.; Ahmed, A.M.; Bungău, S.G.; Almutairi, B.; Bin-Jumah, M.; AlKahtane, A.A.; Alyousif, M.S.; Abdel-Daim, M.M. Fucoidan ameliorates oxidative stress, inflammation, DNA Damage, and hepatorenal injuries in diabetic rats intoxicated with aflatoxin B1. Oxidative Med. Cell. Longev. 2020, 2020, 9316751. [Google Scholar] [CrossRef] [Green Version]
- Topal, A.; Alak, G.; Ozkaraca, M.; Yeltekin, A.C.; Comaklı, S.; Acıl, G.; Kokturk, M.; Atamanalp, M. Neurotoxic responses in brain tissues of rainbow trout exposed to imidacloprid pesticide: Assessment of 8–hydroxy–2–deoxyguanosine activity, oxidative stress and acetylcholinesterase activity. Chemosphere 2017, 175, 186–191. [Google Scholar] [CrossRef]
- Mohamed, A.A.R.; Mohamed, W.A.M.; Khater, S.I. Imidacloprid induces various toxicological effects related to the expression of 3 beta-hsd, nr5a1, and ogg1 genes in mature and immature rats. Environ. Pollut. 2017, 221, 15–25. [Google Scholar] [CrossRef]
- Mishchuk, O.V.; Stoliar, O.B. The effect of pesticide acetamiprid on biochemical markers in tissues of fresh water bivalve mussels anodonta cygnea l. (unionidae). Ukr. Biochem. J. 2008, 80, 117–124. [Google Scholar]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological activities of fucoidan and the factors mediating its therapeutic effects: A review of recent studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [Green Version]
- Fitton, J.H.; Stringer, D.N.; Park, A.Y.; Karpiniec, S.S. Therapies from fucoidan: New developments. Mar. Drugs 2019, 17, 571. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, M.; Allahgholi, L.; Sardari, R.R.; Hreggviðsson, G.O.; Nordberg Karlsson, E. Extraction and modification of macroalgal polysaccharides for current and next-generation applications. Molecules 2020, 25, 930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, H.R.; Biller, P.; Ross, A.B.; Adams, J.M.M. The seasonal variation of fucoidan within three species of brown macroalgae. Algal Res. 2017, 22, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Usov, A.I.; Bilan, M.I. Fucoidans—Sulfated polysaccharides of brown algae. Russ. Chem. Rev. 2009, 78, 785–799. [Google Scholar] [CrossRef]
- Zhao, X.; Xue, C.H.; Cai, Y.P.; Wang, D.F.; Fang, Y. The study of antioxidant activities of fucoidan from Laminaria japonica. High Technol. Lett. 2005, 11, 91–94. [Google Scholar]
- Li, L.H.; Xue, C.H.; Xue, Y.; Li, Z.J.; Fu, X.Y. The effects of fucoidans from Laminaria japonica on AAPH mediated oxidation of human low-density lipoprotein. Acta Oceanol. Sin. 2006, 25, 124–130. [Google Scholar]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef]
- Micheline, R.S.; Cybelle, M.; Celina, G.D.; Fernando, F.S.; Hugo, O.R.; Edda, L. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J. Appl. Phycol. 2007, 19, 153–160. [Google Scholar]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and tissue distribution of fucoidan from Fucus vesiculosus after oral administration to rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [Green Version]
- Lean, Q.Y.; Eri, R.D.; Fitton, J.H.; Patel, R.P.; Gueven, N. Fucoidan extracts ameliorate acute colitis. PLoS ONE 2015, 10, e0128453. [Google Scholar] [CrossRef] [Green Version]
