
The Expression Patterns of Human Cancer-Testis Genes Are Induced through Epigenetic Drugs in Colon Cancer Cells

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Results
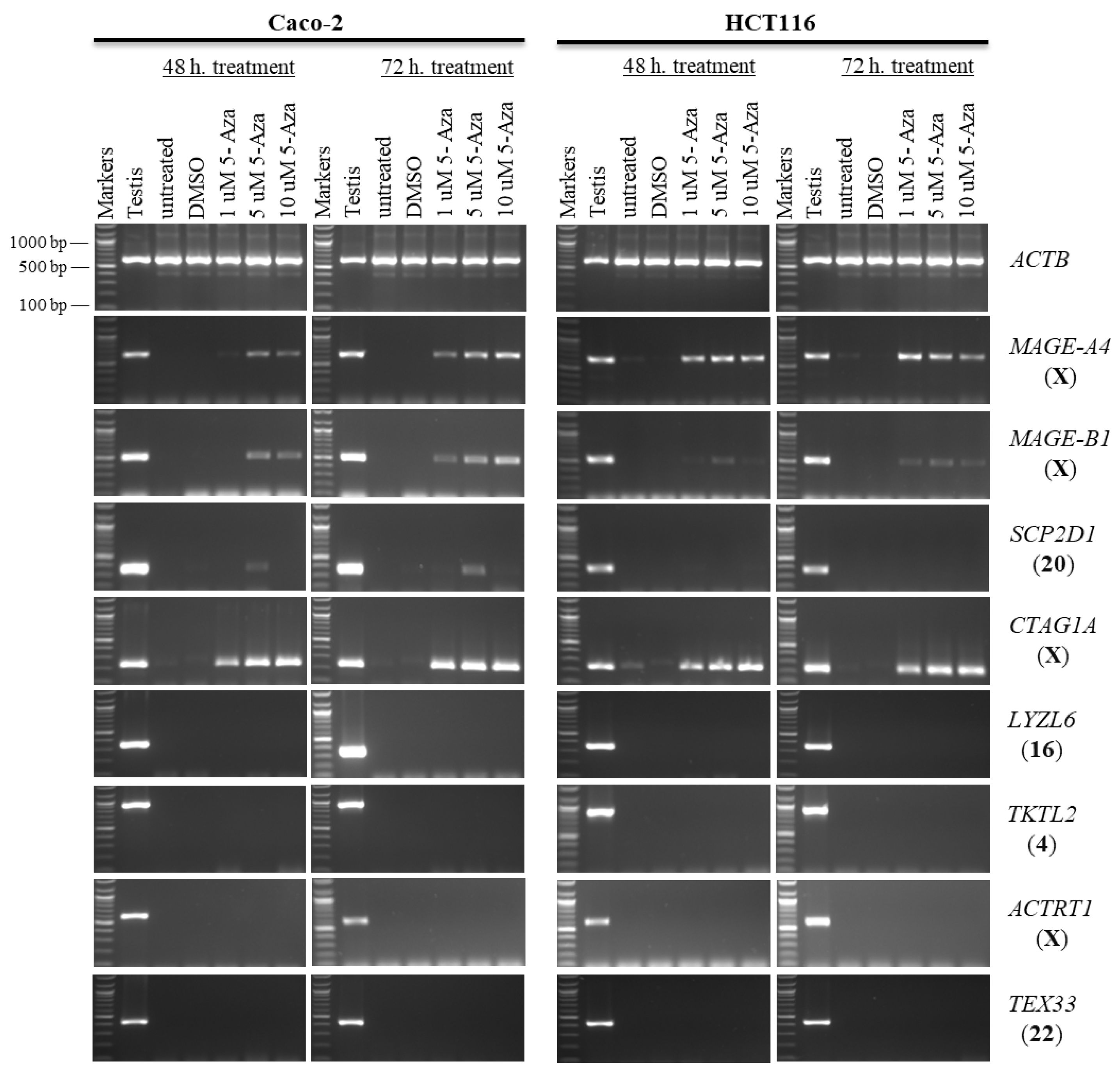
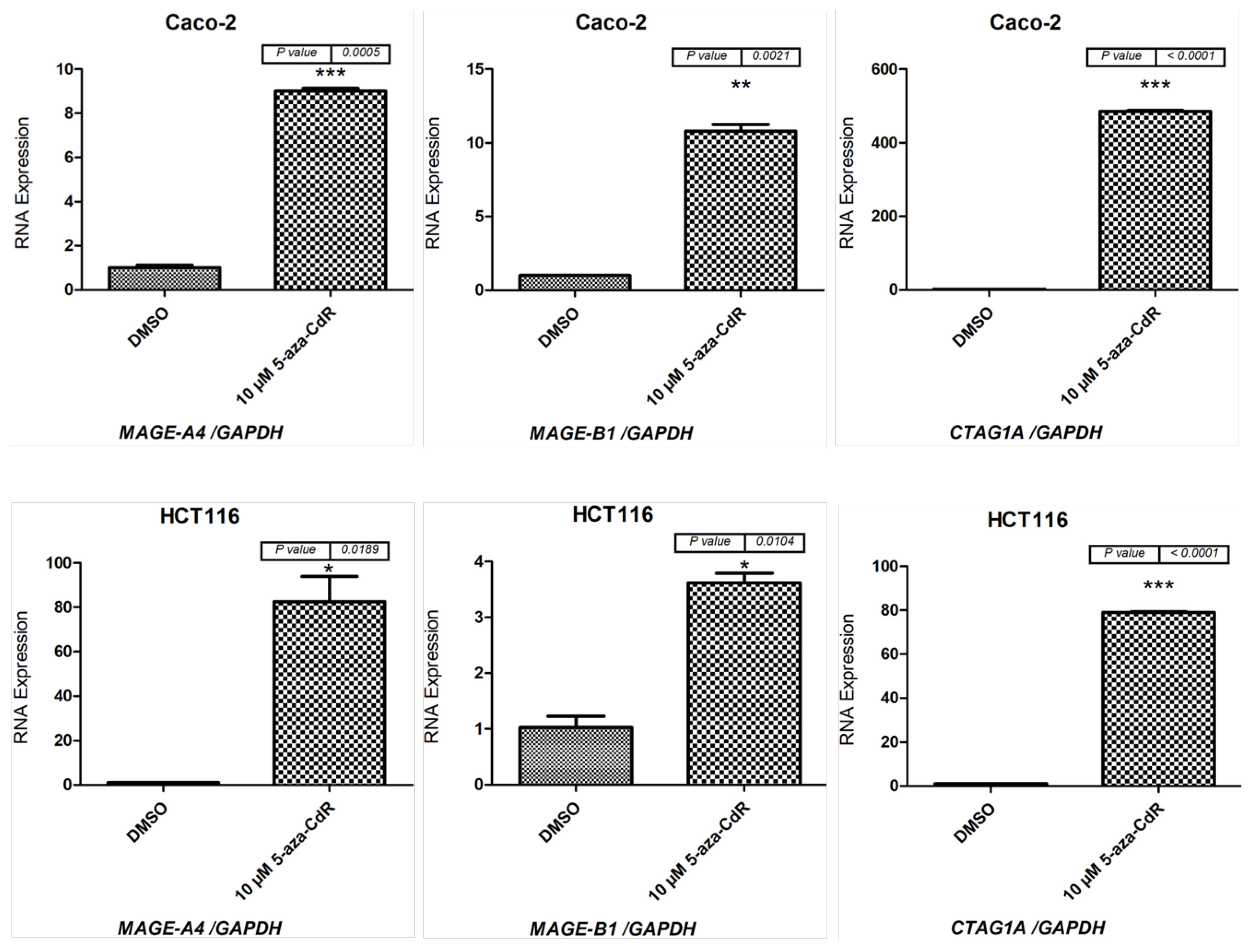
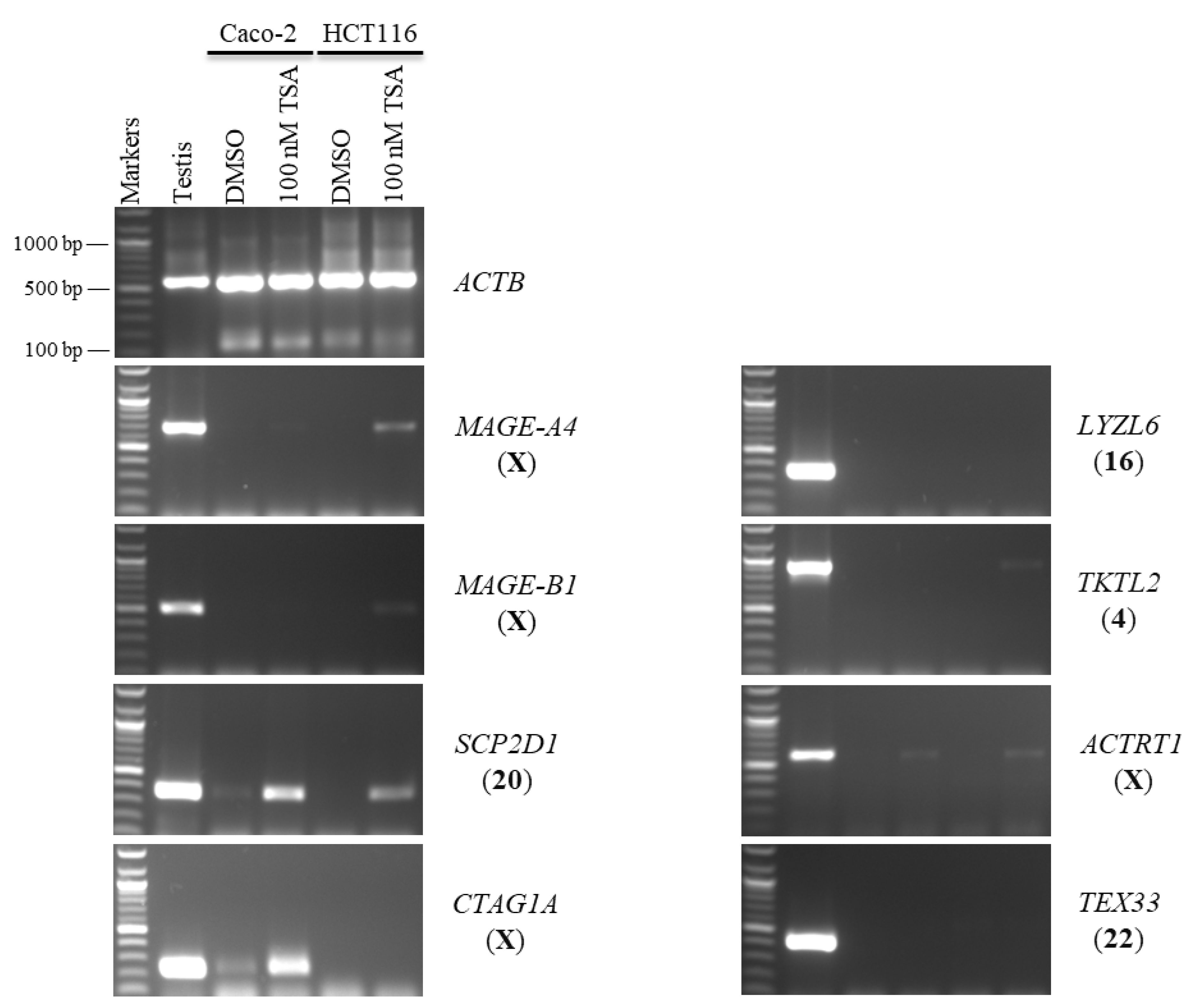
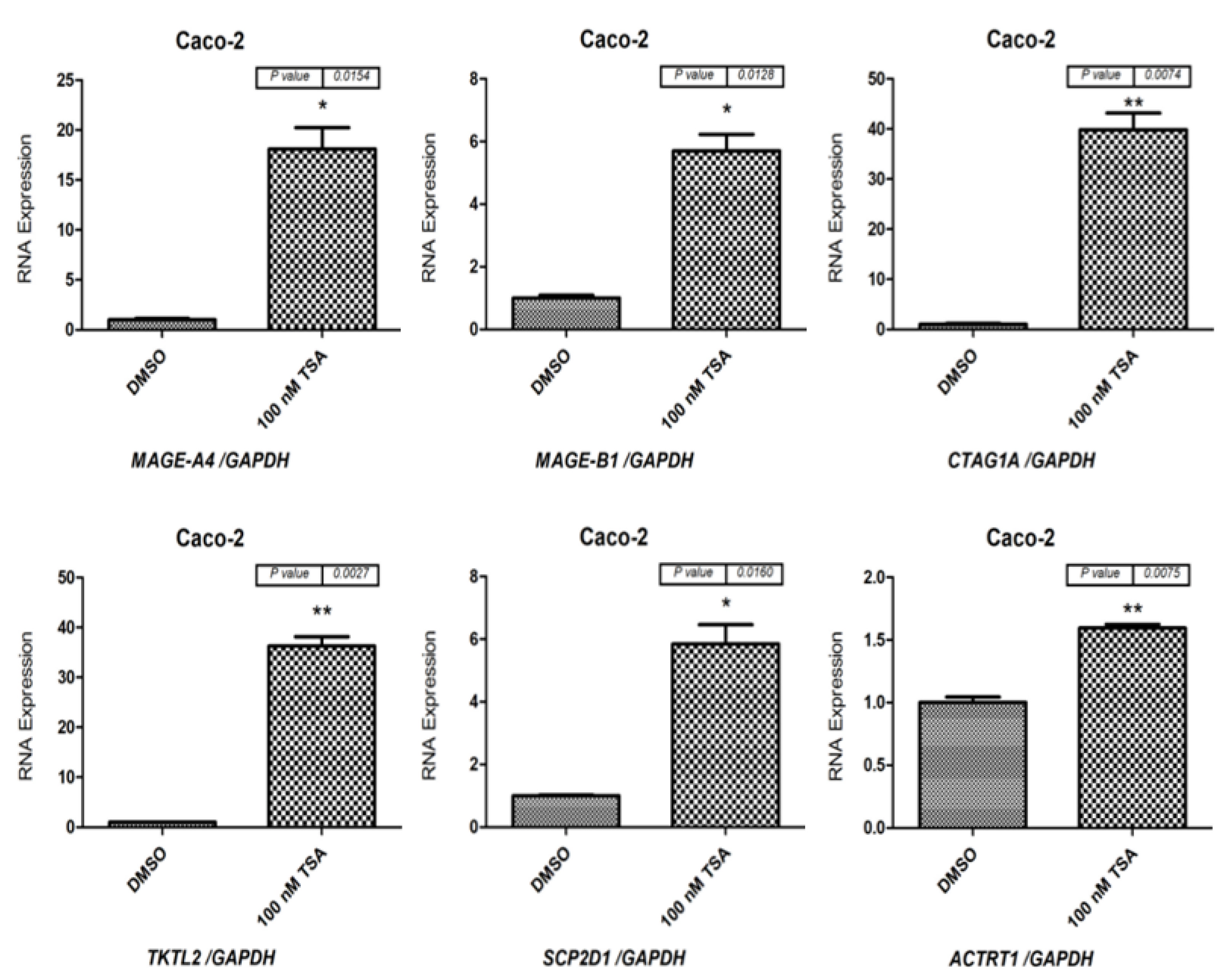
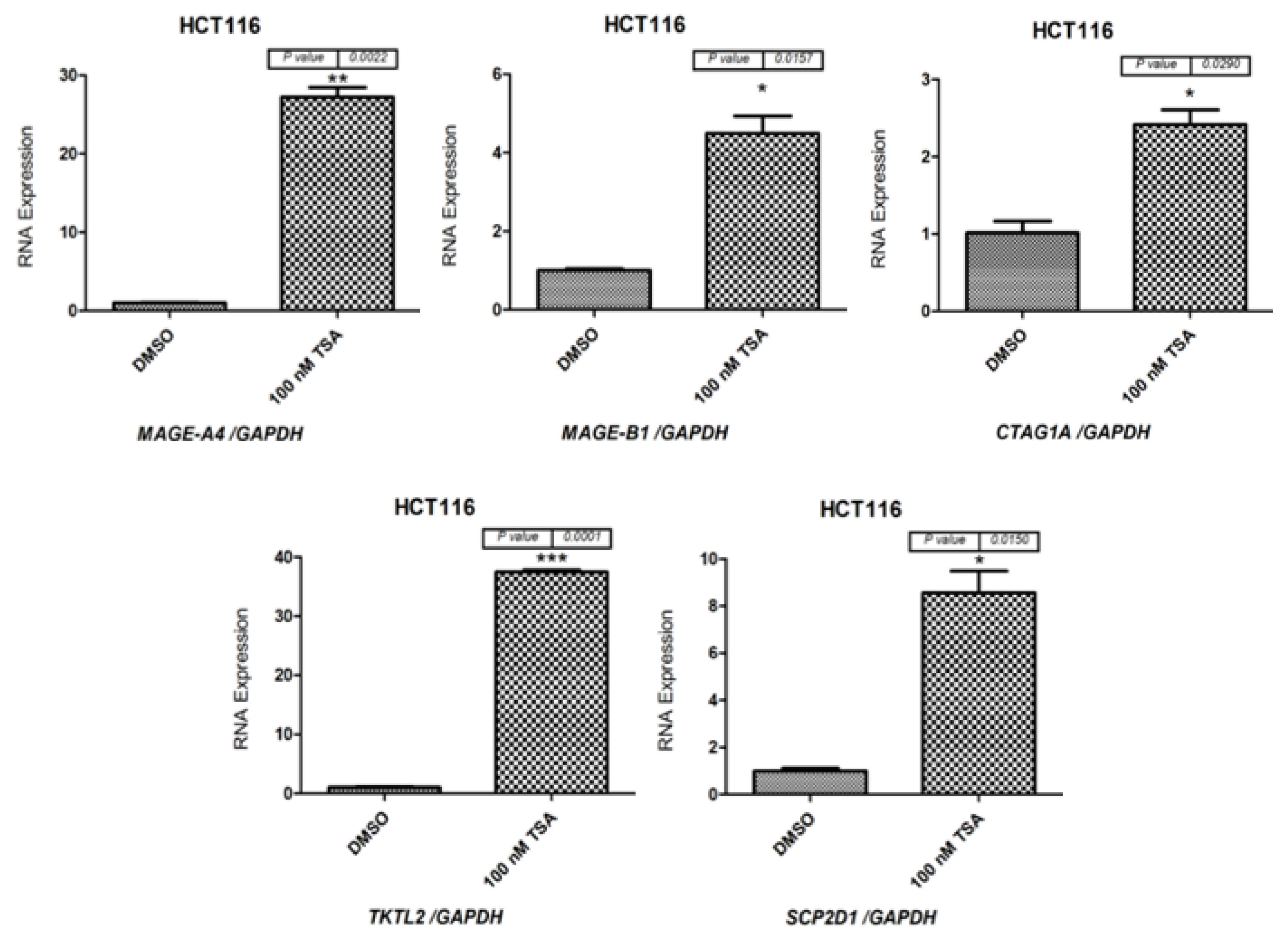
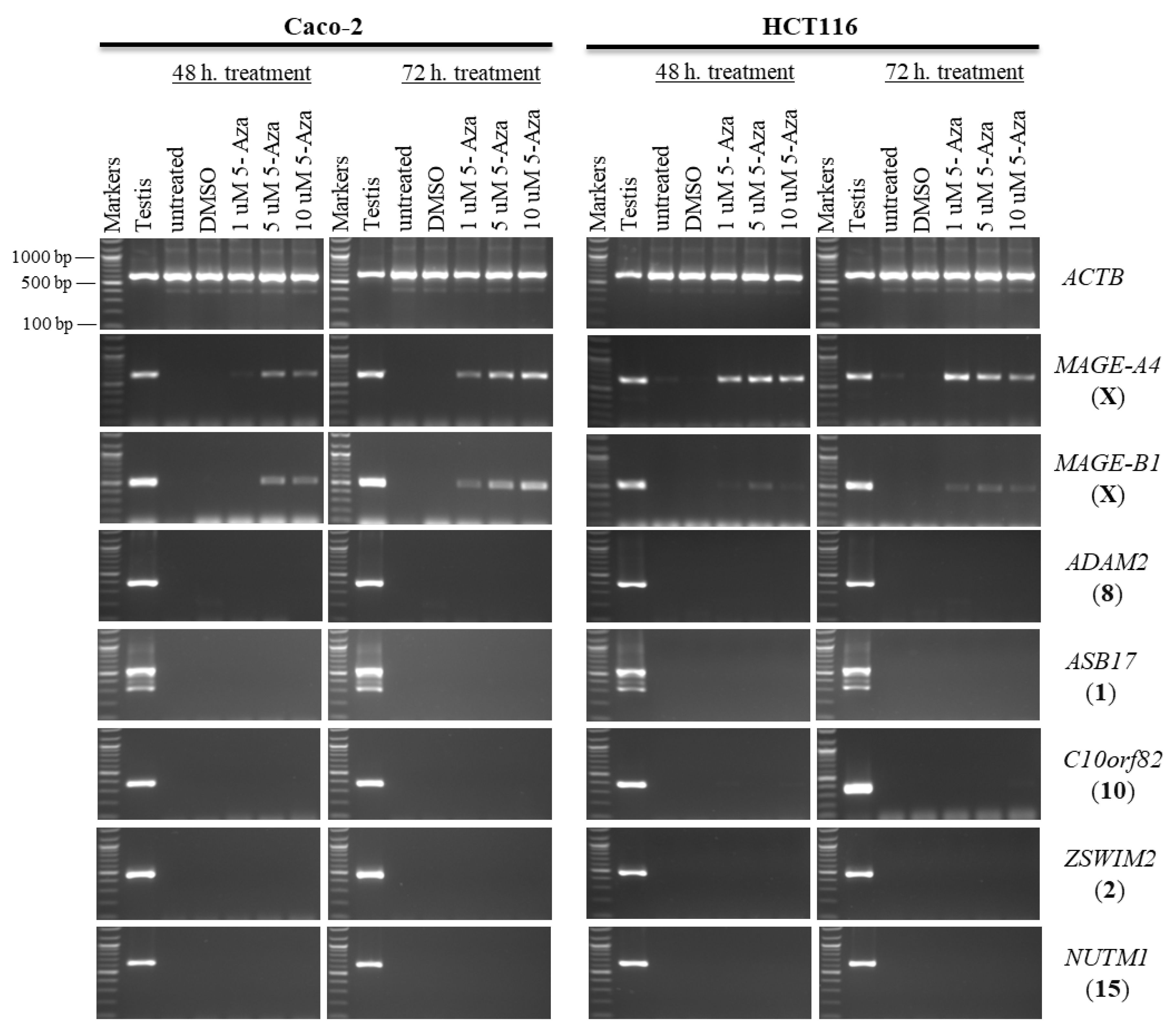
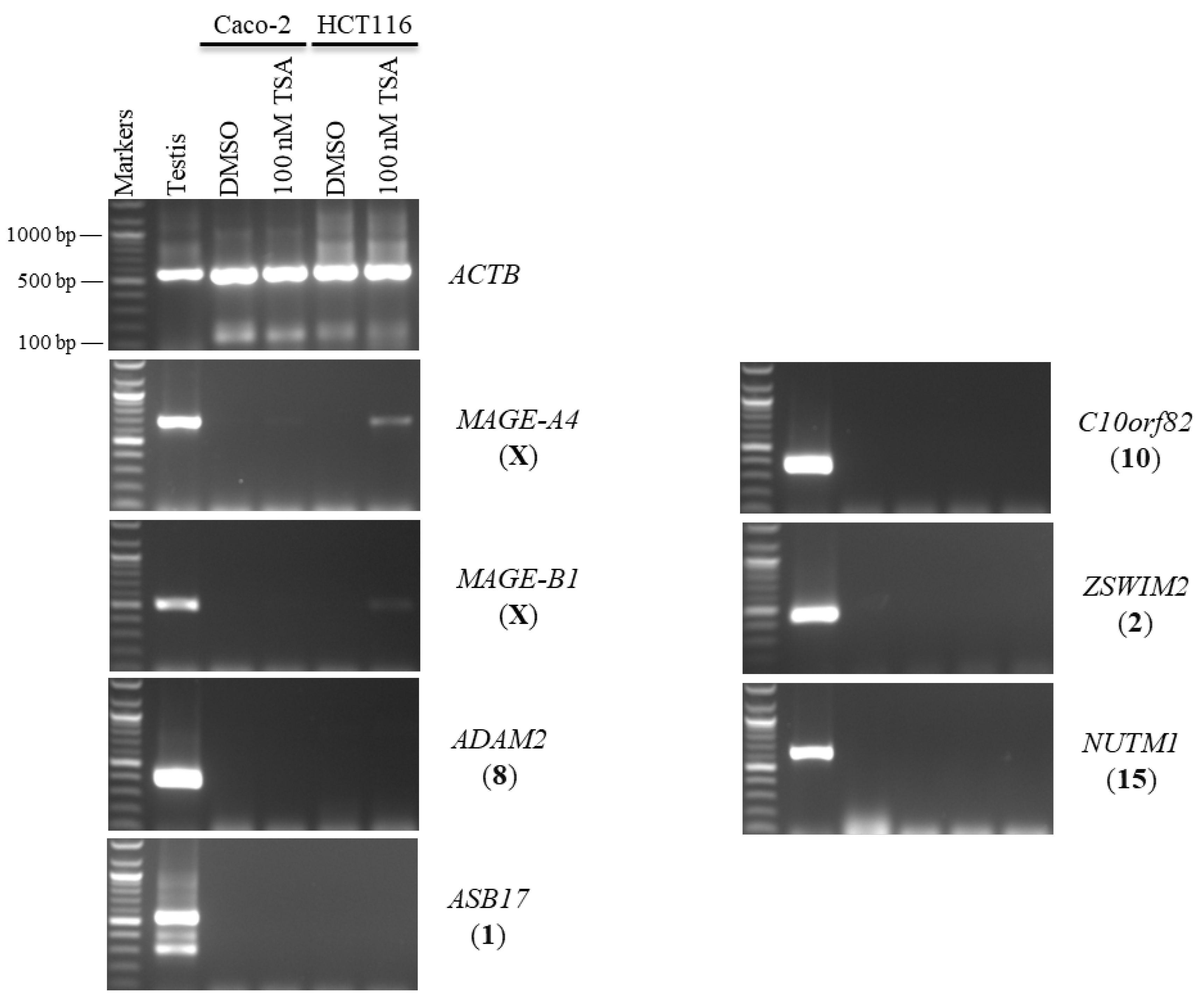
2.1. Effects of 5-aza-CdR and TSA on CT Gene Expression Profiles in CC Cell Lines
2.2. Effects of 5-aza-CdR and TSA on Testis-Specific Gene Expression Profiles in CC Cell Lines
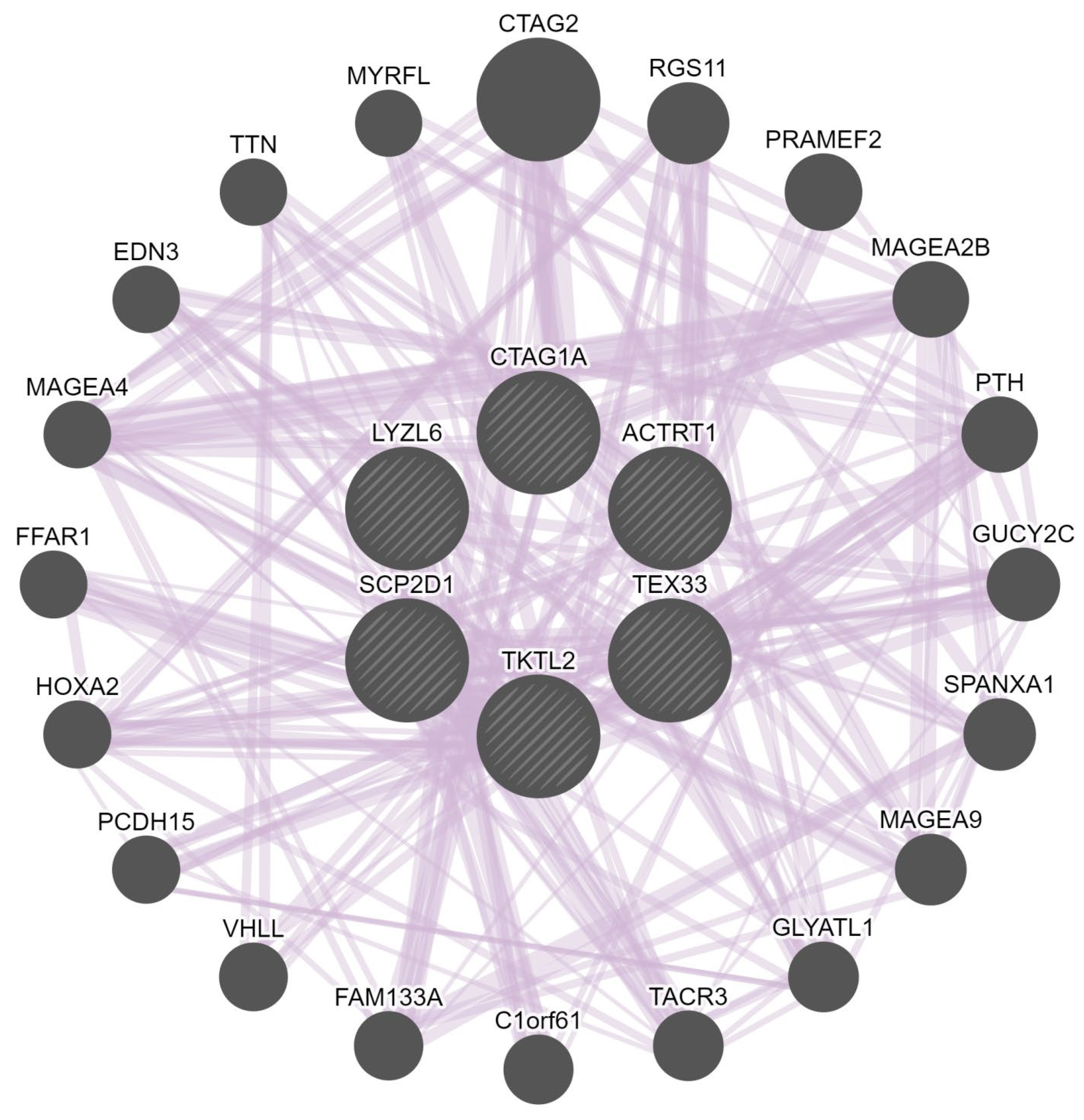
2.3. Gene–Gene Interaction Network
3. Discussion
4. Materials and Methods
4.1. Human CC Cell Line Sources and Cultures
4.2. Treating Cell Lines with 5-aza-2′ Deoxycytidine (5-aza-CdR) or Trichostatin A (TSA)
4.3. Total RNA Isolation and Complementary DNA (cDNA) Synthesis from Cell Cultures
4.4. Primer Design and Setup for Reverse Transcriptase PCR (RT-PCR)
4.5. Agarose Gel Electrophoresis
4.6. Primer Design and Setup for Real Time Quantitative PCR (qRT-PCR)
4.7. Statistical Analysis
4.8. In Silico Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CT | Cancer-testis |
