
The Statin Target HMG-Coenzyme a Reductase (Hmgcr) Regulates Sleep Homeostasis in Drosophila


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
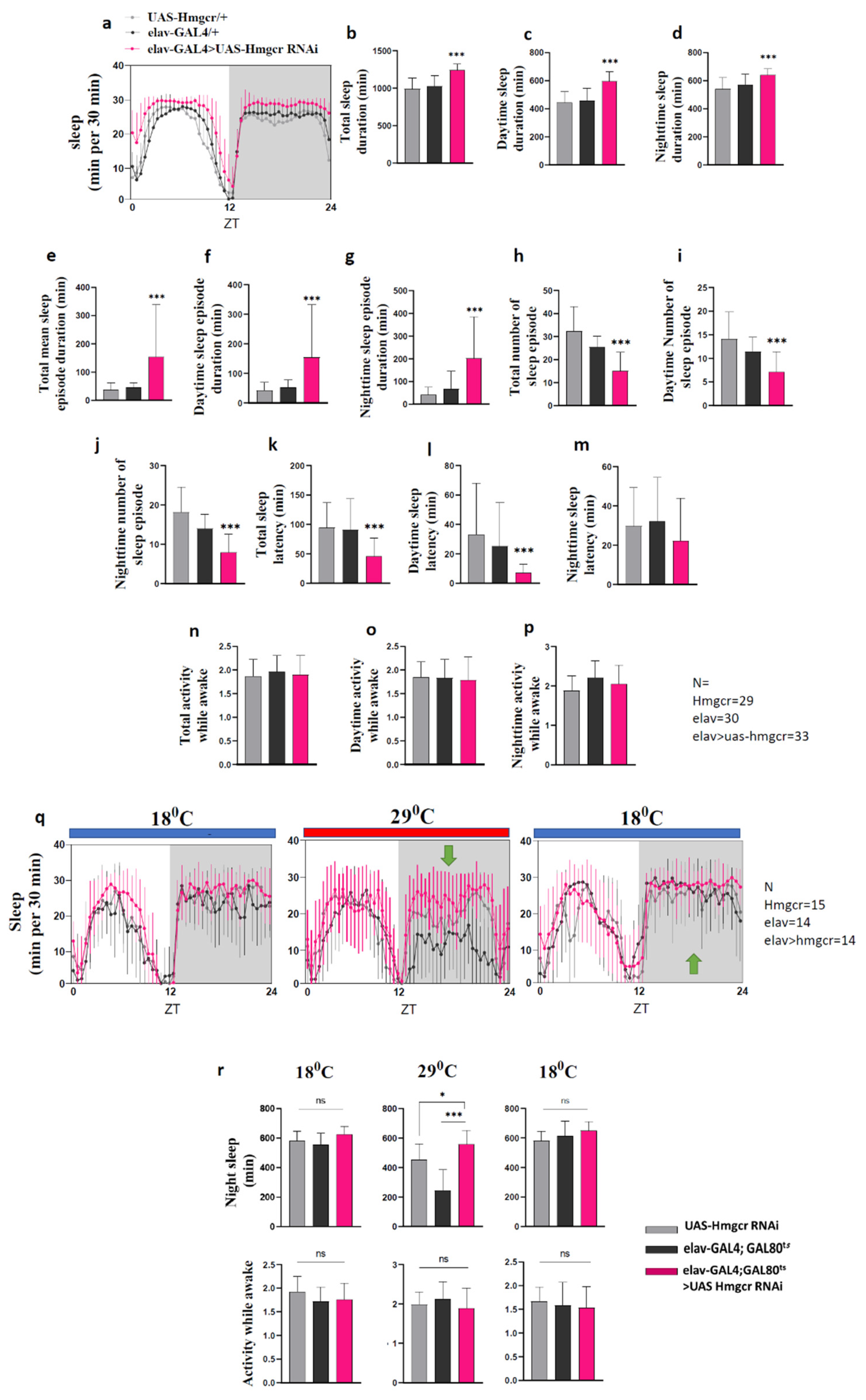
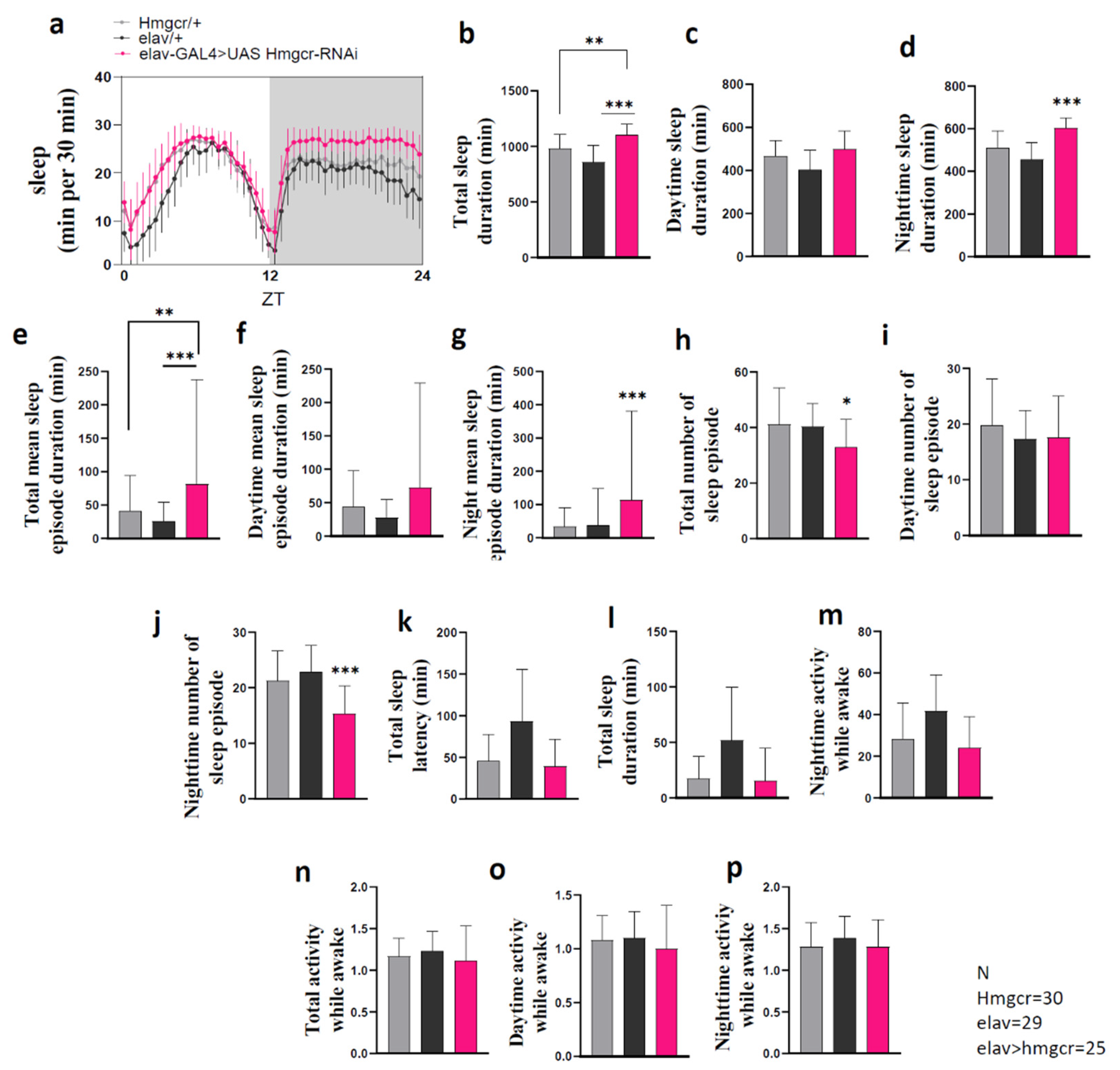
2.1. Down-Regulation of Hmgcr within Neurons Promotes Sleep
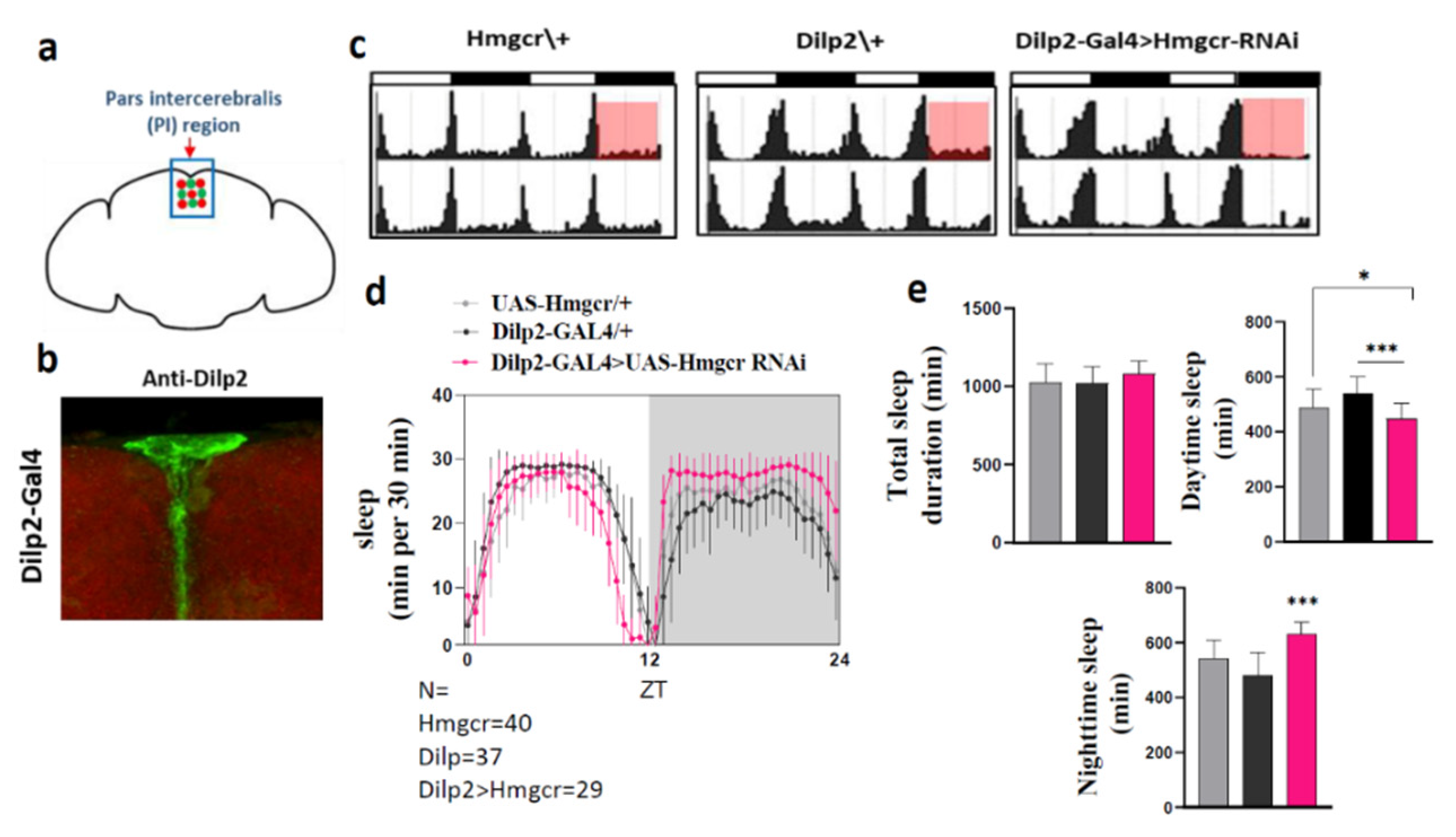
2.2. Loss of Hmgcr in the Pars Intercerebralis Insulin-Producing Neurons Recapitulates the Pan-Neuronal Phenotype
