
Chemical Composition of Impatiens textori Miq. Flower Absolute and Its Potential Wound Repair and Anti-Melanogenesis-Promoting Activities in Skin Cells

 and
and
Abstract
:1. Introduction
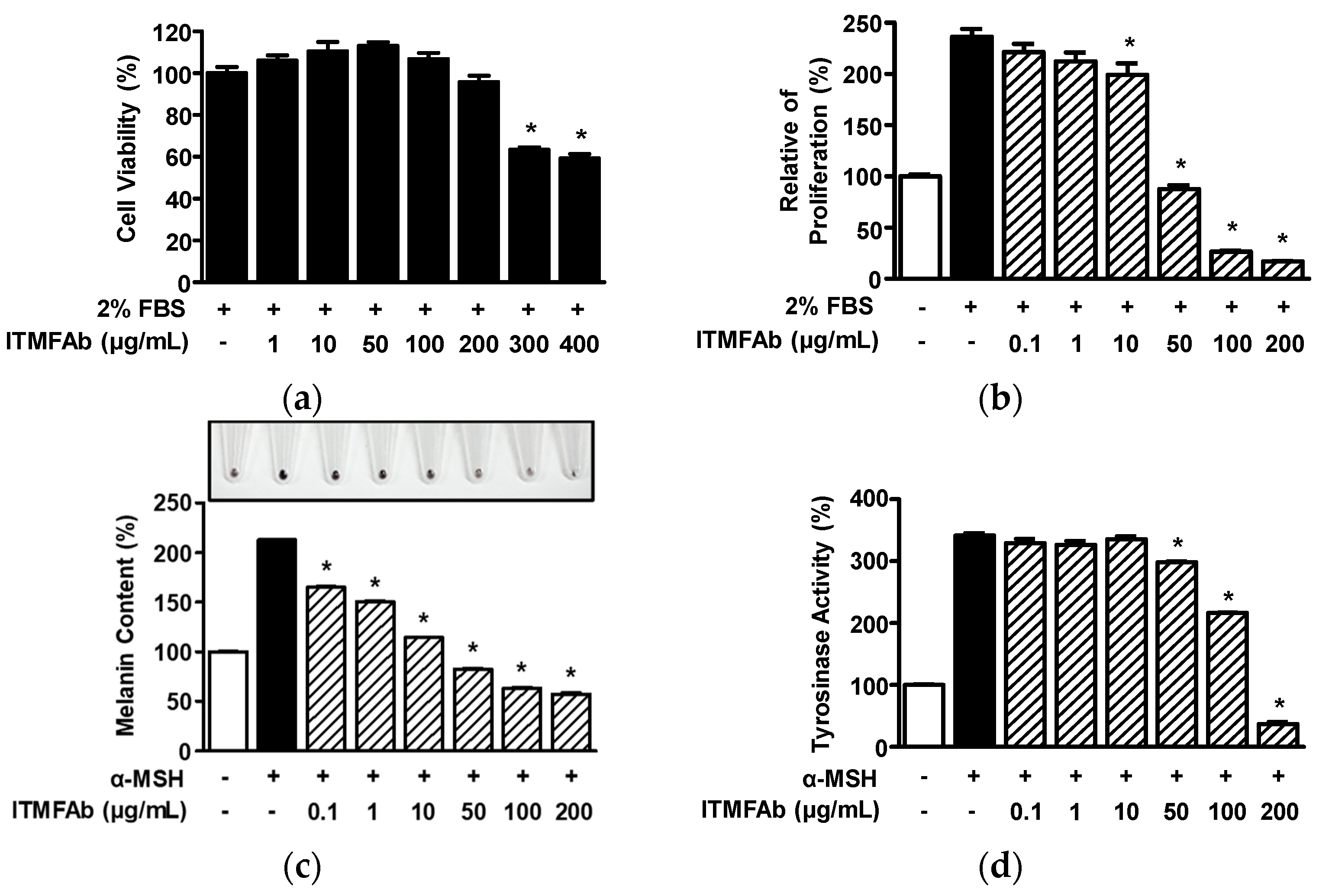
2. Results
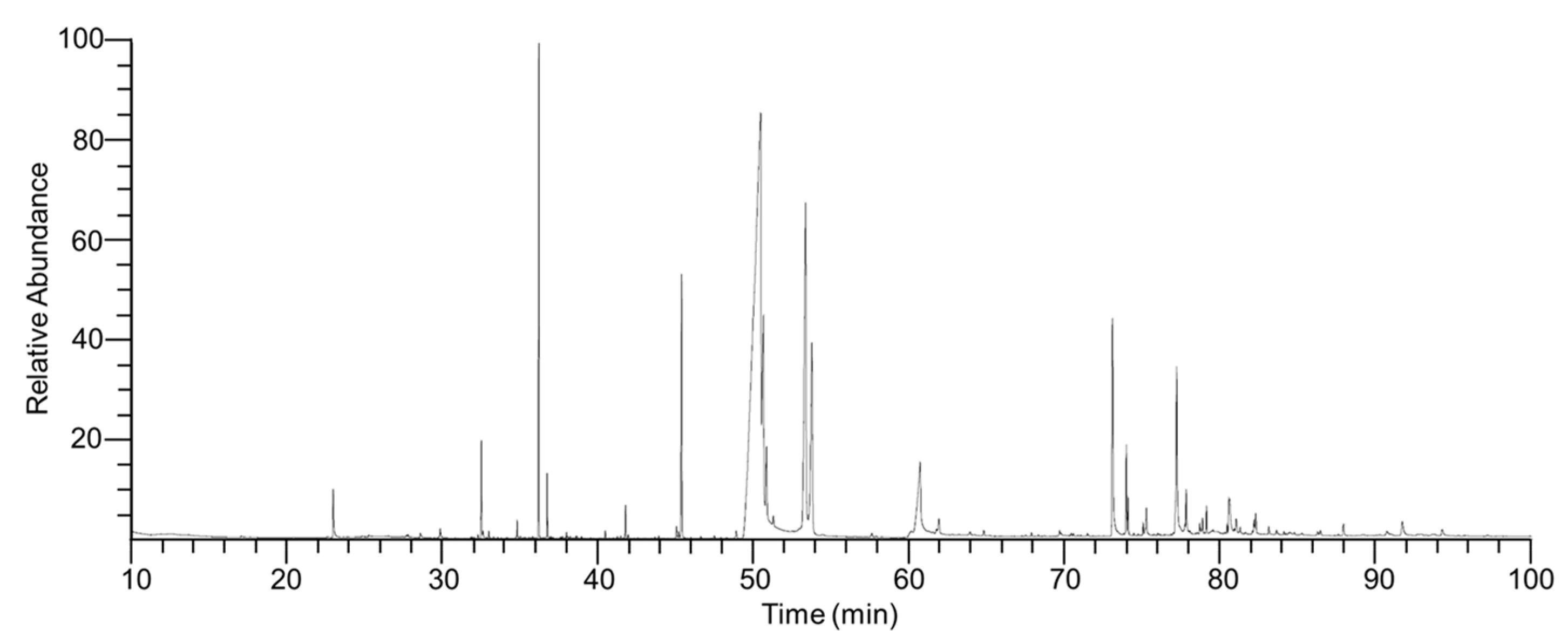
2.1. Chemical Composition of ITMFAb
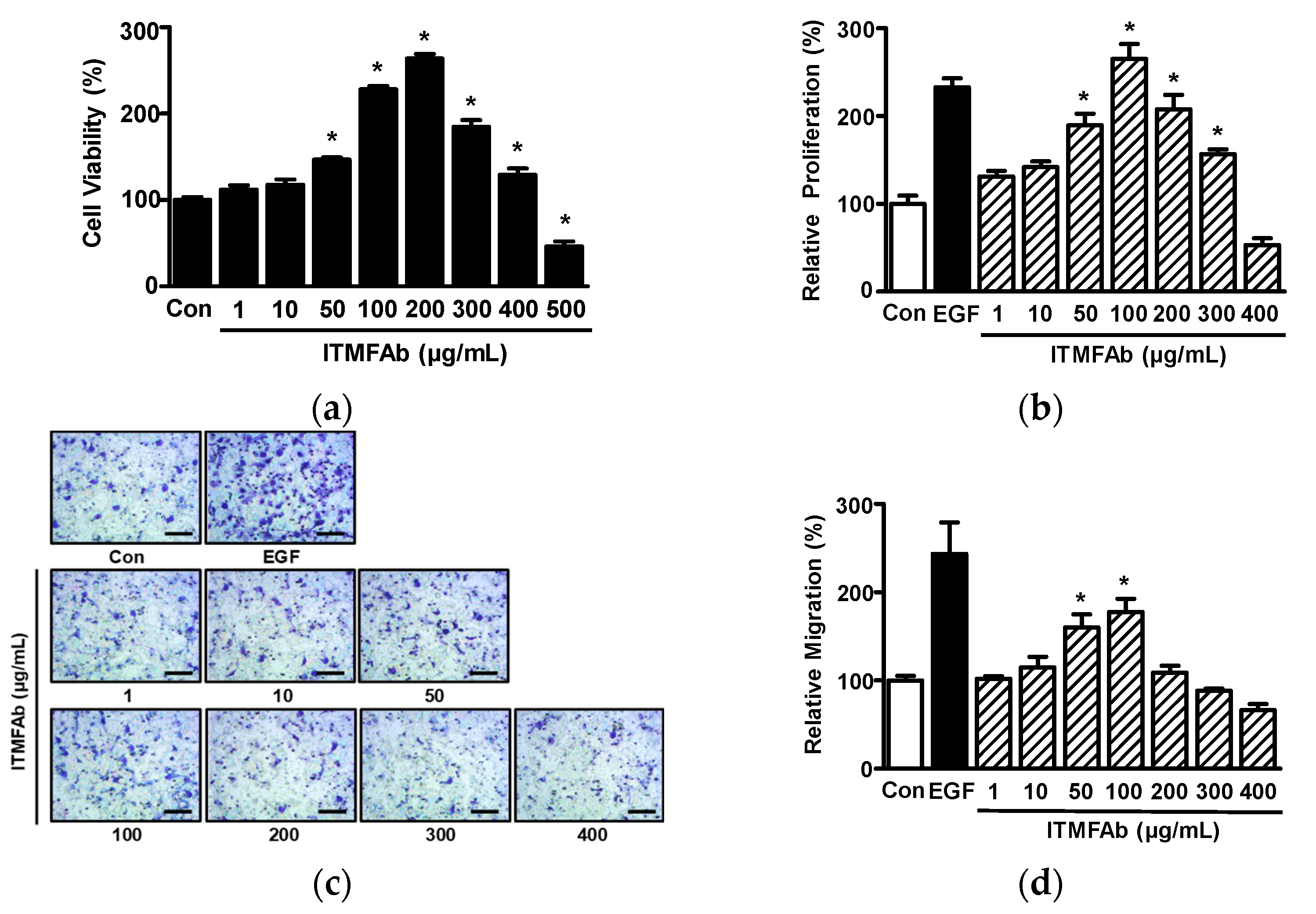
2.2. Effects of ITMFAb on Keratinocyte Proliferation and Migration
2.3. Effect of ITMFAb on Keratinocyte Sprout Outgrowth
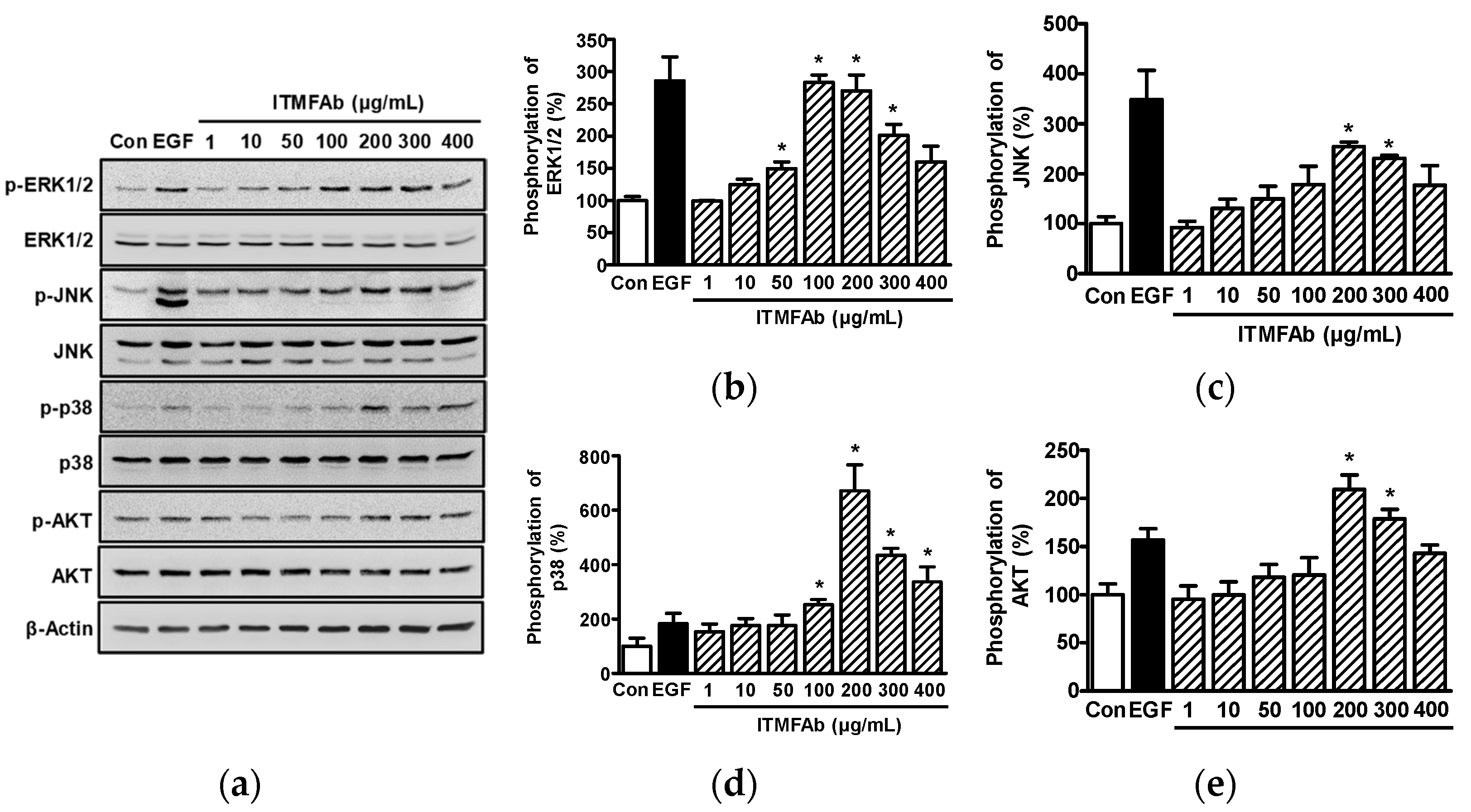
2.4. ITMFAb-Induced Changes in the Activations of Kinases in HaCaT Cells
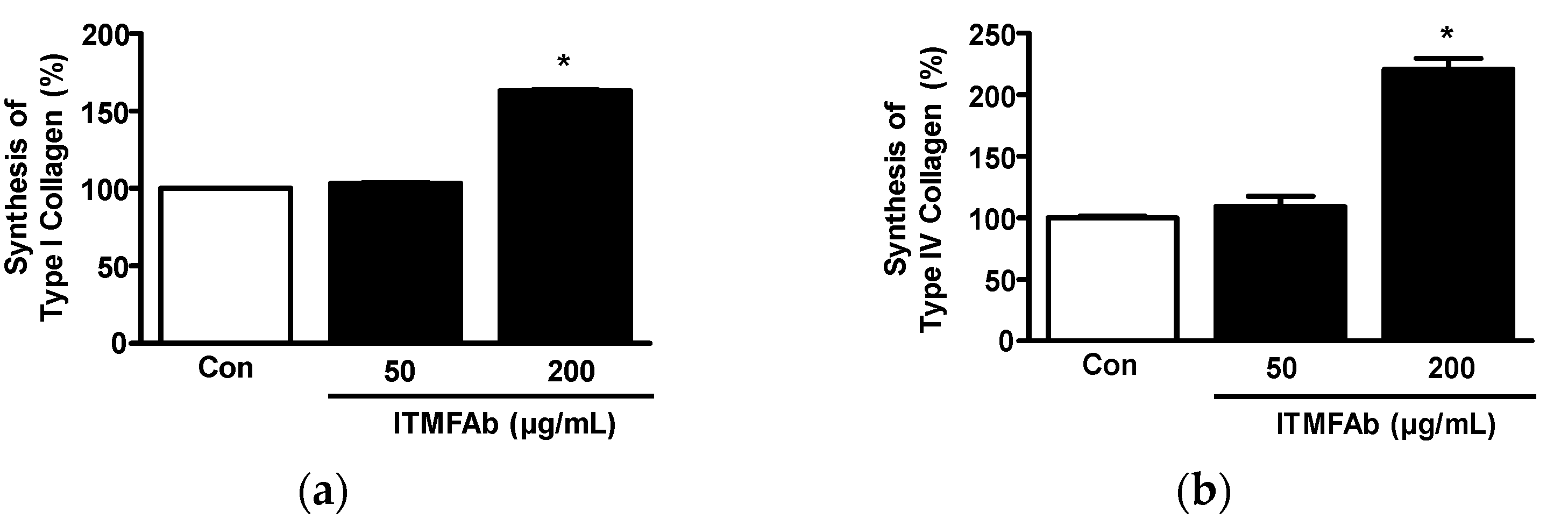
2.5. ITMFAb-Induced Changes in Collagen Synthesis in HaCaT Cells
2.6. Effects of ITMFAb on B16BL6 Melanoma Cells
2.7. Effects of ITMFAb on Melanogenesis-Regulatory Molecules in B16BL6 Melanoma Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Impatiens textori Miq. Flower Absolute
4.3. Analysis and Identification of Compounds in ITMFAb
4.4. Cell Culture
4.5. Cell Viability Assays
4.6. Proliferation Assays
4.7. Migration Assay
4.8. Collagen Sprout Assay
4.9. Collagen Synthesis Assay
4.10. Western Blotting
4.11. Melanin Content Assay
4.12. Tyrosinase Activity Assays
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of acute wound healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2018, 146, 344–365. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.M.; Gaudino, G. Cellular and molecular facets of keratinocyte reepithelization during wound healing. Exp. Cell Res. 2005, 304, 274–286. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin Against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Huang, J.; Lu, J.; Ding, Y.; Jiang, L.; Hu, S.; Chen, J.; Zeng, Q. The role and mechanism of Asian medicinal plants in treating skin pigmentary disorders. J. Ethnopharmacol. 2019, 245, 112173. [Google Scholar] [CrossRef]
- Qian, W.; Liu, W.; Zhu, D.; Cao, Y.; Tang, A.; Gong, G.; Su, H. Natural skin-whitening compounds for the treatment of melanogenesis (Review). Exp. Ther. Med. 2020, 20, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Huh, Y.; Lim, K.-M. Anti-Pigmentary Natural Compounds and Their Mode of Action. Int. J. Mol. Sci. 2021, 22, 6206. [Google Scholar] [CrossRef]
- Tatsuzawa, F.; Saito, N.; Mikanagi, Y.; Shinoda, K.; Toki, K.; Shigihara, A.; Honda, T. An unusual acylated malvidin 3-glucoside from flowers of Impatiens textori Miq. (Balsaminaceae). Phytochemistry 2009, 70, 672–674. [Google Scholar] [CrossRef]