| DNMTi | DNA methyl-transferase inhibitor |
| HDACi | Histone deacetylase inhibitor |
| CC | Colon cancer |
| HDAC | Histone deacetylases |
| 5-aza-CdR | 5-aza-2’-deoxycytidine |
| TSA | Trichostatin A |
| cDNA | Complementary DNA |
| RT-PCR | Reverse transcriptase PCR |
| qRT-PCR | Real time quantitative PCR |
References
- Rasool, M.; Pushparaj, P.N.; Karim, S. Overexpression of CXCL8 gene in Saudi colon cancer patients. Saudi J. Biol. Sci. 2021, 28, 6045–6049. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, M.H.; Alrubie, T.M.; Alamri, A.M.; Almutairi, B.O.; Alrefaei, A.F.; Arafah, M.M.; Alanazi, M.; Semlali, A. Cancer-Testis Gene Biomarkers Discovered in Colon Cancer Patients. Genes 2022, 13, 807. [Google Scholar] [CrossRef] [PubMed]
- Almatroudi, A. The Incidence Rate of Colorectal Cancer in Saudi Arabia: An Observational Descriptive Epidemiological Analysis. Int. J. Gen. Med. 2020, 13, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Almatrafi, A.; Feichtinger, J.; Vernon, E.G.; Escobar, N.G.; Wakeman, J.A.; Larcombe, L.D.; McFarlane, R.J. Identification of a class of human cancer germline genes with transcriptional silencing refractory to the hypomethylating drug 5-aza-2′-deoxycytidine. Oncoscience 2014, 1, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsen, M.K.; Traynor, S.; Stæhr, M.; Duijf, P.G.; Nielsen, A.Y.; Terp, M.G.; Pedersen, C.B.; Guldberg, P.; Ditzel, H.J.; Gjerstorff, M.F. The Cancer/Testis Antigen Gene VCX2 Is Rarely Expressed in Malignancies but Can Be Epigenetically Activated Using DNA Methyltransferase and Histone Deacetylase Inhibitors. Front. Oncol. 2020, 10, 584024. [Google Scholar] [CrossRef] [PubMed]
- Colemon, A.; Harris, T.M.; Ramanathan, S. DNA hypomethylation drives changes in MAGE-A gene expression resulting in alteration of proliferative status of cells. Genes Environ. 2020, 42, 24. [Google Scholar] [CrossRef] [PubMed]
- Chüeh, A.C.; Liew, M.-S.; Russell, P.A.; Walkiewicz, M.; Jayachandran, A.; Starmans, M.H.; Boutros, P.C.; Wright, G.; Barnett, S.A.; Mariadason, J.M.; et al. Promoter hypomethylation of NY-ESO-1, association with clinicopathological features and PD-L1 expression in non-small cell lung cancer. Oncotarget 2017, 8, 74036–74048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, P.; Paluch, B.E.; Matsuzaki, J.; James, S.R.; Collamat-Lai, G.; Blagitko-Dorfs, N.; Ford, L.A.; Naqash, R.; Lübbert, M.; Karpf, A.R.; et al. Induction of cancer testis antigen expression in circulating acute myeloid