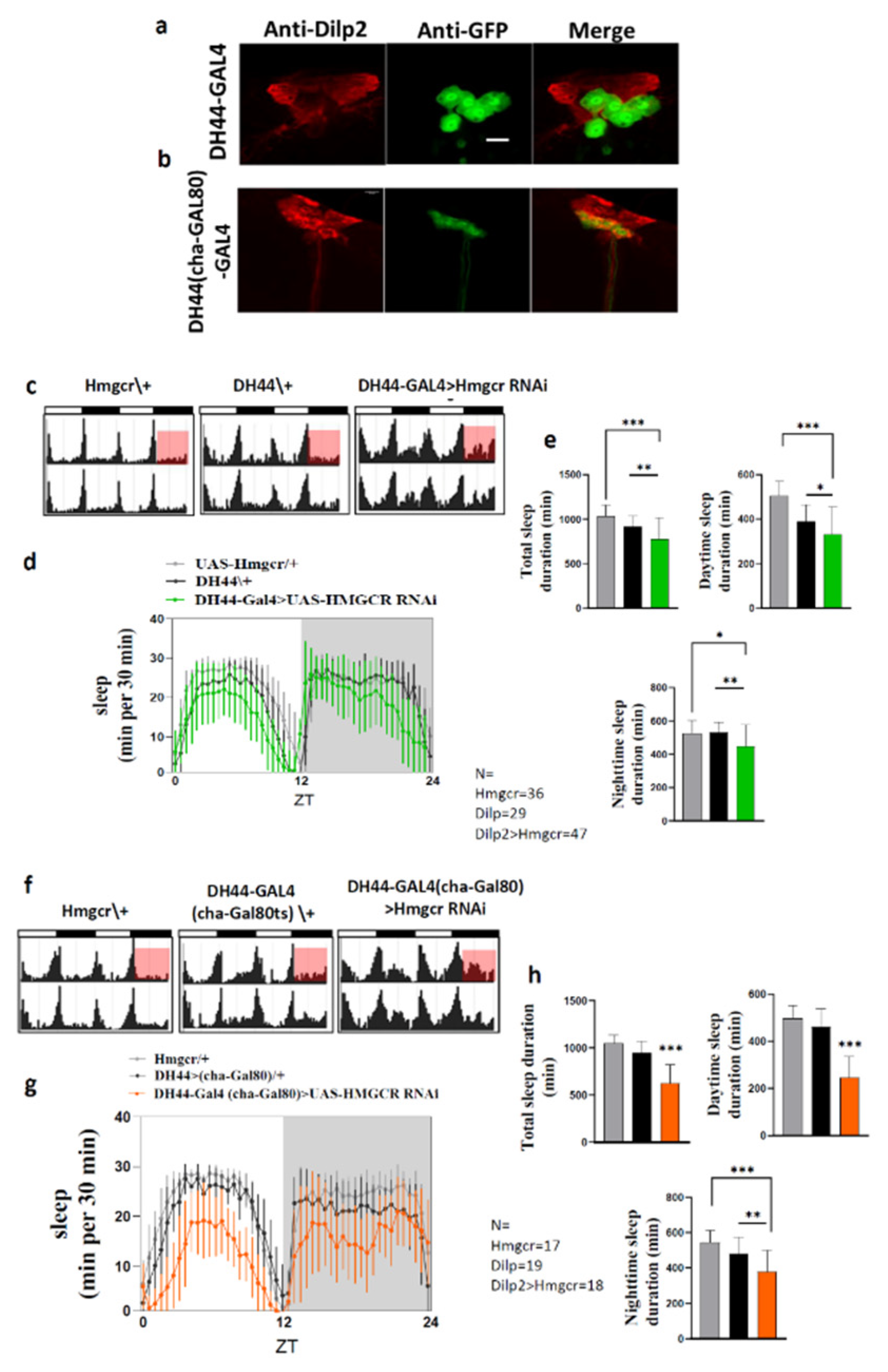
2.3. Hmgcr Is Also Required in Pars Intercerebralis DH44 Neurons for Proper Sleep Regulation
2.4. Hmgcr Regulates Sleep through Homeostatic Pathways
3. Discussion
4. Materials and Methods
4.1. Fly Stocks and Maintenance
4.2. Locomotor Activity Assay
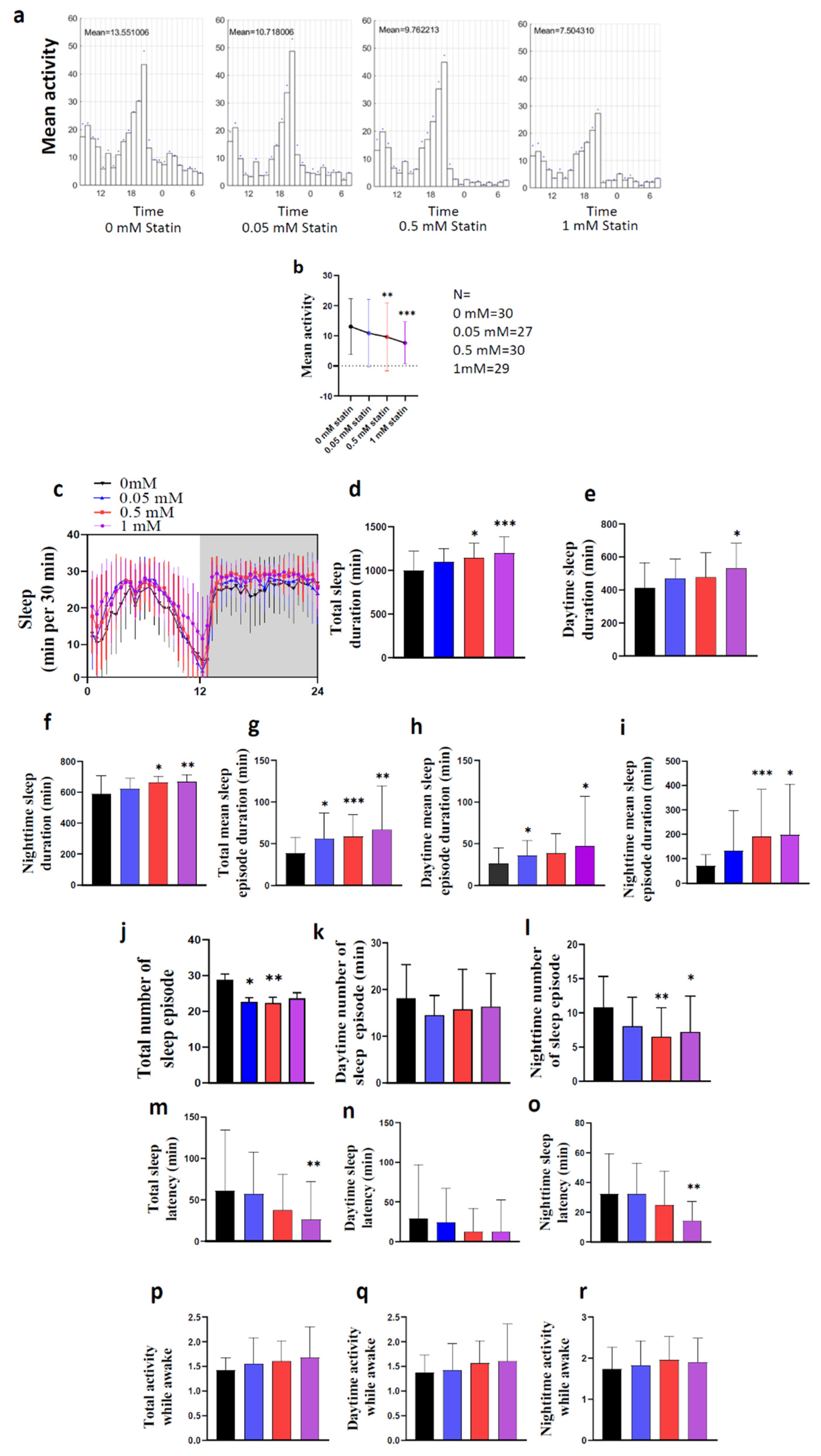
4.3. Statin Experiments