- Iwaoka, E.; Oku, H.; Iinuma, M.; Ishiguro, K. Allergy-Preventive Effects of the Flowers of Impatiens textori. Biol. Pharm. Bull. 2010, 33, 714–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang Su New Medical College. Dictionary of Chinese Crude Drugs; Shanghai Scientific Technological Publishers: Shanghai, China, 1977. [Google Scholar]
- Ueda, Y.; Oku, H.; Iinuma, M.; Ishiguro, K. Antianaphylactic and Antipruritic Effects of the Flowers of Impatiens textori MIQ. Biol. Pharm. Bull. 2005, 28, 1786–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Shim, D.-W.; Han, J.-W.; Shin, W.-Y.; Sim, E.-J.; Kim, M.-K.; Heo, K.-H.; Kim, Y.-K.; Koppula, S.; Kim, T.-J.; et al. Anti-inflammatory effect of Impatiens textori Miq. extract via inhibition of NLRP3 inflammasome activation in in vitro and in vivo experimental models. J. Ethnopharmacol. 2015, 170, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Yang, J. Antioxidant, antiproliferative, and α-glucosidase inhibitory activities of extracts from Impatiens textori Miq. J. Med. Plants Res. 2012, 6. [Google Scholar] [CrossRef]
- Park, S.M.; Won, K.J.; Hwang, D.I.; Kim, D.Y.; Bin Kim, H.; Li, Y.; Lee, H.M. Potential Beneficial Effects of Digitaria ciliaris Flower Absolute on the Wound Healing-Linked Activities of Fibroblasts and Keratinocytes. Planta Med. 2020, 86, 348–355. [Google Scholar] [CrossRef]
- Kang, H.M.; Won, K.J.; Kim, D.Y.; Lee, S.Y.; Kim, M.J.; Won, Y.R.; Kim, B.; Lee, H.M. Chemical Composition of Miscanthus sinensis var. purpurascens Flower Absolute and Its Beneficial Effects on Skin Wound Healing and Melanogenesis-Related Cell Activities. Chem. Biodivers. 2021, 18. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-C.; Yang, J.-S.; Chiu, Y.-J.; Tsai, F.-J.; Hsu, Y.-M.; Yin, M.-C.; Juan, Y.-N.; Ho, T.-J.; Chen, H.-P. Dracorhodin perchlorate enhances wound healing via β-catenin, ERK/p38, and AKT signaling in human HaCaT keratinocytes. Exp. Ther. Med. 2021, 22, 1–9. [Google Scholar] [CrossRef]
- Mathew-Steiner, S.; Roy, S.; Sen, C. Collagen in Wound Healing. Bioengineering 2021, 8, 63. [Google Scholar] [CrossRef]
- Abreu-Velez, A.M.; Howard, M.S. Collagen IV in normal skin and in pathological processes. N. Am. J. Med. Sci. 2012, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pazyar, N.; Yaghoobi, R.; Rafiee, E.; Mehrabian, A.; Feily, A. Skin Wound Healing and Phytomedicine: A Review. Ski. Pharmacol. Physiol. 2014, 27, 303–310. [Google Scholar] [CrossRef]
