
Human Lung Cancer (A549) Cell Line Cytotoxicity and Anti-Leishmania major Activity of Carissa macrocarpa Leaves: A Study Supported by UPLC-ESI-MS/MS Metabolites Profiling and Molecular Docking
 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Results
2.1. In-Vitro Cytotoxicity Investigations
2.2. Antileishmanial Activity
2.3. Comparative UPLC-ESI-MS/MS Metabolite Profiling of C. macrocarpa Leaves Total Extract and Its Fractions
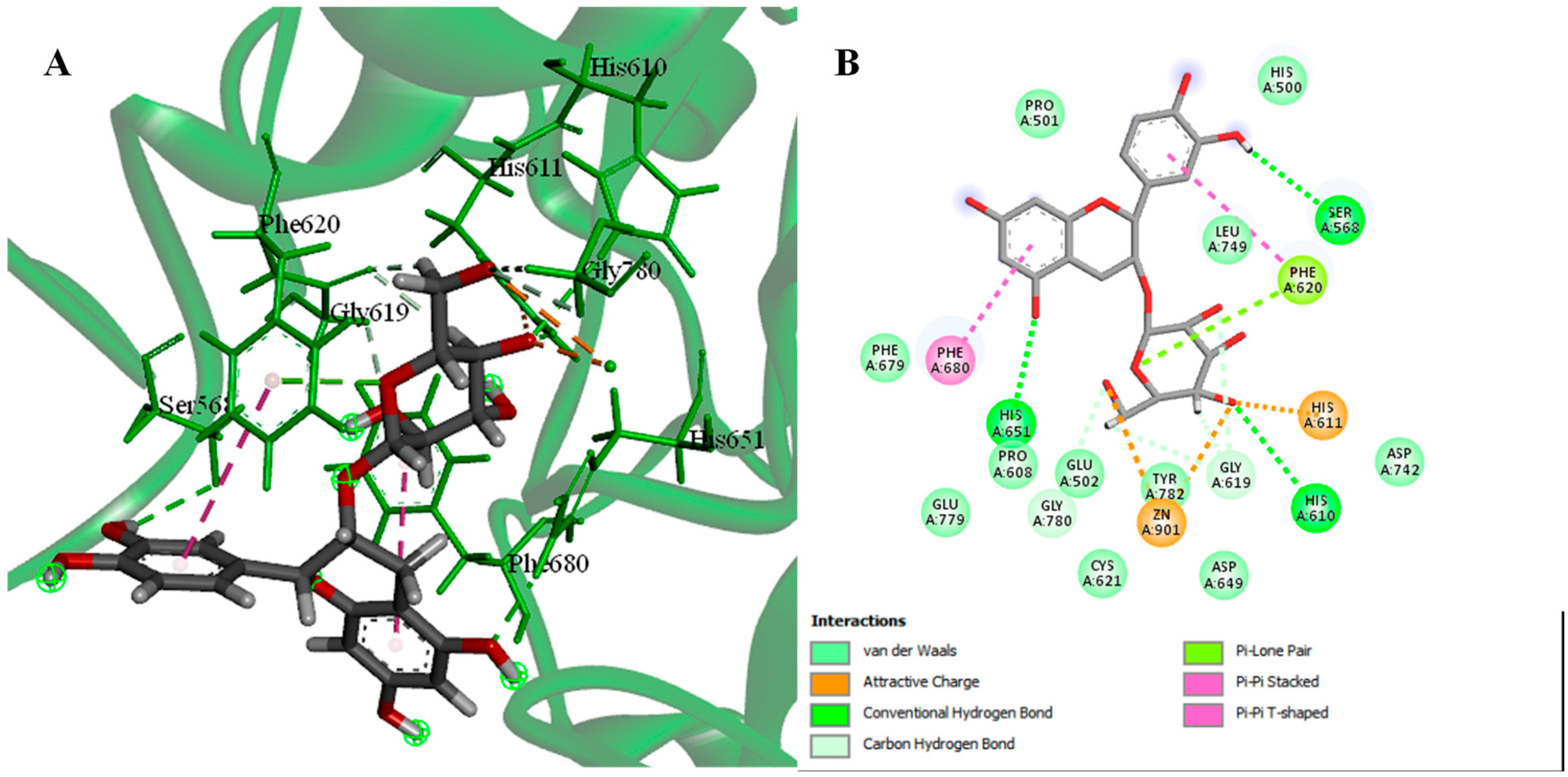
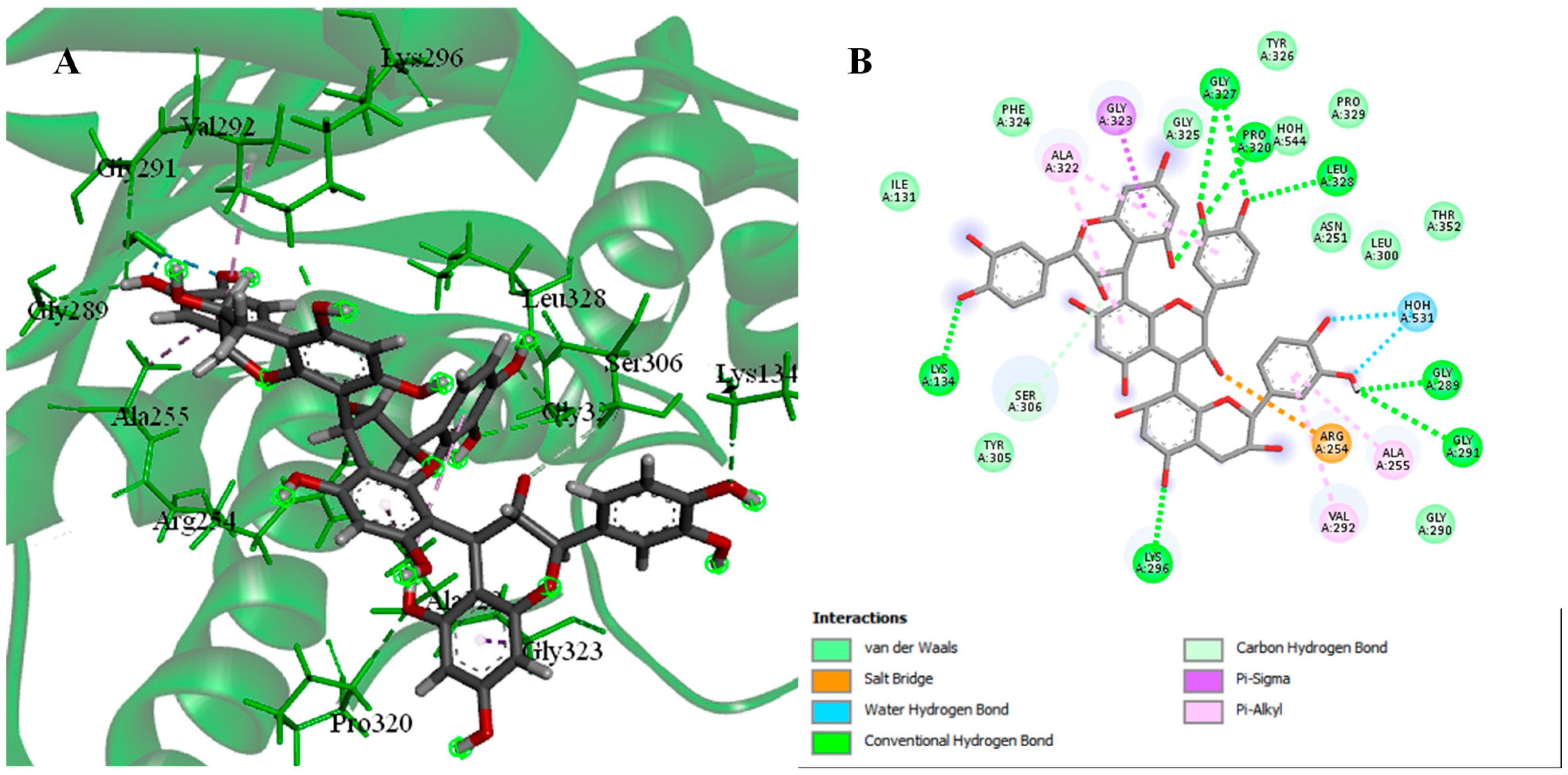
2.4. Molecular Docking
2.4.1. Molecular Docking of Identified Compounds against HDAC6 Enzymes
2.4.2. Molecular Docking of Identified Compounds against PDK3
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Fractionation
4.4. Pharmacological Investigation
4.4.1. Preparation of the Stock Solutions and the Serial Concentrations
4.4.2. Cytotoxicity Investigation
4.4.3. Antileishmanial Promastigotes and Axenic Amastigotes Assays
4.5. UPLC-ESI-MS/MS Investigation
4.6. Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Jazieh, A.R.; Algwaiz, G.; Errihani, H.; Elghissassi, I.; Mula-Hussain, L.; Bawazir, A.A.; Gaafar, R. Lung cancer in the Middle East and North Africa region. J. Thorac. Oncol. 2019, 14, 1884–1891. [Google Scholar] [CrossRef] [PubMed]
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Adverse effects of cancer chemotherapy: Anything new to improve tolerance and reduce sequelae? Front. Media SA 2018, 9, 245. [Google Scholar]
- Mishra, B.B.; Tiwari, V.K. Natural products: An evolving role in future drug discovery. Eur. J. Med. Chem. 2011, 46, 4769–4807. [Google Scholar] [CrossRef] [PubMed]
- Gali-Muhtasib, H.; Hmadi, R.; Kareh, M.; Tohme, R.; Darwiche, N. Cell death mechanisms of plant-derived anticancer drugs: Beyond apoptosis. Apoptosis 2015, 20, 1531–1562. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.A.S.; Lachheb, J.; Chelbi, I.; Louati, D.; Dachraoui, K.; Ben Miled, S.; Zhioua, E. Independent circulation of Leishmania major and Leishmania tropica in their respective sandfly vectors for transmission of zoonotic and chronic cutaneous leishmaniasis co-existing in a mixed focus of central Tunisia. Pathogens 2022, 11, 855. [Google Scholar] [CrossRef]
- Abuzaid, A.A.; Abdoon, A.M.; Aldahan, M.A.; Alzahrani, A.G.; Alhakeem, R.F.; Asiri, A.M.; Alzahrani, M.H.; Memish, Z.A. Cutaneous leishmaniasis in Saudi Arabia: A comprehensive overview. Vector Borne Zoonotic Dis. 2017, 17, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Orabi, M.A.A.; Zidan, S.A.; Sakagami, H.; Murakami, Y.; Ali, A.A.; Alyami, H.S.; Alshabi, A.M.; Matsunami, K. Antileishmanial and lung adenocarcinoma cell toxicity of Withania somnifera (Linn.) dunal root and fruit extracts. Nat. Prod. Res. 2022, 36, 4231–4237. [Google Scholar] [CrossRef] [PubMed]
- Orabi, M.A.A.; Sakagami, H.; Umemura, N.; Alyami, H.S.; Hatano, T. Two new C-glycosidic ellagitannins and accompanying tannins from Lawsonia inermis leaves and their cytotoxic effects. Fitoterapia 2021, 153, 104925. [Google Scholar] [CrossRef] [PubMed]
- Souilem, F.; Dias, M.I.; Barros, L.; Calhelha, R.C.; Alves, M.J.; Harzallah-Skhiri, F.; Ferreira, I.C. Phenolic profile and bioactive properties of Carissa macrocarpa (Eckl.) A. DC.: An in vitro comparative study between leaves, stems, and flowers. Molecules 2019, 24, 1696. [Google Scholar] [CrossRef]
- Souilem, F.; Dias, M.I.; Barros, L.; Calhelha, R.C.; Alves, M.J.; Harzallah-Skhiri, F.; Ferreira, I.C. Amantagula fruit (Carissa macrocarpa (Eckl.) A. DC.): Nutritional and phytochemical characterization. Plant Foods Hum. Nutr. 2019, 74, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Dhatwalia, J.; Kumari, A.; Verma, R.; Upadhyay, N.; Guleria, I.; Lal, S.; Thakur, S.; Gudeta, K.; Kumar, V.; Chao, J.C.-J. Phytochemistry, pharmacology, and nutraceutical profile of Carissa species: An updated review. Molecules 2021, 26, 7010. [Google Scholar] [CrossRef] [PubMed]
- Kaunda, J.S.; Zhang, Y.-J. The genus Carissa: An ethnopharmacological, phytochemical and pharmacological review. Nat. Prod. Bioprospect. 2017, 7, 181–199. [Google Scholar] [CrossRef]
- Patel, S. Food, pharmaceutical and industrial potential of Carissa genus: An overview. Rev. Environ. Sci. Biotechnol. 2013, 12, 201–208. [Google Scholar] [CrossRef]
- Anwar, S.; Mohammad, T.; Shamsi, A.; Queen, A.; Parveen, S.; Luqman, S.; Hasan, G.M.; Alamry, K.A.; Azum, N.; Asiri, A.M. Discovery of Hordenine as a potential inhibitor of pyruvate dehydrogenase kinase 3: Implication in lung cancer therapy. Biomedicines 2020, 8, 119. [Google Scholar] [CrossRef]