4.4. Immunostaining
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Regan, C.; Wu, P.; Arora, P.; Perri, D.; Mills, E.J. Statin Therapy in Stroke Prevention: A Meta-analysis Involving 121,000 Patients. Am. J. Med. 2008, 121, 24–33. [Google Scholar] [CrossRef]
- Takada, M.; Fujimoto, M.; Yamazaki, K.; Takamoto, M.; Hosomi, K. Association of statin use with sleep disturbances: Data mining of a spontaneous reporting database and a prescription database. Drug Saf. 2014, 37, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Vgontzas, A.N.; Kales, A.; Bixler, E.O.; Manfredi, R.L.; Tyson, K.L. Effects of lovastatin and pravastatin on sleep efficiency and sleep stages. Clin. Pharmacol. Ther. 1991, 50, 730–737. [Google Scholar] [CrossRef]
- Barth, J.D.; A Kruisbrink, O.; Van Dijk, A.L. Inhibitors of hydroxymethylglutaryl co-enzyme A reductase for treating hypercholesterolaemia. Br. Med. J. 1990, 301, 669. [Google Scholar] [CrossRef] [Green Version]
- MHRA Public Assessment Report. Statins: Updates to Product Safety Information, 2009. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/852414/Statins_update_to_product_safety_information.pdf (accessed on 11 January 2021).