leukemia blasts following hypomethylating agent monotherapy. Oncotarget 2016, 7, 12840–12856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caponigro, F.; Basile, M.; De Rosa, V.; Normanno, N. New Drugs in Cancer Therapy, National Tumor Institute, Naples, 17–18 June 2004. Anti-Cancer Drugs 2005, 16, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Feichtinger, J.; Aldeailej, I.; Anderson, R.; Almutairi, M.; Almatrafi, M.; Alsiwiehri, N.; Griffiths, K.; Stuart, N.; Wakeman, J.A.; Larcombe, L.; et al. Meta-analysis of clinical data using human meiotic genes identifies a novel cohort of highly restricted cancer-specific marker genes. Oncotarget 2012, 3, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Sammut, S.J.; Feichtinger, J.; Stuart, N.; Wakeman, J.A.; Larcombe, L.; McFarlane, R.J. A novel cohort of cancer-testis biomarker genes revealed through meta-analysis of clinical data sets. Oncoscience 2014, 1, 349–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feichtinger, J.; McFarlane, R.J.; Larcombe, L.D. CancerEST: A web-based tool for automatic meta-analysis of public EST data. Database 2014, 2014, bau024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, P.; Sharghi, M.; Mansourkhani, S.M.; Ardekan, A.P.; Jouybari, L.; Daraei, N.; Peiro, K.; Mohamadian, S.; Rezaei, M.; Heidari, M.; et al. Medicinal Plants in the Prevention and Treatment of Colon Cancer. Oxidative Med. Cell. Longev. 