- Ojha, R.; Nepali, K.; Chen, C.-H.; Chuang, K.-H.; Wu, T.-Y.; Lin, T.E.; Hsu, K.-C.; Chao, M.-W.; Lai, M.-J.; Lin, M.-H. Isoindoline scaffold-based dual inhibitors of HDAC6 and HSP90 suppressing the growth of lung cancer in vitro and in vivo. Eur. J. Med. Chem. 2020, 190, 112086. [Google Scholar] [CrossRef]
- Gezici, S.; Şekeroğlu, N. Current perspectives in the application of medicinal plants against cancer: Novel therapeutic agents. Curr. Med. Chem. Anticancer Agents 2019, 19, 101–111. [Google Scholar] [CrossRef]
- Hakkim, F.L.; Bakshi, H.A.; Khan, S.; Nasef, M.; Farzand, R.; Sam, S.; Rashan, L.; Al-Baloshi, M.S.; Hasson, S.S.A.A.; Al Jabri, A. Frankincense essential oil suppresses melanoma cancer through down-regulation of Bcl-2/Bax cascade signalling and ameliorates hepatotoxicity via phase I and II drug-metabolizing enzymes. Oncotarget 2019, 10, 3472. [Google Scholar] [CrossRef]
- Menke, K.; Schwermer, M.; Schramm, A.; Zuzak, T.J. Preclinical evaluation of antitumoral and cytotoxic properties of Viscum album fraxini extract on pediatric tumour cells. Planta Med. 2019, 85, 1150–1159. [Google Scholar]
- Khalil, H.E.; Mohamed, M.E.; Morsy, M.A.; Kandeel, M. Flavonoid and phenolic compounds from Carissa macrocarpa: Molecular docking and cytotoxicity studies. Pharmacogn. Mag. 2018, 14, 304. [Google Scholar] [CrossRef]
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host-pathogen interface. Nat. Rev. Microbiol. 2011, 9, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Ben Said, R.; Hamed, A.I.; Mahalel, U.A.; Al-Ayed, A.S.; Kowalczyk, M.; Moldoch, J.; Oleszek, W.; Stochmal, A. Tentative characterization of polyphenolic compounds in the male flowers of Phoenix dactylifera by liquid chromatography coupled with mass spectrometry and DFT. Int. J. Mol. Sci. 2017, 18, 512. [Google Scholar] [CrossRef]
- Jaiswal, R.; Matei, M.F.; Subedi, P.; Kuhnert, N. Does roasted coffee contain chlorogenic acid lactones or/and cinnamoyl shikimate esters? Food Res. Int. 2014, 61, 214–227. [Google Scholar] [CrossRef]
- Alakolanga, A.; Siriwardane, A.; Kumar, N.S.; Jayasinghe, L.; Jaiswal, R.; Kuhnert, N. LC-MSn identification and characterization of the phenolic compounds from the fruits of Flacourtia indica (Burm. F.) Merr. and Flacourtia inermis Roxb. Food Res. Int. 2014, 62, 388–396. [Google Scholar] [CrossRef]
- Pereira, E.; Pimenta, A.I.; Calhelha, R.C.; Antonio, A.L.; Verde, S.C.; Barros, L.; Santos-Buelga, C.; Ferreira, I.C. Effects of gamma irradiation on cytotoxicity and phenolic compounds of Thymus vulgaris L. and Mentha x piperita L. LWT—Food Sci. Technol. 2016, 71, 370–377. [Google Scholar] [CrossRef]
- Yuzuak, S.; Ballington, J.; Xie, D.-Y. HPLC-qTOF-MS/MS-based profiling of flavan-3-ols and dimeric proanthocyanidins in berries of two muscadine grape hybrids FLH 13-11 and FLH 17-66. Metabolites 2018, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Jayasinghe, L.; Kuhnert, N. Identification and characterization of proanthocyanidins of 16 members of the Rhododendron genus (Ericaceae) by tandem LC–MS. J. Mass Spectrom. 2012, 47, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.-Y.; Yang, Y.-F.; Li, K. Analysis of hydroxy fatty acids from the pollen of Brassica campestris L. var. oleifera DC. by UPLC-MS/MS. J. Pharm. 2013, 2013, 874875. [Google Scholar] [CrossRef] [PubMed]
- May, A.; El-Sayed, A.A.A.-G.; Hamdan, D.I.; El-Shazly, A.M. Phytoconstituents, LC-ESI-MS profile, antioxidant and antimicrobial activities of Citrus x limon L. Burm. f. cultivar variegated pink lemon. J. Pharm. Sci. Res. 2017, 9, 375–391. [Google Scholar]
- Hussain, S.A.; Hameed, A.; Nazir, Y.; Naz, T.; Wu, Y.; Suleria, H.A.R.; Song, Y. Microencapsulation and characterization of natural polyphenols from PHF Extract. Nutrients 2018, 10, 843. [Google Scholar] [CrossRef] [PubMed]
- Moodley, R.; Koorbanally, N.; Jonnalagadda, S.B. Elemental composition and fatty acid profile of the edible fruits of Amatungula (Carissa macrocarpa) and impact of soil quality on chemical characteristics. Anal. Chim. Acta 2012, 730, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Petta, V.; Gkiozos, I.; Strimpakos, A.; Syrigos, K. Histones and lung cancer: Are the histone deacetylases a promising therapeutic target? Cancer Chemother. Pharmacol. 2013, 72, 935–952. [Google Scholar] [CrossRef]
- Zhou, Y.-X.; Xin, H.-L.; Rahman, K.; Wang, S.-J.; Peng, C.; Zhang, H. Portulaca oleracea L.: A review of phytochemistry and pharmacological effects. BioMed Res. Int. 2015, 2015, 925631. [Google Scholar] [CrossRef] [PubMed]