- Richards, C.; Williams, N.A.; Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S.; Park, A.Y. Oral fucoidan attenuates lung pathology and clinical signs in a severe influenza a mouse model. Mar. Drugs 2020, 18, 246. [Google Scholar] [CrossRef]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A quantitative method to detect fucoidan in human plasma using a novel antibody. Methods Find Exp. Clin. 2005, 27, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Tokita, Y.; Nakajima, K.; Mochida, H.; Iha, M.; Nagamine, T. Development of a fucoidan-specific antibody and measurement of fucoidan in serum and urine by sandwich ELISA. Biosci. Biotechnol. Biochem. 2010, 74, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ordóñez, E.; Jiménez-Escrig, A.; Rupérez, P. Molecular weight distribution of polysaccharides from edible seaweeds by high-performance size-exclusion chromatography (HPSEC). Talanta 2012, 93, 153–159. [Google Scholar] [CrossRef] [Green Version]
- OECD. Guideline for Testing of Chemicals: Acute Oral Toxicity, Acute Toxic Class Method; OECD: Paris, France, 2001. [Google Scholar]
- Brooks, K.J.; Wiescinski, C.M.; Golden, R. MXDE-208: Acute Oral Toxicity Study in CRL: CD1 (ICR) Mice (Up and Down Procedure); DAS Report No. 081059 Submitted to WHO by Dow AgroSciences Europe, Abingdon, UK; The Dow Chemical Company: Midland, MI, USA, 2008. [Google Scholar]
- Li, D.Y.; Xu, R.Y.; Zhou, W.Z.; Sheng, X.B.; Yang, A.Y.; Cheng, J.L. Effects of fucoidan extracted from brown seaweed on lipid peroxidation in mice. Acta Nutr. Sin. 2002, 24, 389–392. [Google Scholar]
- Ponnan, A.; Kulanthaiyesu, A.; Marudhamuthu, M.; Palanisamy, K.; Kadarkarai, M. Protective effects of fucoidan against 4-nitroquinolin-1-oxide provoked genetic damage in mouse bone marrow cells. Environ. Sci. Pollut. Res. Int. 2020, 27, 31760–31766. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24-h | Control | Fucoidan (50 mg/kg/day) | Sulfoxaflor (15 mg/kg/day) | Sulfoxaflor (15 mg/kg/day) + Fucoidan (50 mg/kg/day) |
|---|---|---|---|---|
| RBC (1012/L) | 7.01 ± 1.01 a | 6.69 ± 0.49 a | 7.43 ± 0.73 a | 7.31 ± 0.81 a |
| HGB (g/dL) | 10.56 ± 1.49 ab | 10.51 ± 0.27 b | 11.32 ± 0.81 a | 10.98 ± 0.06 a |
| HCT | 0.35 ± 0.05 a | 0.34 ± 0.02 a | 0.38 ± 0.03 a | 0.36 ± 0.04 a |
| MCV (fL) | 49.98 ± 2.16 a | 50.31 ± 3.25 a | 52.15 ± 4.6 a | 50.56 ± 2.17 a |
| MCH (pg) | 15.06 ± 0.74 a | 15.11 ± 0.8 a | 15.28 ± 1.16 a | 15.05 ± 0.77 a |
| MCHC (g/dL) | 30.17 ± 0.7 a | 30.09 ± 0.74 a | 29 ± 0.09 b | 29.73 ± 0.69 ab |
| Plt (109/L) | 633.62 ± 142.71 ab | 473.93 ± 204.97 b | 677.31 ± 197.2 a | 675.93 ± 140.58 a |
| WBC (109/L) | 6.49 ± 0.26 a | 6.07 ± 0.33 ab | 5.41 ± 1.11 b | 6.46 ± 0.65 a |
| Neu (%) | 0.393. ± 0.10 a | 0.427 ± 0.16 a | 0.306 ± 0.08 a | 0.364 ± 0.11 a |
| Lym (%) | 0.538 ± 0.12 a | 0.499 ± 0.18 a | 0.618 ± 0.11 a | 0.891 ± 0.08 a |
| Mon (%) | 0.048 ± 0.016 a | 0.037 ± 0.15 a | 0.052 ± 0.035 a | 0.034 ± 0.02 a |
| Eos (%) | 0.0187 ± 0.012 a | 0.0194 ± 0.039 a | 0.0215 ± 0.011 b | 0.0277 ± 0.008 b |
| 7-d | Control | Fucoidan (50 mg/kg/day) | Sulfoxaflor (15 mg/kg/day) | Sulfoxaflor (15 mg/kg/day) + Fucoidan (50 mg/kg/day) |
|---|---|---|---|---|
| RBC (1012/L) | 7.41 ± 0.048 a | 7.31 ± 0.32 a | 7.65 ± 0.29 a | 9.14 ± 0.02 a |