- Galehdari, H.; Negahdari, S.; Kesmati, M.; Rezaie, A.; Shariati, G. Wound healing activity of extracts and formulations of aloe vera, henna, adiantum capillus-veneris, and myrrh on mouse dermal fibroblast cells. Int. J. Prev. Med. 2017, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, A.; Spano, F.; Derler, S.; Adlhart, C.; Spencer, N.; Rossi, R. The relationship between skin function, barrier properties, and body-dependent factors. Skin. Res. Technol. 2018, 24, 165–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haensel, D.; Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: Where we are and where we are heading. Dev. Dyn. 2017, 247, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-Y.; Won, K.-J.; Kim, D.-Y.; Kim, M.-J.; Won, Y.-R.; Kim, N.-Y.; Lee, H.-M. Wound Healing-Promoting and Melanogenesis-Inhibiting Activities of Angelica polymorpha Maxim. Flower Absolute In Vitro and Its Chemical Composition. Molecules 2021, 26, 6172. [Google Scholar] [CrossRef] [PubMed]
- Villegas, L.F.; Marçalo, A.; Martin, J.; Fernández, I.D.; Maldonado, H.; Vaisberg, A.J.; Hammond, G.B. (+)-epi-alpha-bisabolol [correction of bisbolol] is the wound-healing principle of Peperomia galioides: Investigation of the in vivo wound-healing ac-tivity of related terpenoids. J. Nat. Prod. 2001, 64, 1357–1359. [Google Scholar] [CrossRef]
- Beserra, F.P.; Vieira, A.J.; Gushiken, L.F.S.; de Souza, E.O.; Hussni, M.F.; Hussni, C.A.; Nóbrega, R.H.; Martinez, E.R.M.; Jackson, C.J.; de Azevedo Maia, G.L.; et al. Lupeol, a dietary triterpene, enhances wound healing in streptozotocin-induced hyperglycemic rats with modulatory effects on inflammation, oxidative stress, and angiogenesis. Oxi. Med. Cell. Longev. 2019, 2019, 20. [Google Scholar] [CrossRef]
- Beserra, F.P.; Gushiken, L.F.S.; Vieira, A.J.; Bérgamo, D.A.; Bérgamo, P.L.; De Souza, M.O.; Hussni, C.A.; Takahira, R.K.; Nóbrega, R.H.; Martinez, E.R.M.; et al. From Inflammation to Cutaneous Repair: Topical Application of Lupeol Improves Skin Wound Healing in Rats by Modulating the Cytokine Levels, NF-κB, Ki-67, Growth Factor Expression, and Distribution of Collagen Fibers. Int. J. Mol. Sci. 2020, 21, 4952. [Google Scholar] [CrossRef]