- Tala, V.R.S.; da Silva, V.C.; Rodrigues, C.M.; Nkengfack, A.E.; dos Santos, L.C.; Vilegas, W. Characterization of Proanthocyanidins from Parkia biglobosa (Jacq.) G. Don. (Fabaceae) by Flow Injection Analysis—Electrospray ionization ion trap tandem mass spectrometry and liquid chromatography/electrospray ionization mass spectrometry. Molecules 2013, 18, 2803–2820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Deng, W.-W.; Wan, X.-C. Advantage of LC-MS metabolomics to identify marker compounds in two types of Chinese dark tea after different post-fermentation processes. Food Sci. Biotechnol. 2014, 23, 355–360. [Google Scholar] [CrossRef]
- Li, J.; Kuang, G.; Chen, X.; Zeng, R. Identification of chemical composition of leaves and flowers from Paeonia rockii by UHPLC-Q-Exactive Orbitrap HRMS. Molecules 2016, 21, 947. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Chen, P.; Harnly, J.M. New phenolic components and chromatographic profiles of green and fermented teas. J. Agric. Food Chem. 2008, 56, 8130–8140. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. HPLC–DAD–ESI-MS/MS screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef]
- Yu, J.-Q.; Xian, Y.-X.; Geng, Y.-L.; Wang, D.-J.; Zhou, H.-L.; Wang, X. Anti-liver cancer constituents from the thorns of Gleditsia sinensis. Phytochem. Lett. 2015, 13, 343–347. [Google Scholar] [CrossRef]
- Izhar, S.; Ahmed, D. Carissa opaca: A plant with great potential for future drugs for degenerative and infectious diseases. ChemistrySelect 2016, 1, 3005–3011. [Google Scholar] [CrossRef]
- Orabi, M.A.A.; Khalil, H.M.; Abouelela, M.E.; Zaafar, D.; Ahmed, Y.H.; Naggar, R.A.; Alyami, H.S.; Abdel-Sattar, E.-S.; Matsunami, K.; Hamdan, D.I. Carissa macrocarpa leaves polar fraction ameliorates doxorubicin-induced neurotoxicity in rats via downregulating the oxidative stress and inflammatory markers. Pharmaceuticals 2021, 14, 1305. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Alhumaydhi, F.A.; Alsahli, M.A.; Rahmani, A.H. Potential therapeutic targets of epigallocatechin gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules 2020, 25, 3146. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives, and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef]
- Arora, I.; Sharma, M.; Tollefsbol, T.O. Combinatorial epigenetics impact of polyphenols and phytochemicals in cancer prevention and therapy. Int. J. Mol. Sci. 2019, 20, 4567. [Google Scholar] [CrossRef]
- Carlos-Reyes, Á.; López-González, J.S.; Meneses-Flores, M.; Gallardo-Rincón, D.; Ruíz-García, E.; Marchat, L.A.; Astudillo-De La Vega, H.; de la Cruz, O.N.H.; López-Camarillo, C. Dietary compounds as epigenetic modulating agents in cancer. Front. Genet. 2019, 10, 79. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- da Silva, E.R.; Brogi, S.; Grillo, A.; Campiani, G.; Gemma, S.; Vieira, P.C.; Maquiaveli, C.D.C. Cinnamic acids derived compounds with antileishmanial activity target Leishmania amazonensis arginase. Chem. Biol. Drug Des. 2019, 93, 139–146. [Google Scholar] [CrossRef]
- Montrieux, E.; Perera, W.H.; García, M.; Maes, L.; Cos, P.; Monzote, L. In vitro and in vivo activity of major constituents from Pluchea carolinensis against Leishmania amazonensis. Parasitol. Res. 2014, 113, 2925–2932. [Google Scholar] [CrossRef]
- Elamin, M.H.; Al-Maliki, S.S. Leishmanicidal and apoptotic activities of oleuropein on Leishmania major. Int. J. Clin. Pharm. Ther. 2014, 52, 880–888. [Google Scholar] [CrossRef]
- Kyriazis, I.; Smirlis, D.; Papadaki, A.; Koutsoni, O.; Aligiannis, N.; Skaltsounis, A. Leishmanicidal activity of oleuropein: Leishmania donovani promastigote cell death through a possibly ROS-independent mechanism. J. Pharm. Nat. Prod. 2017, 3, 141. [Google Scholar] [CrossRef]
- Passero, L.F.; Cruz, L.A.; Santos-Gomes, G.; Rodrigues, E.; Laurenti, M.D.; Lago, J.H.G. Conventional versus natural alternative treatments for leishmaniasis: A review. Curr. Top. Med. Chem. 2018, 18, 1275–1286. [Google Scholar] [CrossRef]
- Yamamoto, E.S.; Campos, B.L.; Jesus, J.A.; Laurenti, M.D.; Ribeiro, S.P.; Kallás, E.G.; Rafael-Fernandes, M.; Santos-Gomes, G.; Silva, M.S.; Sessa, D.P. The effect of ursolic acid on Leishmania (Leishmania) amazonensis is related to programed cell death and presents therapeutic potential in experimental cutaneous leishmaniasis. PLoS ONE 2015, 10, e0144946. [Google Scholar] [CrossRef] [PubMed]
- Jesus, J.A.; Fragoso, T.N.; Yamamoto, E.S.; Laurenti, M.D.; Silva, M.S.; Ferreira, A.F.; Lago, J.H.G.; Gomes, G.S.; Passero, L.F.D. Therapeutic effect of ursolic acid in experimental visceral leishmaniasis. Int. J. Parasitol. Drugs Drug Res. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Ghosh, S.; De, A.K.; Bera, T. Oral delivery of ursolic acid-loaded nanostructured lipid carrier coated with chitosan oligosaccharides: Development, characterization, in vitro and in vivo assessment for the therapy of leishmaniasis. Int. J. Biol. Macromol. 2017, 102, 996–1008. [Google Scholar] [CrossRef] [PubMed]