- Broncel, M.; Gorzelak-Pabiś, P.; Sahebkar, A.; Serejko, K.; Ursoniu, S.; Rysz, J.; Serban, M.C.; Możdżan, M.; Banach, M. Sleep changes following statin therapy: A systematic review and meta-analysis of randomized placebo-controlled polysomnographic trials. Arch. Med. Sci. 2015, 11, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Joiner, W.J.; Crocker, A.; White, B.H.; Sehgal, A. Sleep in Drosophila is regulated by adult mushroom bodies. Nature 2006, 441, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Ganguly-Fitzgerald, I.; Donlea, J.; Shaw, P.J. Waking experience affects sleep need in Drosophila. Science 2006, 313, 1775–1781. [Google Scholar] [CrossRef]
- Cirelli, C.; Bushey, D.; Hill, S.; Huber, R.; Kreber, R.; Ganetzky, B.; Tononi, G. Reduced sleep in Drosophila Shaker mutants. Nature 2005, 434, 1087–1092. [Google Scholar] [CrossRef]
- Bushey, D.; Huber, R.; Tononi, G.; Cirelli, C. Drosophila Hyperkinetic mutants have reduced sleep and impaired memory. J. Neurosci. 2007, 27, 5384–5393. [Google Scholar] [CrossRef]
- Donlea, J.M.; Thimgan, M.S.; Suzuki, Y.; Gottschalk, L.; Shaw, P.J. Inducing sleep by remote control facilitates memory consolidation in Drosophila. Science 2011, 332, 1571–1576. [Google Scholar] [CrossRef] [Green Version]
- Donlea, J.M.; Pimentel, D.; Miesenböck, G. Neuronal machinery of sleep homeostasis in Drosophila. Neuron 2014, 81, 860–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liu, Q.; Tabuchi, M.; Wu, M.N. Sleep Drive Is Encoded by Neural Plastic Changes in a Dedicated Circuit. Cell 2016, 165, 1347–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donlea, J.M.; Pimentel, D.; Talbot, C.B.; Kempf, A.; Omoto, J.J.; Hartenstein, V.; Miesenböck, G. Recurrent Circuitry for Balancing Sleep Need and Sleep. Neuron 2018, 97, 378–389.e4. [Google Scholar] [CrossRef] [Green Version]
- Crocker, A.; Shahidullah, M.; Levitan, I.B.; Sehgal, A. Identification of a Neural Circuit that Underlies the Effects of Octopamine on Sleep:Wake Behavior. Neuron 2010, 65, 670–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, P.J.; Cirelli, C.; Greenspan, R.J.; Tononi, G. Correlates of sleep and waking in Drosophila melanogaster. Science 2000, 287, 1834–1837. [Google Scholar] [CrossRef] [Green Version]
- Tataroglu, O.; Emery, P. Studying circadian rhythms in Drosophila melanogaster. Methods 2014, 68, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Belgacem, Y.H.; Martin, J.R. Hmgcr in the corpus allatum controls sexual dimorphism of locomotor activity and body size via the insulin pathway in Drosophila. Dev. Cell. 2005, 9, 629–638. [Google Scholar] [CrossRef]
- Berger, C.; Renner, S.; Lüer, K.; Technau, G.M. The commonly used marker ELAV is transiently expressed in neuroblasts and glial cells in the Drosophila embryonic CNS. Dev. Dyn. 2007, 236, 3562–3568. [Google Scholar] [CrossRef]
- Helfrich-Förster, C. Differential control of morning and evening components in the activity rhythm of Drosophila melanogaster—Sex-specific differences suggest a different quality of activity. J. Biol. Rhythm. 2000, 15, 135–154. [Google Scholar] [CrossRef]