- Cardoso, C.R.; Favoreto, S., Jr.; Oliveira, L.L.; Vancim, J.O.; Barban, G.B.; Ferraz, D.B.; Silva, J.S. Oleic acid modulation of the immune response in wound healing: A new approach for skin repair. Immunobiology 2011, 216, 409–415. [Google Scholar] [CrossRef]
- Pană, R.D.P.; Watz, C.-G.; Moacă, E.-A.; Vlaia, L.; Marcovici, I.; Macașoi, I.G.; Borcan, F.; Olariu, I.; Coneac, G.; Drăghici, G.-A.; et al. Oleogel Formulations for the Topical Delivery of Betulin and Lupeol in Skin Injuries—Preparation, Physicochemical Characterization, and Pharmaco-Toxicological Evaluation. Molecules 2021, 26, 4174. [Google Scholar] [CrossRef]
- Shin, J.; Yang, S.J.; Lim, Y. Gamma-tocopherol supplementation ameliorated hyper-inflammatory response during the early cutaneous wound healing in alloxan-induced diabetic mice. Exp. Biol. Med. 2016, 242, 505–515. [Google Scholar] [CrossRef]
- Pereira Beserra, F.; Xue, M.; Maia, G.; Leite Rozza, A.; Helena Pellizzon, C.; Jackson, C. Lupeol, a Pentacyclic Triterpene, Promotes Migration, Wound Closure, and Contractile Effect In Vitro: Possible Involvement of PI3K/Akt and p38/ERK/MAPK Pathways. Molecules 2018, 23, 2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Cellular Signalling and Photobiomodulation in Chronic Wound Repair. Int. J. Mol. Sci. 2021, 22, 11223. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Liu, J.-Q.; Zheng, Z.; Zhang, J.; Wang, S.-Y.; Han, S.-C.; Zhou, Q.; Guan, H.; Li, C.; Su, L.-L.; et al. Human amniotic epithelial stem cells promote wound healing by facilitating migration and proliferation of keratinocytes via ERK, JNK and AKT signaling pathways. Cell Tissue Res. 2016, 365, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Kim, Y.K.; Eun, H.C.; Cho, K.H.; Chung, J.H. All-Trans Retinoic Acid Antagonizes UV-Induced VEGF Production and Angiogenesis via the Inhibition of ERK Activation in Human Skin Keratinocytes. J. Investig. Dermatol. 2006, 126, 2697–2706. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, L.; Zhang, Q.; Wang, W.; Zhang, D.; Jia, J.; Lv, Y.; Yuan, H.; Song, H.; Xiang, F.; et al. Microtubule-associated protein 4 phosphorylation regulates epidermal keratinocyte migration and proliferation. Int. J. Biol. Sci. 2019, 15, 1962–1976. [Google Scholar] [CrossRef]