- Moraes Neto, R.N.; Setúbal, R.F.B.; Higino, T.M.M.; Brelaz-de-Castro, M.C.A.; da Silva, L.C.N.; Aliança, A.S.D.S. Asteraceae plants as sources of compounds against leishmaniasis and Chagas disease. Front. Pharmacol. 2019, 10, 477. [Google Scholar] [CrossRef] [PubMed]
- Zadeh Mehrizi, T.; Shafiee Ardestani, M.; Haji Molla Hoseini, M.; Khamesipour, A.; Mosaffa, N.; Ramezani, A. Novel nanosized chitosan-betulinic acid against resistant Leishmania major and first clinical observation of such parasite in kidney. Sci. Rep. 2018, 8, 11759. [Google Scholar] [CrossRef]
- Ribeiro, T.G.; Nascimento, A.M.; Henriques, B.O.; Chávez-Fumagalli, M.A.; Franca, J.R.; Duarte, M.C.; Lage, P.S.; Andrade, P.H.; Lage, D.P.; Rodrigues, L.B. Antileishmanial activity of standardized fractions of Stryphnodendron obovatum (Barbatimão) extract and constituent compounds. J. Ethnopharmacol. 2015, 165, 238–242. [Google Scholar] [CrossRef]
- Sosa, A.M.; Moya Álvarez, A.; Bracamonte, E.; Korenaga, M.; Marco, J.D.; Barroso, P.A. Efficacy of topical treatment with (−)-epigallocatechin gallate, a green tea catechin, in mice with cutaneous leishmaniasis. Molecules 2020, 25, 1741. [Google Scholar] [CrossRef]
- Carter, N.S.; Stamper, B.D.; Elbarbry, F.; Nguyen, V.; Lopez, S.; Kawasaki, Y.; Poormohamadian, R.; Roberts, S.C. Natural products that target the arginase in Leishmania parasites hold therapeutic promise. Microorganisms 2021, 9, 267. [Google Scholar] [CrossRef]
- Keshav, P.; Goyal, D.K.; Kaur, S. Antileishmanial potential of immunomodulator gallic acid against experimental murine visceral leishmaniasis. Parasite Immunol. 2021, 43, e12875. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.M.D.M.; Arcanjo, D.D.R.; Figueiredo, K.A.; Oliveira, J.S.D.S.M.; Viana, F.J.C.; Coelho, E.D.S.; Lopes, G.L.N.; Gonçalves, J.C.R.; Carvalho, A.L.M.; Rizzo, M.D.S. Gallic and ellagic acids are promising adjuvants to conventional amphotericin B for the treatment of cutaneous leishmaniasis. Antimicrob. Agents Chemother. 2020, 64, e00807–e00820. [Google Scholar] [CrossRef] [PubMed]
- Belkhelfa-Slimani, R.; Djerdjouri, B. Caffeic acid and quercetin exert caspases-independent apoptotic effects on Leishmania major promastigotes, and reactivate the death of infected phagocytes derived from BALB/c mice. Asian Pac. J. Trop. Biomed. 2017, 7, 321–331. [Google Scholar] [CrossRef]
- Saini, S.; Rai, A.K. Linoleic acid inhibits the release of Leishmania donovani derived microvesicles and decreases its survival in macrophages. Front. Cell. Infect. Microbiol. 2020, 10, 406. [Google Scholar] [CrossRef]
- Ranjbar, R.; Shayanfar, P.; Maniati, M. In Vitro Antileishmanial Effects of Saffron Compounds, Crocin and Stigmasterol, on Iranian Strain of Leishmania major (MHOM/IR/75/ER). Iran. J. Parasitol. 2021, 16, 151. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, P.K.; Chakraborti, S.; Bagchi, A.; Chakraborti, T. Bioassay-based Corchorus capsularis L. leaf-derived β-sitosterol exerts antileishmanial effects against Leishmania donovani by targeting trypanothione reductase. Sci. Rep. 2020, 10, 20440. [Google Scholar] [CrossRef]
- Shah, S.M.; Ullah, F.; Ayaz, M.; Sadiq, A.; Hussain, S.; Shah, S.A.A.; Nadhman, A. β-Sitosterol from Ifloga spicata (Forssk.) Sch. Bip. as potential anti-leishmanial agent against Leishmania tropica: Docking and molecular insights. Steroids 2019, 148, 56–62. [Google Scholar] [CrossRef]
- Callahan, H.L.; Portal, A.C.; Devereaux, R.; Grogl, M. An axenic amastigote system for drug screening. Antimicrob. Agents Chemother. 1997, 41, 818–822. [Google Scholar] [CrossRef]
- Ephros, M.; Waldman, E.; Zilberstein, D. Pentostam induces resistance to antimony and the preservative chlorocresol in Leishmania donovani promastigotes and axenically grown amastigotes. Antimicrob. Agents Chemother. 1997, 41, 1064–1068. [Google Scholar] [CrossRef]
- Matteucci, K.C.; Pereira, G.J.; Weinlich, R.; Bortoluci, K.R. Frontline Science: Autophagy is a cell autonomous effector mechanism mediated by NLRP3 to control Trypanosoma cruzi infection. J. Leukoc. Biol. 2019, 106, 531–540. [Google Scholar] [CrossRef]
- Hamdan, D.I.; Fayed, M.A.; Adel, R. Echinops taeckholmiana Amin: Optimization of a tissue culture protocol, biological evaluation, and chemical profiling using GC and LC-MS. ACS Omega 2021, 6, 13105–13115. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Keefover-Ring, K.; Ahnlund, M.; Abreu, I.N.; Jansson, S.; Moritz, T.; Albrectsen, B.R. No evidence of geographical structure of salicinoid chemotypes within Populus tremula. PLoS ONE 2014, 9, e107189. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Q.-S.; Xu, L.-L.; Zhang, J.-Y.; Wang, Z.-J.; Jiang, Y.-Y.; Liu, B. Rapid characterization and identification of non-diterpenoid constituents in Tinospora sinensis by HPLC-LTQ-orbitrap MSn. Molecules 2018, 23, 274. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cytotoxicity | Anti-L. major Activity | ||||