| HGB (g/dL) | 11.68 ± 0.7 c | 11.0 ± 0.38 ab | 12.12 ± 0.5 b | 14.68 ± 1.35 a |
| HCT | 0.38 ± 0.02 b | 0.37 ± 0.03 ab | 0.41 ± 0.02 b | 0.50 ± 0.04 a |
| MCV (fL) | 51.91 ± 0.64 ab | 46.92 ± 4.64 a | 51.21 ± 0.68 b | 54.77 ± 2.01 a |
| MCH (pg) | 15.68 ± 0.24 a | 16.95 ± 1.18 a | 15.83 ± 0.28 a | 15.93 ± 0.52 a |
| MCHC (g/dL) | 30.25 ± 0.38 a | 30.23 ± 0.23 a | 29.22 ± 0.21 b | 29.15 ± 0.15 b |
| Plt (109/L) | 583.93 ± 200.97 a | 522.31 ± 136.94 a | 626.62 ± 142.67 a | 628.25 ± 146.06 a |
| WBC (109/L) | 6.50 ± 0.3 a | 6.37 ± 0.3 a | 4.38 ± 0.26 b | 5.88 ± 0.35 a |
| Neu (%) | 0.438 ± 0.115 a | 0.389 ± 0.141 a | 0.303 ± 0.132 a | 0.411 ± 0.134 a |
| Lym (%) | 0.489 ± 0.135 a | 0.550 ± 0.165 a | 0.605 ± 0.128 a | 0.467 ± 0.124 a |
| Mon (%) | 0.053 ± 0.031 b | 0.044 ± 0.023 b | 0.076 ± 0.047 ab | 0.105 ± 0.063 a |
| Eos (%) | 0.0183 ± 0.005 ab | 0.0176 ± 0.005 ab | 0.0131 ± 0.006 b | 0.0242 ± 0.012 a |
| 24-h | Control | Fucoidan (50 mg/kg/day) | Sulfoxaflor (15 mg/kg/day) | Sulfoxaflor (15 mg/kg/day) + Fucoidan (50 mg/kg/day) |
|---|---|---|---|---|
| AST (U/L) | 50.25 ± 6.71 b | 53 ± 5.12 b | 66.25 ± 8.03 a | 55.75 ± 6.36 b |
| ALT) (U/L) | 69 ± 11.1 a | 68.75 ± 6.18 a | 74.87 ± 12.59 a | 68.25 ± 9.03 a |
| GGT (U/L) | 13.06 ± 0.80 a | 13.68 ± 0.54 a | 14.87 ± 12.59 a | 13.55 ± 9.03 a |
| LDH (U/L) | 516.12 ± 115.83 b | 532.25 ± 142.59 b | 1171.12 ± 361.65 a | 439.87 ± 149.82 b |
| Cre (mg/dL) | 0.49 ± 0.047 a | 0.51 ± 0.09 a | 0.53 ± 0.063 a | 0.50 ± 0.03 a |
| BUN (mg/dL) | 57.5 ± 7.75 a | 56.25 ± 5.99 a | 57.12 ± 12.4 a | 53.12 ± 3.97 a |
| TBil (mg/dL) | 0.78 ± 0.31 a | 0.94 ± 0.72 a | 0.71 ± 0.15 a | 0.71 ± 0.62 a |
| 7-d | ||||
| AST (U/L) | 53.25 ± 8.13 a | 37.87 ± 4.51 b | 40.01 ± 4.79 ab | 48.5 ± 7.38 a |
| ALT (U/L) | 73.5 ± 12.31 a | 62.25 ± 4.46 a | 68.25 ± 23.33 a | 76.5 ± 11.45 a |
| GGT (U/L) | 15 ± 2.65 a | 14.75 ± 2.19 a | 14.92 ± 2.29 a | 15.03 ± 1.77 a |
| LDH (U/L) | 442.5 ± 162.43 a | 204.75 ± 38.27 b | 322.12 ± 234.38 ab | 492 ± 276.16 a |
| Cre (mg/dL) | 0.49 ± 0.05 a | 0.47 ± 0.04 a | 0.50 ± 0.04 a | 0.49 ± 0.03 a |
| BUN (mg/dL) | 60.08 ± 5.59 a | 51 ± 1.51 b | 57.2 ± 11.04 ab | 55.6 ± 6.06 ab |
| TBil (mg/dL) | 0.9 ± 0.27 a | 0.67 ± 0.12 b | 0.77 ± 0.19 ab | 0.66 ± 0.11 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piner Benli, P.; Kaya, M.; Dağlıoğlu, Y.K. Fucoidan Protects against Acute Sulfoxaflor-Induced Hematological/Biochemical Alterations and Oxidative Stress in Male Mice. Pharmaceuticals 2022, 15, 16. https://doi.org/10.3390/ph15010016
Piner Benli P, Kaya M, Dağlıoğlu YK. Fucoidan Protects against Acute Sulfoxaflor-Induced Hematological/Biochemical Alterations and Oxidative Stress in Male Mice. Pharmaceuticals. 2022; 15(1):16. https://doi.org/10.3390/ph15010016
Chicago/Turabian StylePiner Benli, Petek, Merve Kaya, and Yusuf Kenan Dağlıoğlu. 2022. "Fucoidan Protects against Acute Sulfoxaflor-Induced Hematological/Biochemical Alterations and Oxidative Stress in Male Mice" Pharmaceuticals 15, no. 1: 16. https://doi.org/10.3390/ph15010016
APA StylePiner Benli, P., Kaya, M., & Dağlıoğlu, Y. K. (2022). Fucoidan Protects against Acute Sulfoxaflor-Induced Hematological/Biochemical Alterations and Oxidative Stress in Male Mice. Pharmaceuticals, 15(1), 16. https://doi.org/10.3390/ph15010016