- Su, L.; Fu, L.; Li, X.; Zhang, Y.; Li, Z.; Wu, X.; Li, Y.; Bai, X.; Hu, D. Loss of CAR promotes migration and proliferation of HaCaT cells and accelerates wound healing in rats via Src-p38 MAPK pathway. Sci. Rep. 2016, 6, 19735. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.Y.; Won, K.J.; Bin Kim, H.; Kim, D.Y.; Kim, M.J.; Won, Y.R.; Lee, H.M. Chemical Composition of Salix koreensis Anderss Flower Absolute and Its Skin Wound Healing Activities In Vitro. Plants 2022, 11, 246. [Google Scholar] [CrossRef]
- Kang, J.; Chen, W.; Xia, J.; Li, Y.; Yang, B.; Chen, B.; Sun, W.; Song, X.; Xiang, W.; Wang, X.; et al. Extracellular matrix secreted by senescent fibroblasts induced by UVB promotes cell proliferation in HaCaT cells through PI3K/AKT and ERK signaling pathways. Int. J. Mol. Med. 2008, 21, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Schüppel, M.; Kürschner, U.; Kleuser, U.; Schäfer-Korting, M.; Kleuser, B. Sphingosine 1-Phosphate Restrains Insulin-Mediated Keratinocyte Proliferation via Inhibition of Akt through the S1P2 Receptor Subtype. J. Investig. Dermatol. 2008, 128, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-E.; Cho, H.; Ishihara, A.; Kim, B.; Kim, O. Cell proliferation and migration mechanism of caffeoylserotonin and serotonin via serotonin 2B receptor in human keratinocyte HaCaT cells. BMB Rep. 2018, 51, 188–193. [Google Scholar] [CrossRef]
- Brett, D. A Review of Collagen and Collagen-based Wound Dressings. Wounds A Compend. Clin. Res. Pract. 2008, 20, 347–356. [Google Scholar]
- O’Toole, E. Extracellular matrix and keratinocyte migration. Clin. Exp. Dermatol. 2001, 26, 525–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, S. Catrix: An easy-to-use collagen treatment for wound healing. Br. J. Community Nurs. 2005, 10, S31–S34. [Google Scholar] [CrossRef]
- Woodley, D.T.; Bachmann, P.M.; O’Keefe, E.J. Laminin inhibits human keratinocyte migration. J. Cell. Physiol. 1988, 136, 140–146. [Google Scholar] [CrossRef]
- Seo, W.-Y.; Kim, J.-H.; Baek, D.-S.; Kim, S.-J.; Kang, S.; Yang, W.S.; Song, J.-A.; Lee, M.-S.; Kim, S.; Kim, Y.-S. Production of recombinant human procollagen type I C-terminal propeptide and establishment of a sandwich ELISA for quantification. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Sierra, J.O.; Kelly, R.; Kirsner, R.S.; Li, J. Wool-derived keratin stimulates human keratinocyte migration and types IV and VII collagen expression. Exp. Dermatol. 