|---|---|---|---|---|---|
| Sample | A549 Cell Line | Promastigotes | Axenic Amastigotes | ||
| IC50 ± SE (μg/mL) | % Inhibition ± SE (at 100 μg/mL) | IC50 ± SE (μg/mL) | % Inhibition ± SE (at 100 μg/mL) | IC50 ± SE (μg/mL) | |
| Total ext. | 3.3 ± 0.19 | 33.21 ± 2.39 | >100 | 7.42 ± 1.4 | >100 |
| MeOH fr. | 1.57 ± 0.04 | nil | >100 | nil | >100 |
| Butanol Fr. | 6.16 ± 0.35 | 61.15 ± 0.86 | 73.17 ± 1.62 | nil | >100 |
| EtOAc fr. | 50.66 ± 1.95 | 88.59 ± 1.68 | 27.52 ± 0.7 | 29.33 ± 4.86 | >100 |
| Hexane fr. | >100 | nil | >100 | nil | >100 |
| Etoposide | 50.08 ± 3.16 | - | - | - | - |
| Sodium stibogluconate | - | - | - | 100 | 22.45 ± 2.22 |
| Miltefosine | - | 100 | 6.39 ± 0.29 | 100 | 12.35 ± 1.8 |
| Sample | Weight (g) | % In the Total Extract | % In the Dry Powder |
|---|---|---|---|
| Total ext. | 66.6 | 100% | 26.64% |
| MeOH fr. | 47.2 | 70.87% | 18.88% |
| Butanol Fr. | 3.63 | 5.45% | 1.452% |
| EtOAc fr. | 5.52 | 8.29% | 2.208% |
| Hexane fr. | 0.2 | 0.3% | 0.08% |
| Peak No. | Rt | Molecular Weight | MS [M − H]−/[M + H]+ | MS2 | Tentatively Identified Compound | Reference | Class | Total | EtOAc fr. | Butanol fr. | MeOH fr. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.74 | 378 | 377.1617/379.2518 | 333, 271, 257, 163, 119 | Carinol | [15] | Miscellaneous | √ | - | √ | √ |
| 2 | 0.75 | 192 | 191.0583/193.1359 | 173 | Quinic acid | [23] | Phenolic acid | - | √ | - | - |
| 3 | 0.80 | 234 | 233.1027/- | 214, 164, 134 | Dehydrocarissone (11-hydroxy-1,4-eudesmadien-3-one) | [14,15] | Miscellaneous | - | √ | - | - |
| 4 | 0.85 | 240 | 239.0533/- | 221, 203, 188, 173, 143 | Cryptomeridiol | [14,15] | Miscellaneous | - | - | √ | - |
| 5 | 0.96 | 418 | -/419.2318 | 386, 359, 356, 255 | 3′-(4″-methoxyphenyl)-3′-oxo-propionyl hexadecanoate | [15] | Fatty acid | √ | - | - | - |
| 6 | 1.87 | 354 | 353.1642/355.1283 | 191,179,161 | 3-O-Caffeolyquinic acid | [11] | Phenolic acid | √ | - | - | √ |
| 7 | 2.03 | 354 | 353.1728/- | 191,173,161 | 4-O-Caffeolyquinic acid | [11] | Phenolic acid | - | - | - | √ |
| 8 | 2.13 | 326 | 325.1349/- | 187, 163, 145 | Coumaroyl-β-glucose | [24,25] | Phenolic acid | - | - | √ | √ |
| 9 | 2.161 | 354 | 353.2025/- | 191,179,161 | 5-O-Caffeolyquinic acid | [11] | Phenolic acid | - | - | √ | √ |
| 10 | 2.24 | 578 | 577.2717/- | 425, 289 | Type B (epi)catechin dimer | [11] | Flavan-3-ol | √ | - | √ | √ |
| 11 | 2.45 | 866 | 865.4962/- | 451, 425, 407, 289 | Type B (epi)catechin trimer | [11] | Flavan-3-ol | √ | - | √ | √ |
| 12 | 2.85 | 320 | 319.1711/- | 301, 275, 257, 231, 203,163, 119 | 5-O-p-Coumaroylshikimic acid | [24] | Phenolic acid | √ | - | √ | √ |
| 13 | 2.87 | 342 | -/343.1820 | 326, 311, 285 | Caffeic acid 3-glucoside | [26] | Phenolic acid | √ | - | √ | √ |
| 14 | 2.87 | 452 | 451.3018/- | 408, 393, 351, 337, 301, 273, 245 | Catechin-3-O-glucoside | [27] | Flavan-3-ol | - | - | - | √ |
| 15 | 2.87 | 320 | 319.1711/- | 275, 257, 199, 163, 119 | 4-O-p-Coumaroylshikimic acid | [24] | Phenolic acid | √ | - | √ | √ |
| 16 | 2.87 | 452 | 451.3018/- | 391, 343, 301, 287, 273, 247 | Epicatechin-3-O-glucoside | [27] | Flavan-3-ol | - | - | - | √ |
| 17 | 3.00 | 290 | 289.0894/- | 245, 205, 203, 187, 179, 161 | (epi) Catechin | [27] | Flavan-3-ol | √ | - | √ | √ |
| 18 | 5.05 | 756 | 755.4382/- | 593, 285 | Kaempferol-7-O-hexoside-3-O-rutinoside | [11] | Flavonoid | √ | - | √ | √ |
| 19 | 5.12 | 756 | 755.5181/- | 609, 301 | Quercetin-7-O-deoxyhexoside-3-O-deoxyhexosyl-hexoside | [11] | Flavonoid | √ | - | √ | √ |
| 20 | 5.14 | 452 | 451.3743/- | 391, 343, 301, 287, 273, 247 | Epicatechin-3-O-glucoside isomer | [27] | Flavan-3-ol | √ | - | √ | √ |
| 21 | 5.52 | 740 | 739.3967/- | 593, 285 | Kaempferol-7-O-deoxyhexoside-3-O-deoxyhexosyl-hexoside isomer 1 | [11] | Flavonoid | √ | - | √ | √ |
| 22 | 5.59 | 740 | 739.4193/- | 593, 285 | Kaempferol-7-O-deoxyhexoside-3-O-deoxyhexosyl-hexoside isomer 2 | [11] | Flavonoid | - | - | - | √ |
| 23 | 5.59 | 610 | 609.2661/- | 301 | Quercetin- 3-O-deoxyhexosyl-hexoside isomer 1 | [11] | Flavonoid | - | - | - | √ |
| 24 | 5.75 | 610 | 609.3278/611.2927 | 465, 303 | Quercetin- 3-O-deoxyhexosyl-hexoside isomer 2 | [11] | Flavonoid | √ | - | √ | √ |
| 25 | 5.80 | 578 | 577.2878/- | 425, 289 | Type B (epi)catechin dimer | [11] | Flavan-3-ol | - | - | √ | √ |