2019, 2019, 2075614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Luo, J.; Guo, J. Development and validation of a five-immune gene prognostic risk model in colon cancer. BMC Cancer 2020, 20, 395. [Google Scholar] [CrossRef]
- Fratta, E.; Coral, S.; Covre, A.; Parisi, G.; Colizzi, F.; Danielli, R.; Nicolay, H.J.M.; Sigalotti, L.; Maio, M. The biology of cancer testis antigens: Putative function, regulation and therapeutic potential. Mol. Oncol. 2011, 5, 164–182. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-F.; Ren, P.; Shen, W.-Z.; Jin, X.; Zhang, J. The expression, modulation and use of cancer-testis antigens as potential biomarkers for cancer immunotherapy. Am. J. Transl. Res. 2020, 12, 7002–7019. [Google Scholar] [PubMed]
- Cannuyer, J.; Loriot, A.; Parvizi, G.K.; De Smet, C. Epigenetic Hierarchy within the MAGEA1 Cancer-Germline Gene: Promoter DNA Methylation Dictates Local Histone Modifications. PLoS ONE 2013, 8, e58743. [Google Scholar] [CrossRef] [PubMed]
- James, S.R.; Cedeno, C.D.; Sharma, A.; Zhang, W.; Mohler, J.L.; Odunsi, K.; Wilson, E.M.; Karpf, A.R. DNA methylation and nucleosome occupancy regulate the cancer germline antigen gene MAGEA11. Epigenetics 2013, 8, 849–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, R.; Kulkarni, P.; Hannenhalli, S. Derepression of Cancer/Testis Antigens in cancer is associated with distinct patterns of DNA Hypomethylation. BMC Cancer 2013, 13, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Official Gene | Forward and Reverse Primer Directions and Sequences (from 5′→3′) | Ta | Product Size (bp) | |
|---|---|---|---|---|
| Symbol | Full name | |||
| ACTB | Actin beta | FP: AGAAAATCTGGCACCACACC | 58 | 553 |
| RP: AGGAAGGAAGGCTGGAAGAG | ||||
| MAGE-A4 | MAGE family member A4 | FP: CTACCATCAGCTTCACTTGC | 647 | |
| RP: CTCCAGGACTTTCACATAGC | ||||
| MAGE-B1 | MAGE family member B1 | FP: CAGGAATGCTGATGCACTTC | 524 | |
| RP: GAGGACTTTCATCTTGGTGG | ||||
| ACTRT1 | Actin related protein T1 | FP: GGGATGACATGGAGAAACTC | 591 | |
| RP: CCATTTTTGAGAGTCCTGGG | ||||
| ASB17 | Ankyrin repeat and SOCS box containing 17 | FP: GTGGGGATATCACTGTTACG | 542 | |
| RP: GCACTCTGGAACATAGTACC | ||||
| LYZL6 | Lysozyme like 6 | FP: GGCGCTACTCATCTATTTGG | 348 | |
| RP: CCGGACACAATCCTTTTTGC | ||||
| TKTL2 | Transketolase like 2 | FP: AGGTACTGCATGTGGAATGG | 896 | |