2012, 21, 458–460. [Google Scholar] [CrossRef]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, Y.; Wu, Y.; Yang, T.; Yang, K.; Wang, R.; Yang, J.; Guo, H. Wnt5a Inhibits the Proliferation and Melanogenesis of Melanocytes. Int. J. Med. Sci. 2013, 10, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, K.; Sakai, C.; Kuge, S.; Nishiyama, S.; Tomita, Y.; Ito, S.; Wakamatsu, K.; Hearing, V.J. The Expression of Tyrosinase, Tyrosinase-Related Proteins 1 and 2 (TRP1 and TRP2), the Silver Protein, and a Melanogenic Inhibitor in Human Melanoma Cells of Differing Melanogenic Activities. Pigment. Cell Res. 1995, 8, 97–104. [Google Scholar] [CrossRef]
- Chang, T.-S. Natural Melanogenesis Inhibitors Acting Through the Down-Regulation of Tyrosinase Activity. Materials 2012, 5, 1661–1685. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Yokokawa, Y.; Hatao, M.; Naganuma, M.; Tomita, Y. Comparison of the melanogenesis in human black and light brown melanocytes. J. Dermatol. Sci. 1997, 14, 199–206. [Google Scholar] [CrossRef]
- Lee, R.; Ko, H.J.; Kim, K.; Sohn, Y.; Min, S.Y.; Kim, J.A.; Na, D.; Yeon, J.H. Anti-melanogenic effects of extracellular vesicles derived from plant leaves and stems in mouse melanoma cells and human healthy skin. J. Extracell. Vesicles 2019, 9, 1703480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.Y. Inhibitory effects of geranic acid derivatives on melanin biosynthesis. J. Cosmet. Sci. 2013, 63, 351–358. [Google Scholar]
- Yoon, W.-J.; Kim, M.-J.; Moon, J.-Y.; Kang, H.-J.; Kim, G.-O.; Lee, N.H.; Hyun, C.-G. Effect of Palmitoleic Acid on Melanogenic Protein Expression in Murine B16 Melanoma. J. Oleo. Sci. 2010, 59, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, J.; Jung, E.; Huh, S.; Park, J.-O.; Lee, J.-W.; Byun, S.Y.; Park, D. Mechanisms of depigmentation by α-bisabolol. J. Dermatol. Sci. 2008, 52, 219–222. [Google Scholar] [CrossRef]
- Ando, H.; Funasaka, Y.; Oka, M.; Ohashi, A.; Furumura, M.; Matsunaga, J.; Matsunaga, N.; Hearing, V.J.; Ichihashi, M. Possible involvement of proteolytic degradation of tyrosinase in the regulatory effect of fatty acids on melanogenesis. J. Lipid Res. 1999, 40, 1312–1316. [Google Scholar] [CrossRef]
- Ando, H.; Ryu, A.; Hashimoto, A.; Oka, M.; Ichihashi, M. Linoleic acid and α-linolenic acid lightens ultraviolet-induced hyperpigmentation of the skin. Arch. Dermatol. Res. 1998, 290, 375–381. [Google Scholar] [CrossRef]