| 26 | 5.90 | 450 | 449.1666/- | 317, 316 | Myricetin-3-O-xyloside | [23,28] | Flavonoid | - | - | √ | √ |
| 27 | 6.04 | 300 | -/302.8930 | 275, 257, 229, 215, 153 | Quercetin | [23] | Flavonoid | - | - | √ | √ |
| 28 | 6.13 | 594 | 593.3281/- | 557, 467, 441, 425, 407, 289 | (epi) Gallocatechin-(epi)catechin | [28] | Flavan-3-ol | - | - | √ | √ |
| 29 | 6.58 | 516 | 515.2966/- | 353, 179 | Dicaffeoylquinic acid | [24] | Phenolic acid | √ | - | √ | √ |
| 30 | 6.80 | 138 | 136.9441/- | 109, 93 | Hydroxy benzoic acid | [23] | Phenolic acid | - | - | √ | √ |
| 31 | 6.87 | 188 | 187.1353/- | 169, 125 | Gallic acid monohydrate | [23] | Phenolic acid | - | √ | - | - |
| 32 | 6.87 | 194 | 193.1353/- | 169, 125 | Ferulic acid | [23] | Phenolic acid | - | √ | - | - |
| 33 | 7.01 | 594 | 593.4289/- | 557, 467, 441, 425, 407, 289 | (epi) Gallocatechin-(epi)catechin | [28] | Flavan-3-ol | - | - | √ | - |
| 34 | 8.11 | 180 | 178.8018/- | 179, 135 | Caffeic acid | [23] | Phenolic acid | - | √ | √ | - |
| 35 | 8.12 | 198 | 196.9333/- | 120, 104, 93, 87 | Syringic acid | [23] | Phenolic acid | - | - | - | √ |
| 36 | 8.71 | 328 | 327.2579/- | 281, 279, 255, 213, 183 | Trihydroxy-octadecadienoic acid | [29] | Fatty acid | √ | √ | - | - |
| 37 | 9.14 | 574 | 573.6962/- | 397, 223, 173 | Feruloyl-O-sinapoylquinic acid | [24] | Phenolic acid | √ | - | - | √ |
| 38 | 9.30 | 330 | 329.2814/- | 311, 293, 229, 211, 171, 143 | Trihydroxy-octadecenoic acid | [29] | Fatty acid | √ | - | - | - |
| 39 | 9.94 | 940 | 939.0552/- | 778, 735, 732, 717, 571 | Diacetoxy-5-methoxyphenyl acroyl-O-p-coumaroyl-O-caffeoylquinic acid derivative | [23] | Phenolic acid | √ | - | √ | √ |
| 40 | 10.21 | 310 | 309.2307/- | 291, 279, 251, 223, 221, 89 | Dihydroxy-octadecadienoic acid | [29] | Fatty acid | - | √ | - | - |
| 41 | 10.21 | 378 | 377.1844/- | 345, 327 | Oleuropein aglycone | [15] | Miscellaneous | - | √ | - | - |
| 42 | 10.22 | 342 | 341.2001/- | 326, 311, 285 | Tetramethoxyflavone | [30] | Flavonoid | - | - | √ | - |
| 43 | 10.29 | 310 | 309.2261/- | 291, 279, 251, 223, 221, 89 | Dihydroxy-octadecatrienoic acid isomer | [29] | Fatty acid | - | √ | - | - |
| 44 | 11.21 | 344 | 343.2618/- | 191, 169 | Galloylquinic acid | [23] | Phenolic acid | - | - | √ | - |
| 45 | 11.40 | 306 | 305.1978/- | 261, 219, 221, 179, 165, 125 | (epi)-Gallocatechin | [28] | Flavan-3-ol | - | √ | - | - |
| 46 | 11.54 | 294 | 293.1970/- | 275, 251, 221, 207 | Monohydroxy-Octadecatrienoic acid | [29] | Fatty acid | - | √ | - | - |
| 47 | 12.06 | 176 | 174.9668/- | 157 | Ascorbic acid | [23] | Phenolic acid | - | - | √ | - |
| 48 | 12.60 | 288 | 287.3836/- | 151, 135, 125, 107 | Eriodictyol | [30] | Flavonoid | - | - | √ | - |
| 49 | 12.78 | 436 | -/437.2773 | 307, 181 | Epigallocatechin-3-O-cinnamate | [31] | Flavan-3-ol | - | - | √ | - |
| 50 | 12.98 | 472 | 471.4764/- | 441, 407, 313, 303, 287, 269, 257, 243, 161, | Methyl-3-O-gallocatechin gallate | [27] | Flavan-3-ol | - | √ | √ | - |
| 51 | 12.98 | 454 | -/455.3868 | 439, 411, 393, 248, 207, 203, 191, 189 | 3β-hydroxyolean-11-en-28,13β-olide | [14,32] | Triterpene | - | √ | - | - |
| 52 | 13.11 | 414 | 413.3687/- | 366, 270, 255, 189, 175, 161 | β-Sitosterol | [33,34] | Sterol | - | √ | - | - |
| 53 | 13.26 | 578 | 577.4777/- | 425, 289 | (epi) Catechin dimer | [11] | Flavan-3-ol | - | - | - | √ |
| 54 | 13.30 | 544 | 543.5621/- | 353, 173 | Dimethoxycinnamoyl-O-caffeoylquinic acid | [23] | Phenolic acid | - | - | - | √ |
| 55 | 13.35 | 312 | 311.2344/- | 293, 275, 253, 235, 223 | Dihydroxy-octadecadienoic acid | [29] | Fatty acid | - | √ | - | - |
| 56 | 13.78 | 472 | 471.4489/- | 441, 407, 303, 288, 257, 243, 201, 169, 161 | Methyl-3-O- epigallocatechin gallate | [27] | Flavan-3-ol | - | √ | √ | - |
| 57 | 14.37 | 634 | 633.4683/- | 481, 305 | (epi)Gallocatechin-O-gallate-O-glucuronide | [27] | Flavan-3-ol | - | - | √ | - |
| 58 | 14.55 | 432 | 431.1743/- | 341, 311 | Vitexin | [23] | Flavonoid | - | √ | - | - |
| 59 | 14.69 | 560 | 559.4719/- | 397, 223 | 3-O Caffeoyl-4-O-sinapoylquinic acid | [24] | Phenolic acid | - | - | √ | - |
| 60 | 14.79 | 560 | 559.6325/- | 397, 223 | 3-O-Sinapoyl-4-O-caffeoylquinic acid | [24] | Phenolic acid | - | - | √ | - |
| 61 | 14.82 | 472 | 471.4497/- | 441, 407, 297, 269, 241, 213, 199, 168, 161 | Methyl-3-O-epigallocatechin gallate isomer | [27] | Flavan-3-ol | - | √ | √ | - |
| 62 | 15.03 | 472 | 471.4144/- | 453, 435, 407, 389 | 23-Hydroxybetulinic acid | [14] | Sterol | - | √ | - | - |
| 63 | 15.06 | 560 | 559.4719/- | 397, 223 | 1-O-Caffeoyl-3-O-sinapoylquinic acid | [24] | Phenolic acid | - | - | √ | - |