| RP: CATCTTCTCCAGTGGATACC | ||||
| ZSWIM2 | Zinc finger SWIM-type containing 2 | FP: GACAAACACCTTGGGATTCC | 469 | |
| RP: GGCATGAATTGCACTTGTGG | ||||
| ADAM2 | ADAM metallopeptidase domain 2 | FP: GTCTTGTTTCTGCTCAGCGG | 60 | 397 |
| RP: AGCCAACTGAAGACTCCAGG | ||||
| C10orf82 | Chromosome 10 open reading frame 82 | FP: CTGCCAAGGAATGTCCAAG | 370 | |
| RP: ATGTGCCTTCTTGGCCCTCT | ||||
| TEX33 | Testis expressed 33 | FP: GATCCTCCTCGAGAGAGAAC | 426 | |
| RP: GCCAGTGTTCTAAGTCCCTC | ||||
| SCP2D1 | SCP2 sterol binding domain containing 1 | FP: CAGTTCGAGGTTCTGGGTTC | 369 | |
| RP: GCTAAGCAGAACCTTGCCAC | ||||
| NUTM1 | NUT midline carcinoma family member 1 | FP: CACCACCAGTTGCTCAACTG | 623 | |
| RP: CTCCTTCACAGCTTCTGGTG | ||||
| CTAG1A | Cancer/testis antigen 1A | FP: CCTGCTTGAGTTCTACCTCG | 235 | |
| RP: CTGCGTGATCCACATCAACA | ||||
| Official Gene Symbol | Forward and Reverse Primer Directions and Sequences (from 5′→3′) | Product Size (bp) |
|---|---|---|
| GAPDH | FP: GGGAAGCTTGTCATCAATGG | 173 |
| RP: GAGATGATGACCCTTTTGGC | ||
| MAGE-A4 | FP: CTACCATCAGCTTCACTTGC | 134 |
| RP: AGCCAACTCATCCACCTTGT | ||
| MAGE-B1 | FP: GAAGGCAGATATGCTGAAGG | 125 |
| RP: CACTAGGGTTGTCTTCCTTC | ||
| TKTL2 | FP: CATGGTAAGTGTGGCACTAG | 149 |