- Huh, S.; Kim, Y.-S.; Jung, E.; Lim, J.; Jung, K.S.; Kim, M.-O.; Lee, J.; Park, D. Melanogenesis Inhibitory Effect of Fatty Acid Alkyl Esters Isolated from Oxalis triangularis. Biol. Pharm. Bull. 2010, 33, 1242–1245. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.-C.; Wei, C.-M.; Siao, J.-H.; Tsai, T.-C.; Ko, W.-P.; Chang, K.-J.; Hii, C.-H.; Chang, T.-M. Supercritical Fluid Extract of Spent Coffee Grounds Attenuates Melanogenesis through Downregulation of the PKA, PI3K/Akt, and MAPK Signaling Pathways. Evid. -Based Complement. Altern. Med. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Hosoi, J.; Abe, E.; Suda, T.; Kuroki, T. Regulation of melanin synthesis of B16 mouse melanoma cells by 1 alpha, 25-dihydroxyvitamin D3 and retinoic acid. Cancer Res. 1985, 45, 1474–1478. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Component Name | RT 1 | RI 2 | Area (%) | CAS No. |
|---|---|---|---|---|---|
| 1 | Ethyl 3-ethoxypropionate | 22.93 | 1225 | 0.66 | 763-69-9 |
| 2 | Linalool | 27.75 | 1256 | 0.08 | 78-70-6 |
| 3 | 2-Phenylethanol | 28.53 | 1261 | 0.07 | 60-12-8 |
| 4 | 3,6-Nonadien-1-ol, (E,Z)- | 29.82 | 1269 | 0.16 | 56805-23-3 |
| 5 | Geraniol | 32.46 | 1286 | 0.92 | 106-24-1 |
| 6 | [(3,7-Dimethylocta-2,6-dien-1-yl)oxy] (trimethyl) silane | 34.77 | 1301 | 0.15 | 72237-31-1 |
| 7 | Methyl undecanoate | 36.16 | 1312 | 4.34 | 1731-86-8 |
| 8 | Geranic acid TMS | 36.7 | 1317 | 0.53 | 97779-61-8 |
| 9 | 2-tert-Butyl-4-isopropyl-5-methylphenol | 37.93 | 1327 | 0.10 | None |
| 10 | Undecanoic acid, TMS derivative | 38.55 | 1333 | 0.03 | 146846-83-5 |
| 11 | Dodecanoic acid, trimethylsilyl ester | 40.43 | 1348 | 0.07 | 55520-95-1 |
| 12 | Maalialcohol | 41.42 | 1357 | 0.05 | 527-90-2 |
| 13 | α-Bisabolol | 41.73 | 1359 | 0.36 | 515-69-5 |
| 14 | Tetradecanoic acid, trimethylsilyl ester | 45.34 | 1390 | 3.57 | 18603-17-3 |
| 15 | 2-Heptadecanone | 47.44 | 1413 | 0.05 | 2922-51-2 |
| 16 | n-Pentanoic acid, trimethylsilyl ester | 48.85 | 1435 | 0.15 | 74367-22-9 |
| 17 | Palmitoleic acid | 50.43 | 1458 | 55.41 | 373-49-9 |
| 18 | Palmitelaidic acid, TMS | 53.31 | 1501 | 15.85 | 1206693-35-7 |
| 19 | 6-Pentadecenoic acid,13-methyl-, (6Z)- | 57.57 | 1555 | 0.08 | 682751-34-4 |
| 20 | Linolenic acid | 60.68 | 1594 | 4.02 | 463-40-1 |
| 21 | Ethyl linolenate | 61.88 | 1615 | 0.41 | 1191-41-9 |
| 22 | Oleic acid | 63.88 | 1654 | 0.12 | 112-80-1 |
| 23 | Emulphor | 64.77 | 1671 | 0.08 | 5353-25-3 |
| 24 | 2-Nonadecanone | 67.83 | 1742 | 0.05 | 629-66-3 |
| 25 | Glyceryl palmitate | 69.64 | 1790 | 0.31 | 542-44-9 |