| 64 | 15.06 | 354 | 353.3186/- | 179,161 | 4-O-Caffeoylshikimic acid | [24] | Phenolic acid | - | - | √ | - |
| 65 | 15.30 | 618 | 617.6021/- | 599, 465, 289 | (epi) Catechin-O-gallate-O-glucuronide | [35] | Flavan-3-ol | √ | - | √ | - |
| 66 | 15.28 | 296 | 295.2867/ | 277, 253, 223, 167 | 13-hydroxyoctadec-2-enoic acid | [29] | Fatty acid | - | √ | - | - |
| 67 | 15.30 | 470 | -/471.3879 | 456, 439, 411, 393, 248, 207, 203, 191, 189 | Methyloleanolate | [14] | Triterpene | - | √ | - | - |
| 68 | 15.46 | 648 | 647.6069/- | 485, 470, 455, 440 | 3,27-Dihydroxy-12-ursen-28-oic acid; 3β-form, 27-(4-Hydroxy-3-methoxycinnamoyl) (E-form) | [14] | Triterpene | - | √ | √ | - |
| 69 | 15.48 | 454 | -/455.4161 | 307, 179, 137 | Gallocatechin derivative | [36] | Flavan-3-ol | - | √ | √ | - |
| 70 | 15.66 | 544 | 543.3333/- | 353, 173 | Dimethoxycinnamoyl-O-caffeoylquinic acid isomer | [23] | Phenolic acid | - | - | - | √ |
| 71 | 15.70 | 618 | 617.5588/- | 599, 465, 289 | (epi)Catechin-O-gallate-O-glucuronide isomer | [35] | Flavan-3-ol | - | - | √ | - |
| 72 | 15.70 | 454 | -/455.4180 | 307, 179, 137 | Gallocatechin derivative | [36] | Flavan-3-ol | - | √ | √ | - |
| 73 | 15.92 | 646 | 647.5842/- | 485, 470, 455, 440 | 27-Coumaroyloxyursolic acid | [23] | Triterpene | - | √ | - | - |
| 74 | 16.52 | 354 | 352.9919/- | 179,161 | 3-O-Caffeoylshikimic acid | [24] | Phenolic acid | - | - | - | √ |
| 75 | 16.59 | 382 | -/383.2513 | 369, 351, 195 | Dimethoxycinnamoylquinic acid | [24] | Phenolic acid | √ | - | √ | √ |
| 76 | 16.59 | 326 | 325.1193/- | 187, 163, 145 | Coumaric acid hexoside | [25] | Phenolic acid | - | - | √ | - |
| 77 | 17.08 | 338 | 337.1823/- | 202, 190, 163 | p-Coumaroylquinic acid | [23] | Phenolic acid | - | - | √ | - |
| 78 | 17.55 | 382 | 381.2726/- | 367, 349, 193 | Dimethoxycinnamoylquinic acid | [24] | Phenolic acid | - | - | √ | - |
| 79 | 18.45 | 600 | -/601.5302 | 447, 313, 285, 284, 169, 151, 125 | Kaempferol galloylglucoside | [37] | Flavonoid | √ | - | - | - |
| 80 | 18.58 | 456 | 455.4745/- | 439, 419, 411, 410, 407, 397 | Ursolic acid | [28,32] | Triterpene | √ | √ | - | - |
| 81 | 19.06 | 456 | 455.4576/- | 439, 419, 411, 410, 407, 397 | Carissic acid (isomer of ursolic acid) | [28,32] | Triterpene | √ | √ | √ | - |
| 82 | 19.16 | 456 | 455.4955/457.4337 | 439, 419, 411, 410, 407, 397 | Oleanolic acid | [28,32] | Triterpene | √ | √ | √ | - |
| 83 | 20.28 | 340 | 339.3065/- | 295, 251, 179 | Caffeoyl-2-hydroxyethane-1,1,2-tricarboxylic acid | [23] | Phenolic acid | - | √ | √ | - |
| 84 | 20.73 | 376 | 375.3291/- | 361, 347, 294, 123 | Carissanol | [15] | Miscellaneous | - | - | √ | - |
| 85 | 20.81 | 238 | -/239.2097 | 221 | Germacrenone | [14,15] | Miscellaneous | √ | - | √ | - |
| 86 | 20.92 | 280 | 279.2659/- | 237, 222, 208, 194, 166, 152, 137, 111, 97, 83, 69, 57, 43 | Linoleic acid | [32] | Fatty acid | - | √ | - | - |
| 87 | 21.35 | 318 | 316.9698/- | 299, 289, 273, 245 | Dimethyl (epi)catechin | [27] | Flavan-3-ol | - | √ | - | - |
| 88 | 21.55 | 594 | 593.4359/- | 285 | Kaempferol-O-deoxyhexosyl-hexoside | [37] | Flavonoid | √ | - | √ | - |
| 89 | 22.09 | 594 | 593.4612/- | 285 | Kaempferol-3-O-rutinoside | [37] | Flavonoid | √ | - | √ | - |
| 90 | 22.29 | 328 | 327.4435/- | 309, 239, 229, 211, 171, 163 | Oxo-dihydroxy-octadecenoic acid | [29] | Fatty acid | - | √ | - | - |
| 91 | 22.36 | 328 | 327.4144/- | 309, 239, 229, 211, 171, 163 | Oxo-dihydroxy-octadecenoic acid isomer | [29] | Fatty acid | - | √ | - | - |
| 92 | 22.51 | 256 | 255.2160/- | 211, 183, 155, 127, 99 | Palmitic acid | [32] | Fatty acid | - | √ | - | - |
| 93 | 22.78 | 440 | -/441.3986 | 323,179, 161, 133 | Caffeoyl cyclohexanediol hexoside | [38] | Phenolic acid | - | √ | - | - |
| 94 | 22.90 | 422 | -/423.4181 | 307, 163, 145, 119 | p-Coumaroyl cyclohexanediol hexoside | [38] | Phenolic acids | - | √ | - | - |
| 95 | 23.12 | 328 | 317.0337/- | 179, 151, 137 | Myricetin | [39] | Flavonoid | √ | √ | - | - |
| 96 | 25.30 | 412 | -/413.3087 | 395, 256, 214 | Stigmasterol | [33,34] | Sterol | √ | √ | √ | - |
| 97 | 25.85 | 318 | 316.9427/- | 299 | Methyldihydroquercetin (Cedeodarin) | [39] | Flavonoid | √ | √ | - | - |
| 98 | 26.53 | 612 | -/613.6197 | 595, 521, 491, 449, 327, 287 | Rhamnosyl-hexosyl-methyl-quercetin | [38] | Flavonoid | - | - | √ | - |
| 99 | 27.02 | 464 | -/465.4451 | 301, 300, 257, 255, 229, 179. 151 | Hyperoside | [38] | Flavonoid | - | - | √ | √ |
| 100 | 27.21 | 464 | -/465.4246 | 301, 300, 257, 255, 229, 179. 151 | Isoquercetin | [38] | Flavonoid | - | - | √ | √ |
| 101 | 27.31 | 622 | 621.6783/- | 501 | 2(R)-26-([(2E)-3-(4-hydroxy-3-methoxyphenyl)-1-oxo-2- propen-1-yloxy)-2,3-dihydroxypropyl ester | [40] | Miscellaneous | √ | - | √ | √ |