| RP: CACAGTGGGAACCAATAAGG | ||
| SCP2D1 | FP: CCAGCAGACACTGTCTTTAC | 129 |
| RP: CTTCCAGCTAAGCAGAACCT | ||
| ACTRT1 | FP: GGGATGACATGGAGAAACTC | 153 |
| RP: GCACACTGAAGGTCTCAAAC | ||
| CTAG1A | FP: CCTGCTTGAGTTCTACCTCG | 132 |
| RP: CCGGACACAGTGAACTCCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almutairi, M.H.; Alrubie, T.M.; Almutairi, B.O.; Alamri, A.M.; Alrefaei, A.F.; Arafah, M.M.; Alanazi, M.; Semlali, A. The Expression Patterns of Human Cancer-Testis Genes Are Induced through Epigenetic Drugs in Colon Cancer Cells. Pharmaceuticals 2022, 15, 1319. https://doi.org/10.3390/ph15111319
Almutairi MH, Alrubie TM, Almutairi BO, Alamri AM, Alrefaei AF, Arafah MM, Alanazi M, Semlali A. The Expression Patterns of Human Cancer-Testis Genes Are Induced through Epigenetic Drugs in Colon Cancer Cells. Pharmaceuticals. 2022; 15(11):1319. https://doi.org/10.3390/ph15111319
Chicago/Turabian StyleAlmutairi, Mikhlid H., Turki M. Alrubie, Bader O. Almutairi, Abdullah M. Alamri, Abdulwahed F. Alrefaei, Maha M. Arafah, Mohammad Alanazi, and Abdelhabib Semlali. 2022. "The Expression Patterns of Human Cancer-Testis Genes Are Induced through Epigenetic Drugs in Colon Cancer Cells" Pharmaceuticals 15, no. 11: 1319. https://doi.org/10.3390/ph15111319