| 26 | 2-Bromooctadecanal | 71.44 | 1848 | 0.03 | 56599-95-2 |
| 27 | Docosanol | 73.04 | 1901 | 3.83 | 661-19-8 |
| 28 | 2-Phenyl-1,3-dithiane | 75.22 | 1985 | 0.61 | 5425-44-5 |
| 29 | 1-Tetracosanol | 77.16 | 2059 | 4.20 | 506-51-4 |
| 30 | 10-Hydroxy-1,6-dimethyl-9-(propan-2-yl)-5,12-dioxatricyclo [9.1.0.04,6]dodecan-8-yl-3-phenylprop-2-enoate (isomer 2) | 78.84 | 2119 | 0.30 | None |
| 31 | (E,E,E,E)-Squalene | 79.09 | 2126 | 0.25 | 7683-64-9 |
| 32 | Dotriacontane | 80.42 | 2168 | 0.08 | 544-85-4 |
| 33 | 22-Tricosenoic acid | 80.54 | 2172 | 1.40 | 65119-95-1 |
| 34 | 17-[5-Hydroxy-6-(2-hydroxypropan-2-yl)oxan-3-yl]-4,4,10,13,14-pentamethyl-1,2,5,6,9,11,12,15,16,17-decahydrocyclopenta[a]phenanthren-3-one, 2TMS | 82.25 | 2220 | 0.58 | None |
| 35 | 9,10 DIDEUTERO OCTADECANAL | 83.09 | 2241 | 0.12 | 56554-44-0 |
| 36 | Tocopherols | 83.58 | 2253 | 0.08 | 7616-22-0 |
| 37 | Quercetin 7,3’,4’-trimethyl ether | 84.07 | 2265 | 0.05 | 6068-80-0 |
| 38 | Benzoic acid;tetracosan-1-ol | 87.9 | 2392 | 0.20 | 103569-99-9 |
| 39 | Betulin | 90.68 | 2570 | 0.20 | 473-98-3 |
| 40 | Lupeol | 91.67 | 2698 | 0.48 | 545-47-1 |
| Total Identified (%) | 100.00 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, Y.R.; Won, K.J.; Kim, D.Y.; Kim, M.J.; Hong, B.S.; Lee, H.M. Chemical Composition of Impatiens textori Miq. Flower Absolute and Its Potential Wound Repair and Anti-Melanogenesis-Promoting Activities in Skin Cells. Pharmaceuticals 2022, 15, 1397. https://doi.org/10.3390/ph15111397
Won YR, Won KJ, Kim DY, Kim MJ, Hong BS, Lee HM. Chemical Composition of Impatiens textori Miq. Flower Absolute and Its Potential Wound Repair and Anti-Melanogenesis-Promoting Activities in Skin Cells. Pharmaceuticals. 2022; 15(11):1397. https://doi.org/10.3390/ph15111397
Chicago/Turabian StyleWon, Yu Rim, Kyung Jong Won, Do Yoon Kim, Mi Jung Kim, Bok Sil Hong, and Hwan Myung Lee. 2022. "Chemical Composition of Impatiens textori Miq. Flower Absolute and Its Potential Wound Repair and Anti-Melanogenesis-Promoting Activities in Skin Cells" Pharmaceuticals 15, no. 11: 1397. https://doi.org/10.3390/ph15111397
APA StyleWon, Y. R., Won, K. J., Kim, D. Y., Kim, M. J., Hong, B. S., & Lee, H. M. (2022). Chemical Composition of Impatiens textori Miq. Flower Absolute and Its Potential Wound Repair and Anti-Melanogenesis-Promoting Activities in Skin Cells. Pharmaceuticals, 15(11), 1397. https://doi.org/10.3390/ph15111397