| 102 | 31.25 | 430 | 429. 3132/430.9172 | 205, 191, 177, 149, 121 | α-Tocopherol | [41] | Miscellaneous | √ | √ | √ | √ |
| No. | Compound Name | HDAC6 | PDK3 | ||
|---|---|---|---|---|---|
| Score (kcal/mol) | RMSD Refine (Å) | Score (kcal/mol) | RMSD Refine (Å) | ||
| 6 | 3-O-Caffeolyquinic acid | −10.7626 | 1.40 | −13.1642 | 1.59 |
| 7 | 4-O-Caffeolyquinic acid | −16.1666 | 1.09 | −17.3380 | 1.26 |
| 8 | Coumaroyl-5-β-glucose | −9.1090 | 1.22 | −14.4638 | 0.97 |
| 9 | 5-O-Caffeolyquinic acid | −11.5285 | 1.77 | −14.4531 | 1.44 |
| 10 | Procyanidin Bl | −8.8345 | 2.19 | −15.8844 | 1.85 |
| 10a | Procyanidin B2 | −9.5399 | 1.93 | −17.5420 | 1.73 |
| 10b | Procyanidin B3 | −10.7608 | 1.26 | −17.4908 | 1.81 |
| 10c | Procyanidin B4 | −13.3266 | 2.04 | −16.6954 | 1.55 |
| 10d | Procyanidin B5 | −9.8460 | 2.31 | −23.9701 | 1.45 |
| 10e | Procyanidin B6 | −12.0986 | 1.64 | −19.2123 | 1.27 |
| 10f | Procyanidin B8 | −9.9105 | 1.38 | −18.6158 | 2.19 |
| 11 | Procyanidin C2 | −10.8164 | 2.61 | −24.2314 | 2.23 |
| 13 | Caffeic acid 3-glucoside | −17.0231 | 1.23 | −17.1953 | 1.15 |
| 14 | Catechin 3-O-β-D-glucopyranoside | −19.2377 | 2.56 | −15.7921 | 1.40 |
| 14a | Catechin 5-O-β-D-glucopyranoside | −15.7346 | 1.57 | −15.8820 | 1.62 |
| 14b | Catechin 7-O-β-D-glucopyranoside | −20.8137 | 1.38 | −21.3350 | 1.86 |
| 15 | 4-O-p-Coumaroylshikimic acid | −14.0624 | 0.89 | −14.3744 | 1.29 |
| 16 | Epicatechin 3-O-β-D-glucopyranoside | −23.6583 | 1.70 | −19.5019 | 1.39 |
| 16a | Epicatechin 6-C-glucoside | −15.2686 | 1.80 | −14.3566 | 1.26 |
| 16b | Epicatechin 8-C-glucoside | −7.8402 | 2.12 | −17.8381 | 1.60 |
| 16c | Epicatechin-3′-O-glucoside | −17.6491 | 1.42 | −14.1809 | 1.87 |
| 17 | Catechin | −10.1261 | 0.93 | −15.4482 | 0.83 |
| 17a | Epicatechin | −7.3314 | 1.64 | −14.7507 | 0.97 |
| 26 | Myricetin-3-O-xyloside | −9.5913 | 1.23 | −22.7275 | 1.36 |
| 27 | Quercetin | −12.8749 | 0.86 | −14.3401 | 1.29 |
| 29 | 1,4-Dicaffeoylquinic acid | −12.3875 | 1.16 | −18.8016 | 1.35 |
| 33 | Epigallocatechin-(4-β-6)-(+)-catechin | −11.4416 | 2.09 | −16.0503 | 1.89 |
| 33a | Epigallocatechin-(4-β-8)-catechin | −9.4011 | 1.35 | −19.7638 | 1.97 |
| 33b | Epicatechin (4 β.8) epigallocatechin | −9.1194 | 1.82 | −19.5053 | 1.61 |
| 33c | Catechin-(4α-8)-(-)-epigallocatechin | −12.9583 | 1.26 | −18.4410 | 1.08 |
| 35 | Syringic acid | −6.8037 | 2.86 | −10.8198 | 0.88 |
| 37 | Feruloyl-O-sinapoylquinic acid | −12.1606 | 1.41 | −17.0816 | 1.20 |
| 50 | 4″-Methyl-3-O-epigallocatechin gallate | −18.0349 | 0.97 | −17.8967 | 1.88 |
| 54 | Dimethoxycinnamoyl-O-caffeoylquinic acid | −8.1889 | 1.64 | −12.5443 | 1.88 |
| 74 | 3-Caffeoylshikimic acid | −14.4907 | 1.57 | −15.3836 | 1.40 |
| 75 | Dimethoxycinnamoylquinic acid | −14.6326 | 1.68 | −11.3988 | 1.69 |
| 77 | 5-p-Coumaroylquinic acid | −11.3266 | 1.65 | −16.5329 | 1.24 |
| 99 | Hyperoside | −9.9397 | 1.43 | −20.6101 | 1.49 |
| 100 | Isoquercetin | −10.0772 | 1.48 | −14.7778 | 1.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orabi, M.A.A.; Alqahtani, O.S.; Alyami, B.A.; Al Awadh, A.A.; Abdel-Sattar, E.-S.; Matsunami, K.; Hamdan, D.I.; Abouelela, M.E. Human Lung Cancer (A549) Cell Line Cytotoxicity and Anti-Leishmania major Activity of Carissa macrocarpa Leaves: A Study Supported by UPLC-ESI-MS/MS Metabolites Profiling and Molecular Docking. Pharmaceuticals 2022, 15, 1561. https://doi.org/10.3390/ph15121561
Orabi MAA, Alqahtani OS, Alyami BA, Al Awadh AA, Abdel-Sattar E-S, Matsunami K, Hamdan DI, Abouelela ME. Human Lung Cancer (A549) Cell Line Cytotoxicity and Anti-Leishmania major Activity of Carissa macrocarpa Leaves: A Study Supported by UPLC-ESI-MS/MS Metabolites Profiling and Molecular Docking. Pharmaceuticals. 2022; 15(12):1561. https://doi.org/10.3390/ph15121561
Chicago/Turabian StyleOrabi, Mohamed A. A., Omaish Salman Alqahtani, Bandar A. Alyami, Ahmed Abdullah Al Awadh, El-Shaymaa Abdel-Sattar, Katsuyoshi Matsunami, Dalia I. Hamdan, and Mohamed E. Abouelela. 2022. "Human Lung Cancer (A549) Cell Line Cytotoxicity and Anti-Leishmania major Activity of Carissa macrocarpa Leaves: A Study Supported by UPLC-ESI-MS/MS Metabolites Profiling and Molecular Docking" Pharmaceuticals 15, no. 12: 1561. https://doi.org/10.3390/ph15121561
APA StyleOrabi, M. A. A., Alqahtani, O. S., Alyami, B. A., Al Awadh, A. A., Abdel-Sattar, E.-S., Matsunami, K., Hamdan, D. I., & Abouelela, M. E. (2022). Human Lung Cancer (A549) Cell Line Cytotoxicity and Anti-Leishmania major Activity of Carissa macrocarpa Leaves: A Study Supported by UPLC-ESI-MS/MS Metabolites Profiling and Molecular Docking. Pharmaceuticals, 15(12), 1561. https://doi.org/10.3390/ph15121561