- Levine, B.D.; Cagan, R.L. Drosophila Lung Cancer Models Identify Trametinib plus Statin as Candidate Therapeutic. Cell Rep. 2016, 14, 1477–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, P.; Han, Z.; Li, X.; Olson, E.N. The mevalonate pathway controls heart formation in Drosophila by isoprenylation of Gγ1. Science 2006, 313, 1301–1303. [Google Scholar] [CrossRef] [Green Version]
- Dus, M.; Lai, J.S.-Y.; Gunapala, K.M.; Min, S.; Tayler, T.D.; Hergarden, A.C.; Geraud, E.; Joseph, C.M.; Suh, G. Nutrient Sensor in the Brain Directs the Action of the Brain-Gut Axis in Drosophila. Neuron 2015, 87, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Cavanaugh, D.J.; Geratowski, J.D.; Wooltorton, J.; Spaethling, J.M.; Hector, C.E.; Zheng, X.; Johnson, E.C.; Eberwine, J.H.; Sehgal, A. Identification of a circadian output circuit for rest: Activity rhythms in drosophila. Cell 2014, 157, 689–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.M.; Daubnerová, I.; Isaac, R.E.; Zhang, C.; Choi, S.; Chung, J.; Kim, Y.-J. A neuronal pathway that controls sperm ejection and storage in female drosophila. Curr. Biol. 2015, 25, 790–797. [Google Scholar] [CrossRef] [Green Version]
- Shafer, O.T.; Keene, A.C. The Regulation of Drosophila Sleep. Curr. Biol. 2021, 31, R38–R49. [Google Scholar] [CrossRef] [PubMed]
- Hirsh, B.J.; Smilowitz, N.R.; Rosenson, R.S.; Fuster, V.; Sperling, L.S. Utilization of and Adherence to Guideline-Recommended Lipid-Lowering Therapy After Acute Coronary Syndrome: Opportunities for Improvement. J. Am. Coll. Cardiol. 2015, 66, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Saver, J.L.; Wu, Y.L.; Tang, S.; Lee, J.; Rao, N.M.; Wang, H.; Jeng, J.; Lee, T.; Chen, P.; et al. Utilization of statins beyond the initial period after stroke and 1-year risk of recurrent stroke. J. Am. Heart Assoc. 2017, 6, e005658. [Google Scholar] [CrossRef]
- De Vera, M.; Bhole, V.; Burns, L.C.; Lacaille, D. Impact of statin adherence on cardiovascular disease and mortality outcomes: A systematic review. Br. J. Clin. Pharmacol. 2014, 78, 684–698. [Google Scholar] [CrossRef]
- de Velasco, B.; Erclik, T.; Shy, D.; Sclafani, J.; Lipshitz, H.; McInnes, R.; Hartenstein, V. Specification and development of the pars intercerebralis and pars lateralis, neuroendocrine command centers in the Drosophila brain. Dev. Biol. 2007, 302, 309–323. [Google Scholar] [CrossRef]
- Siegmund, T.; Korge, G. Innervation of the ring gland of drosophila melanogaster. J. Comp. Neurol. 2001, 431, 481–491. [Google Scholar] [CrossRef]
- Seugnet, L.; Suzuki, Y.; Thimgan, M.; Donlea, J.; Gimbel, S.I.; Gottschalk, L.; Duntley, S.P.; Shaw, P.J. Identifying sleep regulatory genes using a Drosophila model of insomnia. J. Neurosci. 2009, 29, 7148–7157. [Google Scholar] [CrossRef]
- King, A.N.; Sehgal, A. Molecular and circuit mechanisms mediating circadian clock output in the Drosophila brain. Eur. J. Neurosci. 2020, 51, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seugnet, L.; Dissel, S.; Thimgan, M.; Cao, L.; Shaw, P.J. Identification of genes that maintain behavioral and structural plasticity during sleep loss. Front. Neural Circuits 2017, 11, 79. [Google Scholar] [CrossRef] [Green Version]
- DiAngelo, J.R.; Erion, R.; Crocker, A.; Sehgal, A. The central clock neurons regulate lipid storage in Drosophila. PLoS ONE 2011, 6, e19921. [Google Scholar] [CrossRef] [Green Version]
- Rajan, A.; Perrimon, N. Drosophila cytokine unpaired 2 regulates physiological homeostasis by remotely controlling insulin secretion. Cell 2012, 151, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Rulifson, E.J.; Kim, S.K.; Nusse, R. Ablation of insulin-producing neurons in files: Growth and diabetic phenotypes. Science 2002, 296, 1118–1120. [Google Scholar] [CrossRef]
- Ikeya, T.; Galic, M.; Belawat, P.; Nairz, K.; Hafen, E. Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr. Biol. 2002, 12, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Brown, M.R. Localization of an insulin-like peptide in brains of two flies. Cell Tissue Res. 2001, 304, 317–321. [Google Scholar] [CrossRef]
- Cong, X.; Wang, M.H.; Liu, Z.; He, C.; An, C.; Zhao, Z. Regulation of sleep by insulin-like peptide system in drosophila melanogaster. Sleep 2015, 38, 1075–1083. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.B.; Shah, K.D.; Faville, R.; Kottler, B.; Keene, A.C. Drosophila insulin-like peptide 2 mediates dietary regulation of sleep intensity. PLoS Genet. 2020, 16, e1008270. [Google Scholar] [CrossRef]
- Ohhara, Y.; Kobayashi, S.; Yamakawa-Kobayashi, K.; Yamanaka, N. Adult-specific insulin-producing neurons in Drosophila melanogaster. J. Comp. Neurol. 2018, 526, 1351–1367. [Google Scholar] [CrossRef] [PubMed]
- Donelson, N.C.; Kim, E.Z.; Slawson, J.B.; Vecsey, C.G.; Huber, R.; Griffith, L.C. Correction: High-Resolution Positional Tracking for Long-Term Analysis of Drosophila Sleep and Locomotion Using the “Tracker” Program. PLoS ONE 2012, 7, e37250. [Google Scholar] [CrossRef]
- Hendricks, J.C.; Finn, S.M.; A Panckeri, K.; Chavkin, J.; A Williams, J.; Sehgal, A.; Pack, A. Rest in Drosophila is a sleep-like state. Neuron 2000, 25, 129–138. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsehli, A.M.; Liao, S.; Al-Sabri, M.H.; Vasionis, L.; Purohit, A.; Behare, N.; Clemensson, L.E.; Williams, M.J.; Schiöth, H.B. The Statin Target HMG-Coenzyme a Reductase (Hmgcr) Regulates Sleep Homeostasis in Drosophila. Pharmaceuticals 2022, 15, 79. https://doi.org/10.3390/ph15010079
Alsehli AM, Liao S, Al-Sabri MH, Vasionis L, Purohit A, Behare N, Clemensson LE, Williams MJ, Schiöth HB. The Statin Target HMG-Coenzyme a Reductase (Hmgcr) Regulates Sleep Homeostasis in Drosophila. Pharmaceuticals. 2022; 15(1):79. https://doi.org/10.3390/ph15010079
Chicago/Turabian StyleAlsehli, Ahmed M., Sifang Liao, Mohamed H. Al-Sabri, Lukas Vasionis, Archana Purohit, Neha Behare, Laura E. Clemensson, Michael J. Williams, and Helgi B. Schiöth. 2022. "The Statin Target HMG-Coenzyme a Reductase (Hmgcr) Regulates Sleep Homeostasis in Drosophila" Pharmaceuticals 15, no. 1: 79. https://doi.org/10.3390/ph15010079
APA StyleAlsehli, A. M., Liao, S., Al-Sabri, M. H., Vasionis, L., Purohit, A., Behare, N., Clemensson, L. E., Williams, M. J., & Schiöth, H. B. (2022). The Statin Target HMG-Coenzyme a Reductase (Hmgcr) Regulates Sleep Homeostasis in Drosophila. Pharmaceuticals, 15(1), 79. https://doi.org/10.3390/ph15010079